Abstracts
PURPOSE: Reactive astrocytes are implicated in several mechanisms after central or peripheral nervous system lesion, including neuroprotection, neuronal sprouting, neurotransmission and neuropathic pain. Schwann cells (SC), a peripheral glia, also react after nerve lesion favoring wound/repair, fiber outgrowth and neuronal regeneration. We investigated herein whether cell therapy for repair of lesioned sciatic nerve may change the pattern of astroglial activation in the spinal cord ventral or dorsal horn of the rat. METHODS: Injections of a cultured SC suspension or a lesioned spinal cord homogenized extract were made in a reservoir promoted by a contiguous double crush of the rat sciatic nerve. Local injection of phosphate buffered saline (PBS) served as control. One week later, rats were euthanized and spinal cord astrocytes were labeled by immunohistochemistry and quantified by means of quantitative image analysis. RESULTS: In the ipsilateral ventral horn, slight astroglial activations were seen after PBS or SC injections, however, a substantial activation was achieved after cord extract injection in the sciatic nerve reservoir. Moreover, SC suspension and cord extract injections were able to promote astroglial reaction in the spinal cord dorsal horn bilaterally. Conclusion: Spinal cord astrocytes react according to repair processes of axotomized nerve, which may influence the functional outcome. The event should be considered during the neurosurgery strategies.
Sciatic nerve lesion; Astrocyte; Neuronal protection; Neuroplasticity; Neuroregeneration; Pain; Review
OBJETIVO: Astrócitos reativos participam de vários mecanismos após lesões do sistema nervoso central e periférico, os quais incluem neuroproteção, brotamento neuronal, neurotransmissão e dor neuropática. As células de Schwann (CS), um tipo de glia periférica, também reagem com a lesão do nervo, podendo interferir com o reparo e cicatrização, crescimento de fibras e regeneração neuronais. Investigamos aqui a possibilidade da terapia celular para o reparo do nervo ciático poder alterar o padrão da ativação astrocitária nos cornos anterior e posterior da medula espinal do rato. MÉTODOS: Suspensão de CS cultivadas ou extrato homogeneizado de medula espinal lesada de rato foram inoculados num reservatório feito a partir de dois esmagamentos aplicados no nervo ciático do rato distantes 0,5mm entre si. Injeção local de salina tamponada serviu como controle. Os ratos foram mortos uma semana após e os astrócitos da medula espinal marcados por método imunohistoquímico e quantificados por análise de imagem. RESULTADOS: No corno anterior da medula, ipsilateral à lesão, ativação astrocitária leve foi vista após as injeções de tampão ou CS, entretanto, ativação celular intensa foi observada nesta região com a inoculação neural do extrato homogeneizado de tecido medular lesado. Adicionalmente, as inoculações de CS e de extrato homogeneizado de tecido medular promoveram forte reação astrocitária no corno dorsal da medula espinal, bilateralmente. CONCLUSÕES: Os astrócitos da medula espinal reagem em função do processo de reparo do nervo lesado, o que pode influenciar o resultado funcional esperado, algo que deve ser considerado durante o planejamento da estratégia neurocirúrgica.
Lesão do nervo ciático; Astrócito; Proteção neuronal; Neuroplasticidade; Neurorregeneração; Dor; Revisão
ORIGINAL ARTICLE
NEUROREGENERATION
Differential astroglial responses in the spinal cord of rats submitted to a sciatic nerve double crush treated with local injection of cultured Schwann cell suspension or lesioned spinal cord extract. Implications on cell therapy for nerve repair1 1 . Research performed at Neuroregeneration Center (LIM 45), Department of Neurology, School of Medicine, University of São Paulo (USP), Brazil.
Respostas astrocitárias na medula espinal do rato submetido ao esmagamento duplo do nervo ciático e tratado com injeção local de suspensão de células de Schwann cultivadas ou de extrato de medula espinal lesada. Implicações na terapia celular para o reparo do nervo
João Gabriel Martins DalloI; Bernardo Vergara ReichertI; José Benedito Ramos Valladão JúniorI; Camila SilvaII; Bianca Aparecida de LucaII; Beatriz de Freitas Azevedo LevyII; Gerson ChadiIII
IGraduate student, School of Medicine, USP, São Paulo, Brazil
IIFellow Master degree, Neuroscience and Behavior, Psychology Institute, USP, São Paulo, Brazil
IIIPhD, Full Professor, Experimental Neurology, Department of Neurology, School of Medicine, USP, São Paulo, Brazil
Correspondence Correspondence: Gerson Chadi Full Professor Department of Neurology University of São Paulo Av. Dr. Arnaldo, 455/2119 01246-903 São Paulo - Brazil Phone: (55 11)3061-7460 gerchadi@usp.br
ABSTRACT
PURPOSE: Reactive astrocytes are implicated in several mechanisms after central or peripheral nervous system lesion, including neuroprotection, neuronal sprouting, neurotransmission and neuropathic pain. Schwann cells (SC), a peripheral glia, also react after nerve lesion favoring wound/repair, fiber outgrowth and neuronal regeneration. We investigated herein whether cell therapy for repair of lesioned sciatic nerve may change the pattern of astroglial activation in the spinal cord ventral or dorsal horn of the rat.
METHODS: Injections of a cultured SC suspension or a lesioned spinal cord homogenized extract were made in a reservoir promoted by a contiguous double crush of the rat sciatic nerve. Local injection of phosphate buffered saline (PBS) served as control. One week later, rats were euthanized and spinal cord astrocytes were labeled by immunohistochemistry and quantified by means of quantitative image analysis.
RESULTS: In the ipsilateral ventral horn, slight astroglial activations were seen after PBS or SC injections, however, a substantial activation was achieved after cord extract injection in the sciatic nerve reservoir. Moreover, SC suspension and cord extract injections were able to promote astroglial reaction in the spinal cord dorsal horn bilaterally. Conclusion: Spinal cord astrocytes react according to repair processes of axotomized nerve, which may influence the functional outcome. The event should be considered during the neurosurgery strategies.
Key words: Sciatic nerve lesion. Astrocyte. Neuronal protection. Neuroplasticity. Neuroregeneration. Pain. Review.
RESUMO
OBJETIVO: Astrócitos reativos participam de vários mecanismos após lesões do sistema nervoso central e periférico, os quais incluem neuroproteção, brotamento neuronal, neurotransmissão e dor neuropática. As células de Schwann (CS), um tipo de glia periférica, também reagem com a lesão do nervo, podendo interferir com o reparo e cicatrização, crescimento de fibras e regeneração neuronais. Investigamos aqui a possibilidade da terapia celular para o reparo do nervo ciático poder alterar o padrão da ativação astrocitária nos cornos anterior e posterior da medula espinal do rato.
MÉTODOS: Suspensão de CS cultivadas ou extrato homogeneizado de medula espinal lesada de rato foram inoculados num reservatório feito a partir de dois esmagamentos aplicados no nervo ciático do rato distantes 0,5mm entre si. Injeção local de salina tamponada serviu como controle. Os ratos foram mortos uma semana após e os astrócitos da medula espinal marcados por método imunohistoquímico e quantificados por análise de imagem.
RESULTADOS: No corno anterior da medula, ipsilateral à lesão, ativação astrocitária leve foi vista após as injeções de tampão ou CS, entretanto, ativação celular intensa foi observada nesta região com a inoculação neural do extrato homogeneizado de tecido medular lesado. Adicionalmente, as inoculações de CS e de extrato homogeneizado de tecido medular promoveram forte reação astrocitária no corno dorsal da medula espinal, bilateralmente.
CONCLUSÕES: Os astrócitos da medula espinal reagem em função do processo de reparo do nervo lesado, o que pode influenciar o resultado funcional esperado, algo que deve ser considerado durante o planejamento da estratégia neurocirúrgica.
Descritores: Lesão do nervo ciático. Astrócito. Proteção neuronal. Neuroplasticidade. Neurorregeneração. Dor. Revisão.
Introduction
Axons of peripheral neurons possess the capacity of regeneration provided the repair of lesioned nerve stumps. The development of adequate microneurosurgery techniques for nerve repair has allowed axons to growth out of proximal stump, to cross the wound and to reenter distal stump, thus establishing functional contacts with appropriate targets 1,2. Cellular events that take place in the nerve region of regenerating fibers, as well as close to cell bodies (central nervous system in the case of motor neurons, and sensory ganglia in the case of sensory neurons) create the conditions to maintain neuronal trophism which is necessary to fiber outgrowth. These events however could be involved in the failure of regeneration and in the death of committed neurons 3,2. Glial cells are the pivotal elements involved in inflammation, wound, repair and neuroregeneration both in the central and peripheral nervous systems. Activated Schwann cells (SC) and satellite cells in the nerve, and also the activation of astrocytes and microglia in the spinal cord and brain regions express the molecules involved in the success or failure of axonal regeneration. Among many other substances, neurotrophic factors possess the ability to modulate regeneration 4,5. Cooperative responses of central and peripheral glial have been considered to be necessary to the process of regeneration of peripheral fibers, emphasizing the necessity to study the details of the central glial mechanisms after nerve injury. Axotomy of nerve fibers in the periphery triggers retrograde signals leading astroglial reaction close to perikarya of motor neurons or to axonal terminals of sensory neurons. For instance, when facial nerve is submitted either to axonal crush or transection, reactive astrocytes are seen in the facial nucleus in the pons 6-8. Moreover, abundant activated astrocytes are encountered in the anterior and posterior horns of the spinal cord after axotomy of sciatic nerve, which contains both sensory and motor fibers respectively 5. Reports have been mentioned that the adult sensory and motor neurons are partially resistant to axotomy because postnatal neurons diminish their trophic dependency from the targets thus becoming more dependent from the trophic support supplied by the cell body (autocrine regulation) and also by the neighbor non neuronal cells, particularly the adjacent glial cells, in a paracrine trophic fashion 2,5,9. Astrocytes are a class of glial cells which react to neuronal injury, becoming activated. Activated astrocytes are able to increase in number by proliferating cell precursors and to enlarge in shape by extending cell processes 10. The hypertrophic features of activated astrocytes facilitate environmental recognition, and subsequent deliver of molecules of neurotrophic properties 9 and also immunoregulatory modulators 11. In fact one of the most important roles of activated astrocytes is their ability to upregulate the synthesis of neurotrophic factor after neuronal injury triggering neuroprotection, wound/repair, neuroregeneration and neuronal plasticity 12. Glial cell proliferation in the dorsal horn induced either by diseases or physical trauma of peripheral nerves has recently become into attention because of the possibility of the glial involvement in naturopathic pain. Both inflammation1,3-15 and nerve injury 16 are able to activate glial cells in the dorsal horn of the spinal cord, which have been related to development and maintenance of persistent pain states15,17. Like the astroglia, which respond in the central nervous system after central or peripheral axotomy, Schwann cells, a type of glia encountered in the peripheral nervous system, which form myelin sheet of nerve axons, are also capable to become reactive after axotomy of peripheral nerves 8,18. Activated SC are also able to release neurotrophic factors contributing to protect and to stimulate neuronal regeneration 8. Schwann cell therapy has been proposed to fulfil long gaps between proximal and distal stumps of a lesioned nerve accompanied by nerve tissue lost. Interposed cultured SC on peripheral fiber regeneration and the functional outcome have not been analyzed in details yet. Moreover, in spite of the fact that those glial cells, the astrocyte in the central nervous system, and the SC in the peripheral nervous system, may favor protection and regeneration of axotomized peripheral neurons, the effects of a combined therapy employing both cells has not been investigated. The present work analyzed the effects of a cultured Schwann cell inoculation in a reservoir produced by a double contiguous crush in the rat sciatic nerve on the state of astroglial reaction in the correspondent level of the spinal cord. Considering the possible beneficial effects of local presence of SC, we have compared the results with the effects of a local application of a 7-day lesioned spinal cord extract which contains a great amount of neurodegenerative and also regenerative factors. Astroglial reaction was evaluated by means of a very specific immunohistochemistry labeling combined to quantitative image analysis.
Methods
Adult, male, pathogen-free Wistar rats from University of São Paulo Medical School, São Paulo, Brazil, [body weight (b.w.) 220-250 g] were used in the present study. Rats were kept under controlled temperature and humidity conditions with a standardized light and dark cycle (light on at 7 a.m. and off at 7 p.m.) and with free access to food pellets and tap water. The study was conducted under protocols approved by the Animal Care and Use Ethic Committee at the University of São Paulo and in accordance with the Guide for the Care and Use of Laboratory Animals adopted by the National Institutes of Health.
Schwann cell culture
Pure cultures of SC were obtained from sciatic nerves explants of adult Wistar rats as described previously 19. Briefly, sciatic nerves were dissected under sterile conditions, placed in 60 mm dishes containing Leibovitz-15 medium (L-15; Gibco, Grand Island, NY), and divested of their epineurial sheaths. Nerves were chopped into 1 mm2 pieces and transferred to 35mm dishes containing DMEM (Gibco) supplemented with 10% fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT), 50 mg/ml gentamycin (DMEM-10; Gibco, NY). Nerve pieces were transferred to another 35 mm dish every week for 5 weeks and were fed twice a week. After five transfers, explants were devoid of fibroblasts and contained only SC. Explants were replated onto 35 mm dishes with 1.25 U/ml dispase (Boehringer Mannheim, Mannheim, Germany), 0.05% collagenase (Worthington, Freehold, NJ), and 15% FBS in DMEM and were incubated at 37°C in 6% CO2 overnight. After incubation, explants were washed with DMEM-10 and dissociated, and the SC were seeded onto poly-L-lysine-coated 100 mm dishes at a density of 2-3x106 cells/dish. After 24 hours, the culture medium was changed to DMEM-10 supplemented with 2 mM forskolin (Sigma, St.Louis, MO) and 20 mg/ml pituitary extract (Gibco). Cells, fed twice a week with that medium, were allowed to expand until confluence. The purified SC were suspended in culture medium to yield a final density of 106 cells/ml.
Lesioned spinal cord extract
A separate set of rats was anaesthetized with sodium pentobarbital (45 mg/kg; Cristália, São Paulo, SP, Brazil) and placed in a microneurosurgical frame. Spinal cord contusion was performed as previously described 20. Briefly, the spinous process and the vertebral lamina were removed to expose a circular region of dura at the 10th thoracic spinal cord level. By means of the NYU Impactor, a contuse lesion of moderate severity was induced by dropping a device rod (10 g) from a distance of 25 mm onto the intact exposed dura. Animals received sodium cephalotin antibiotic (40 mg i.m. daily, Sigma Pharma, Hortolândia, Brazil) for 10 days. Fourteen days after surgery, animals were decapitated and their spinal cord removed quickly. Injury region was dissected out in small fragments, triturated manually and homogenized by means of centrifugation.
Microneurosurgery for crush lesion of the sciatic nerve and local inoculations
Rats (n=15) were anaesthetized with sodium pentobarbital 3% (minimum dose of 38mg/kg or maximum dose of 48 mg/kg, ip; Cristália). Prior to surgery, Gentamycin (100 mg/kg body weight, i.m.; Gibco) was administered, and Duratears lubricant eye ointment (Alcon Laboratories, Fort Worth, TX) was applied to the eyes to prevent drying. During surgery, animals were placed on a waterheated platform (37 ± 0.5 °C) to maintain body temperature. Using aseptic technique, the right sciatic nerve was exposed 1.0 cm distal to the sciatic notch, and then double crushed in two points separated by 0.5mm distance with small forceps for 30s. The space (0.5mm) between these contiguous crush lesions formed a short lasting reservoir (Figure 1) where inoculations were performed, as described bellow. Contralateral unoperated side served as control. The rats were randomly divided into three groups: PBS group (n = 5) received 5μl of phosphate buffered saline (PBS; pH 7.2, room temperature), cell suspension group (n = 5) received 5μl of 106 Schwann cells/ml suspended in culture medium, and cord extract group (n = 5) received 5μl of the spinal cord homogenate.
Tissue preparation and staining
Animals were deeply anaesthetized with sodium pentobarbital and sacrificed by a transcardiac perfusion with 100 ml isotonic saline at room temperature followed by 500 ml of fixation fluid (4ºC) over a period of 6 minutes 4,21,22. The fixative consisted of 4% paraformaldehyde (w/v, Merck, Germany) in 0.1 M phosphate buffer, pH 6.9. The lumbar level of spinal cords were removed (2.5 cm long), kept in the fixative solution at 4ºC for 90 minutes, and then rinsed in 10% sucrose (Merck) dissolved in 0.1 M phosphate buffered saline (PBS), pH 7.4, for 48 hours. The cords were orientated in the rostro-caudal axis and frozen in dry ice-cooled (-40ºC) isopentane (Sigma, USA) and stored at 70ºC freezer until use. Thaw-mounted sections were sampled systematically during sectioning. Series in a rostro-caudal order including every 100th section were obtained. One series of sections was submitted to cresyl violet staining in order to demonstrate Nissl substance in the spinal cord neurons and glial cells. Other series of sections was submitted to immunohistochemistry of glial fibrillary acid protein (GFAP), as a marker for astrocytes. Immunoreactivity was detected by the avidin-biotin peroxidase technique 23-25. Sections were washed for 2x10 min in PBS, and incubated with 5% normal goat serum for 30 min at room temperature. One series of spinal cord sections were incubated for 48 h at 4ºC with a rabbit polyclonal antiserum against GFAP (Dakopatts) diluted (1:1200) in PBS containing 0.5% Triton X-100 (Sigma) and 1% bovine serum albumin (Sigma). The sections were washed again in PBS (2x10 min) and incubated with biotinylated goat anti-rabbit immunoglobulins diluted 1:250 (Vector, USA) for 2 hours. After rinsing in PBS, they were incubated with an avidin-biotin peroxidase complex (both diluted 1:100, Vectastain, Vector) for 90 min. Immunoreactivity was visualized using 3-3'-diaminobenzidine tetrahydrocloride (Sigma) as a chromogen and H2O2 (0.05%, v/v, Sigma) for 8 min. The sections were analyzed and photographed with an Olympus AX70 photomiscroscope (USA).Control experiments. As a control, sections were incubated with the solvent of the primary and secondary antibody solutions and the solvent of the avidin-biotin solution, and processed simultaneously with the experimental sections.
Semiquantitative microdensitometric image analysis
The semiquantitative microdensitometric/morphometric image analysis of the GFAP immunoreactivity was performed in the sampled sections of rats from three groups. Fields of measurements were sampled in the anterior and posterior horns (gray matter), bilaterally. The procedures using a Kontron-Zeiss KS400 image analyzer (Germany) have been described previously 4,21,26,28. Briefly, a television camera acquired images from the microscope (63x objective). After shading correction, a discrimination procedure was performed as follows. The mean gray values (MGV) and s.e.m. of white matter or gray matter were measured in the above described sampled fields and also in corresponding areas of the spinal cord devoid of specific labeling (background, bg). Gray values darker than bgMGV-3 s.e.m. were considered specific labeling. The specific (sp) MGV was then defined as the difference between the bgMGV value and the MGV of the discriminated profiles (results not show in this paper). The size of the sampled field was 3.36 x 10-2 μm2. The glass value was kept constant at 200 MGV. The procedure was repeated for each section to correct every specific labeling measurement for background. In the morphometric evaluation, the area of the GFAP immunoreactivity discriminated astroglial profiles, including cytoplasm and process was measured and expressed as area per unit area (area/µm2).
Statistical analysis
Statistical analysis was performed using the parametric ANOVA to compare each side among groups and the two-tailed t-test to compare ipsilateral and contralateral sides.
Results
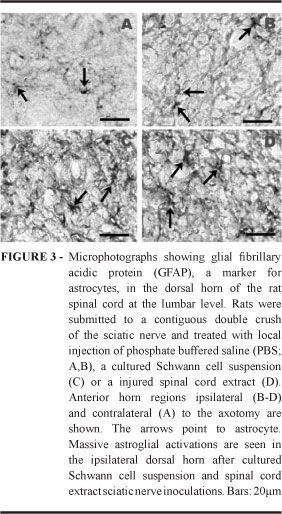
GFAP immunoreactive profiles resembling quiescent astrocytes with small size and thin processes were seen distributed throughout the white and gray matters of the studied levels of contralateral side of the sciatic nerve injury and treated with local injection of PBS (Figures 2 and 3). Moreover, an increased number of the GFAP immunoreactive profiles with large cytoplasm and thick processes was seen in the ventral and dorsal horns in the lumbar level of the spinal cords of the rats that received the crush injury of the sciatic nerve and treated with local injection of the SC suspension or spinal cord extracts (Figures 2 and 3).
The semiquantitative microdensitometric/morphometric image analysis of the GFAP positive astrocytes in the ipsilateral side of the ventral horn showed trends for increases of the area of the astroglial profiles after PBS (117%) and SC suspension (53%) treatments and a significant elevation after cord extract inoculation (90%) compared to respective contralateral side (Figure 4A). Furthermore, no changes among groups were detected in the ventral horn either in the ipsilateral side or in the contralateral side of sciatic nerve crush and local injections (Figure 4A). Moreover, GFAP positive astrocytes in the ipsilateral side of the dorsal horn showed significant elevations after PBS (279%), SC suspension (183%) and cord extract (136%) inoculations (Figure 4B) compared to respective contralateral side. ANOVA analysis of variance showed in the ipsilateral side of the dorsal horn a 53% increased of the area of the GFAP immunoreactive astrocytes of the SC suspension group compared to PBS treatment (Fig. 4B); the same analysis of the dorsal horn in the contralateral side showed that the area of GFAP positive astrocytes of the SC suspension group (105%) and the cord extract group (116%) were increased compared to PBS treatment (Figure 4B). The analysis of Nissl substance by means of cresyl violet staining showed an absence of changes in the number and in the shape of neuronal profiles in the ipsilateral ventral horn of rats submitted to SC suspension or cord extract treatments compared to PBS injection (Figure 5). Cresyl violet also stained glial profiles, however astroglial, microglial or olygodendroglial identity could not be distinguish by the method. Cresyl violet putative glial profiles were increased in the entire gray matter of the ipsilateral side of injections (Figure 5), and also in the contralateral side of SC suspension and cord extract treatments, remarkably in this late one, in which side differences could not be detected.

Discussion
It is well established that an adequate repair of the lesioned nerve prompts neuronal protection and survival, favoring axonal growth and reinnervation of appropriate targets, and also leading to degree of functional recovery. There are several mechanisms participating in the repair and regeneration of the peripheral neurons. Events that occur in the wound/repair zone depend mainly on the SC responses. SC signals in turn can be taken by the injured neurons triggering neuronal trophism and fiber outgrowth. SC signals are also responsible for fiber attraction, helping the growing fibers to reenter into the distal nerve stump and to extend towards denervated targets 8. It has been considered that the neurotrophic factors produced and released by SC are the mainly class of molecules involved in the local events related to wound, repair and regeneration. Those descriptions have stimulated researchers and surgeons to consider SC in the nerve repair, actually in the cases of a large gap promoted by the loss of nerve tissue in which is necessary to bridge the lesioned stumps. Interposed tube may be filled with cultured SC obtained from peaces of small sensory nerves of the same patient. These issues have been considered experimentally. Here we have not used the tubulization technique to repair sciatic nerve lesion and to deliver the treatments. Instead, we employed a double contiguous crush in order to promote a 5mm long reservoir in the rat sciatic nerve which is ensheathed by the undamaged perineurium. Within the reservoir, between the two applied crushes, a total axonal degeneration will take place, leaving intact the endoneural tubes which are important for immediate fiber growth. Local endogenous SC released by the disrupted myelin sheath, will initiate the repair process, which might be influenced by the local injections performed in this work. Astrocytes are the counterpart of SC in the central nervous system. Likewise, astrocytes in the brain and spinal cord react after injury by changing their shape and by producing neurotrophic factors and immunoregulatory molecules 27. Initial action is to promote wound and repair, by eliminating injured neurons and axons, and also restoring brain barriers 29. Subsequent events are related to further protection of remaining neurons, fiber growth and restoration of function 20. In the case of peripheral nerve lesion, local and retrograde signals can reach the central nervous system regions leading to astrocyte activation close to neuronal cell body and afferent projections of peripheral neurons. As there is no local wound related to tissue disruption, astroglial activation in the central nervous system after nerve lesion is mainly involved in paracrine mechanisms favoring neuronal survival and fiber outgrowth. It seems likely that the astroglial, centrally, and SC, peripherally, may cooperate in the process of nerve repair, however only recently this issue started to be investigated 8,27. Actually, there are many publications concerning the responses of peripheral neurons following a nerve lesion, however, a detailed analysis of the retrograde signals leading subsequent activation of glial cells close to motor neuron perikarya and to terminals of sensory neurons is still lacking. In fact, astrocytes can respond to signals from a variety of neuronal changes, including not only Wallerian degeneration, but also retrograde and transganglionic changes 30,31. Injury to peripheral nerve axons results in a series of structural and molecular alterations in the affected sensory and motor neurons. Downregulation of neuronal stability molecules like high molecular weight neurofilament proteins, and an upregulation of axonal sprouting/elongation molecules like GAP43, actin and tubulin are commonly seen 18. Furthermore, transganglionic changes extend to the central terminals of sensory neurons, which are accompanied by retraction and sprout of neighbor fibers. Moreover, retraction of dendritic tree of motor neurons is also seen after peripheral axotomy 32. The above described events are concomitant to local astroglial and microglial activation; however, glial responses initiate shortly after axotomy probably before the neuronal commitment. Initially, microglia become activated and release factors like interleukin- 1 and -6 33 which are able to promote astroglial cell proliferation and hypertrophy. Activated astrocytes in turn may maintain microglial activation by releasing colony stimulating factor-1 and other molecules 11. The interaction of axotomized neurons with activated non-neuronal cells may participate in a large number of mechanisms that are implicated on axonal growth/ neuronal plasticity responses and the functional implications started to be investigated. Hypertrophy and hyperplasia of astrocytes were found in the ipsilateral axotomized facial nucleus which lasted longer up to one year in the case of an impairment of axonal regeneration 34. Furthermore, a compression of facial nerve, by inserting a thin nylon thread in the facial canal, was enough to trigger astrocyte activation in the facial nucleus 35. Moreover, the intensity of the synaptic changes and the glial activation seen in axotomized facial nucleus were associated with the degree of nerve lesion, responses that could be reversed after nerve suture reanastomosis 6. Interestingly, mechanisms inducing gliosis following neuronal injury was also evaluated in vitro. Mechanically injured purified rat dorsal root ganglion neurons growing on monolayers of rat neonatal cortical astrocytes or sciatic nerve-derived SC led to astrocyte proliferation and morphological changes 7. Likewise, astroglial reaction has also been evaluated in the motor ventral horn after the lesion of sciatic nerve 1,9. We demonstrated here trends for glial activation after PBS and SC suspension injections in the ipsilateral ventral horn of the spinal cord gray matter compared to the contralateral side, responses that differed from those seen in the dorsal horn of those animals. These results are in line with previous descriptions on the central glial responses after peripheral nerve injury discussed above. It should be mentioned that glial activation does not occur only close to compromised neurons, but also it can be more widespread as recently described for small lesions in the central nervous system 12,36-38. It is possible that double crush lesion of the sciatic nerve plus local injections performed herein has also triggered some degree of astroglial reaction in the contralateral grey matter of the spinal cord, impairing demonstration of statistical differences antimerically. In fact, as we are going to discuss later, SC suspension and cord extract injections promoted also contralateral astroglial activation in the spinal cord dorsal horn compared to PBS injection, further emphasizing the involvement of contralateral astrocytes in the paracrine mechanisms after specific repair of unilateral axotomized sciatic nerve 39. These aspects may be further evaluated because PBS and SC suspension injections promoted astroglial activation in the dorsal horn compared to contralateral side, but only trends for activation have been seen in the ventral horn after those treatments. Differential glial responses in the dorsal and ventral horns after peripheral axotomy were already postulated, at least concerning the time course of microglial proliferation 40. The injection of the cord extract however triggered sufficient signals to promote astroglial activation in the ventral and dorsal horns independent of the contralateral responses compromising this glial cell population. Regarding the comparisons of the effects of treatments, CS suspension injection may have promoted a major response in the dorsal horn ipsilaterally while CS suspension and cord extract inoculations have led to additional changes contralaterally. Taken the results all together, it is likely that astroglial responses in ventral and dorsal horns are related to different mechanisms. In fact, as discussed above, differential responses are seen in axotomized sensory and motor peripheral neurons. It should be taken into attention that adult peripheral motor neurons are more resistant to insult than sensory neurons 18. Astrocytes and SC express several proteins with neurotrophic properties in vivo and invitro 41. Thus, SC suspension employed in this work was a source of neurotrophic factors. Moreover, a one week spinal cord lesion performed herein in order to obtain the cord extract was sufficient to trigger a massive astroglial activation with increased level of astroglial neurotrophic factor as well 12. However, cord extract may also possess a number of toxic substances which are released by degenerated neurons. It is possible that toxic substances of the cord extract employed herein generated retrograde signals which, after nerve inoculation, may have recruited spinal cord astrocytes close to motor cell body in the ventral horn to become activated in order to trigger local trophic (protective) mechanisms. Those events may have also happened close to sensory perikarya in the dorsal hoot ganglia, however they were not studied in this work but were evaluated previously by our group 8. Moreover, the neurotrophic source supplied by SC suspension and cord extract my have triggered retrograde signals in the sensory neurons able to induce astroglial activation in the dorsal horn, thus in the region of central projection of afferent from peripheral sensory axotomized neurons. If so, it becomes a very important issue to be analyzed because astrocytes have the ability to modulate neurotransmission at synapses. Impaired neurotransmission at spinal cord dorsal horn has been implicated in the mechanisms of chronic pain, which in fact is a common occurrence accompanying many cases of nerve lesion, repaired or not. It has to be emphasized that the neurotrophic factors at synapses play a pivotal role on local plasticity modulation, which is also related to chronic pain post injury. Researchers have been now investigated how activated astrocytes might modulate neurotrophic factor and other intra/inter cellular signaling leading to changes in neurotransmission at dorsal horn after a sensory fiber lesion, and also the possible implication for neuropathic pain. Responses of astrocytes and microglia to nerve injury differ temporally. While astroglial activation is characterized by a slight increase in the cell number/size that continues in a moderate and progressive manner, microglial reaction is an acute event 42. It was mentioned that activated microglia make an important contribution to the initiation of neuropathic pain while activated astrocytes might contribute to its maintenance. In fact, there have been additional indications that activated astrocytes are important for pain modulator. Activated astrocytes in the dorsal horn following sciatic nerve constriction were correlated with the degree of hyperalgesia 3 and tactile allodynia. Moreover, glial inhibitors exhibited anti-allodynic properties 13,14,43. Furthermore, impairments of glial signaling by drugs that target glial activation 44 inhibited the synthesis and/or action of pro-inflammatory cytokines 45,46 and reduced neuropathic pain experimentally. Other evidences are related to the increased levels of prostaglandin E2 (PGE2) and COX2 in the spinal cord after peripheral inflammation and neuropathic pain which might take place in local activated astrocytes. It should be taken into attention that cultured astrocytes stimulated by interleukin-1b can upregulate PGE2 and COX2, event that is mediated by ERK/MAP kinase and protein kinase C intracellular signaling in those glial cells 47,48. Nitric oxide production and kappa opioid receptor/endogenous dynorphin opioid regulation on activated astrocytes are also considered as additional mediators 49,50 (Figure 6).
Conclusion
The present paper evaluated the glial responses in the ventral and dorsal horns of the spinal cord of rats submitted to a double crush sciatic nerve injury and local injection of cultured SC suspension or cord extracts. Differential astroglial reaction in those regions may be related to specific mechanisms of reactive astroglial-promoted paracrine neurotrophic and neuroplastic responses to sensory and motor neurons (illustrated in Figure 6). The implications of central glial reactions on neuronal survival, regeneration and chronic pain must be further evaluated in relation to the microneurosurgical methods employed in nerve repair.
Received: April 12, 2007
Review: June 14, 2007
Accepted: July 18, 2007
Conflict of interest: none
Financial source: FAPESP (98/13122-5; 99/01319-1; 07/00491-3) and CNPq
- 1. Svensson M, Eriksson P, Persson JK, Molander C, Arvidsson J, Aldskogius H. The response of central glia to peripheral nerve injury. Brain Res Bull. 1993;30(3-4):499-506.
- 2. Rossi F, Gianola S, Corvetti L. Regulation of intrinsic neuronal properties for axon growth and regeneration. Prog Neurobiol. 2007;81(1):1-28.
- 3. Garrison CJ, Dougherty PM, Kajander KC, Carlton SM. Staining of glial fibrillary acidic protein (GFAP) in lumbar spinal cord increases following a sciatic nerve constriction injury. Brain Res. 1991;565(1):1-7.
- 4. Chadi G, Moller A, Rosen L, Janson AM, Agnati LA, Goldstein M, et al. Protective actions of human recombinant basic fibroblast growth factor on MPTP-lesioned nigrostriatal dopamine neurons after intraventricular infusion. Exp Brain Res. 1993;97(1):145-58.
- 5. Terenghi G. Peripheral nerve regeneration and neurotrophic factors. J Anat. 1999;194(Pt 1):1-14.
- 6. Guntinas-Lichius O, Neiss WF, Gunkel A, Stennert E. Differences in glial, synaptic and motoneuron responses in the facial nucleus of the rat brainstem following facial nerve resection and nerve suture reanastomosis. Eur Arch Otorhinolaryngol. 1994;251(7):410-7.
- 7. Guenard V, Frisch G, Wood PM. Effects of axonal injury on astrocyte proliferation and morphology in vitro: implications for astrogliosis. Exp Neurol. 1996;137(2):175-90.
- 8. Levy BFA, do Carmo Cunha J, Chadi G. Cellular analysis of the S100beta; and fibroblast growth factor-2 in the dorsal root ganglia and sciatic nerve of rodents. Focus on paracrine actions of activated satellite cells after axotoxy. Int J Neurosci. 2007;117:1481-503.
- 9. Aldskogius H, Liu L, Svensson M. Glial responses to synaptic damage and plasticity. J Neurosci Res. 1999;58(1):33-41.
- 10. Martins EF, Chadi G. Glial reaction in the hippocampus after global cardiogenic ischemia. Acta Cir Bras. 2001; 16 (1):05-14.
- 11. Raivich G, Jones LL, Werner A, Bluthmann H, Doetschmann T, Kreutzberg GW. Molecular signals for glial activation: pro- and anti-inflammatory cytokines in the injured brain. Acta Neurochir. 1999;73:21-30.
- 12. Leme RJ, Chadi G. Distant microglial and astroglial activation secondary to experimental spinal cord lesion. Arq Neuropsiquiatr. 2001;59(3-A):483-92.
- 13. Meller ST, Dykstra C, Grzybycki D, Murphy S, Gebhart GF. The possible role of glia in nociceptive processing and hyperalgesia in the spinal cord of the rat. Neuropharmacology .1994;33(11):1471-8.
- 14. Watkins LR, Martin D, Ulrich P, Tracey KJ, Maier SF. Evidence for the involvement of spinal cord glia in subcutaneous formalin induced hyperalgesia in the rat. Pain. 1997;71(3):225-35.
- 15. Sweitzer SM, Colburn RW, Rutkowski M, DeLeo JA. Acute peripheral inflammation induces moderate glial activation and spinal IL-1beta expression that correlates with pain behavior in the rat. Brain Res .1999;829(1-2):209-21.
- 16. DeLeo JA, Colburn RW, Rickman AJ. Cytokine and growth factor immunohistochemical spinal profiles in two animal models of mononeuropathy. Brain Res. 1997;759(1):50-7.
- 17. Coyle DE. Partial peripheral nerve injury leads to activation of astroglia and microglia which parallels the development of allodynic behavior. Glia. 1998;23(1):75-83.
- 18. Bisby MA, Tetzlaff W. Changes in cytoskeletal protein synthesis following axon injury and during axon regeneration. Mol Neurobiol. 1992;6(2-3):107-23.
- 19. Morrissey TK, Kleitman N, Bunge RP. Isolation and functional characterization of Schwann cells derived from adult peripheral nerve. J Neurosci. 1991;11(8):2433-42.
- 20. do Carmo Cunha J, de Freitas Azevedo Levy B, de Luca BA, de Andrade MS, Gomide VC, Chadi G. Responses of reactive astrocytes containing S100beta protein and fibroblast growth factor-2 in the border and in the adjacent preserved tissue after a contusion injury of the spinal cord in rats: implications for wound repair and neuroregeneration. Wound Repair Regen. 2007;15(1):134-46.
- 21. Chadi G, Rosen L, Cintra A, Tinner B, Zoli M, Pettersson RF, et al. Corticosterone increases FGF-2 (bFGF) immunoreactivity in the substantia nigra of the rat. Neuroreport. 1993;4(6):783-6.
- 22. Chadi G, Tinner B, Agnati LF, Fuxe K. Basic fibroblast growth factor (bFGF, FGF-2) immunoreactivity exists in the noradrenaline, adrenaline and 5-HT nerve cells of the rat brain. Neurosci Letter. 1993;160(2):171-6.
- 23. Gomide V, Chadi G. Glial bFGF and S100 immunoreactivities increase in ascending dopamine pathways following striatal 6-OHDA-induced partial lesion of the nigrostriatal system: a sterological analysis. Int J Neurosci. 2005;115(4):537-55.
- 24. Rodrigues RW, Gomide VC, Chadi G. Astroglial and microglial reaction after a partial nigrostriatal degeneration induced by the striatal injection of different doses of 6-hydroxydopamine. Int J Neurosci. 2001;109(1-2):91-126.
- 25. Rodrigues RW, Gomide VC, Chadi G. Astroglial and microglial activation in the wistar rat ventral tegmental area after a single striatal injection of 6-hydroxydopamine. Int J Neurosci. 2004;114(2):197-216.
- 26. Chadi G, Gomide VC. FGF-2 and S100beta immunoreactivities increase in reactive astrocytes, but not in microglia, in ascending dopamine pathways following a striatal 6-OHDA-induced partial lesion of the nigrostriatal system. Cell Biol Int. 2004;28(12):849-61.
- 27. Chadi G, Castelucci P, Gomide VC. Experimetal microneurosurgery of the central and peripheral nervous system in the study of the neuronal and glial trophism and plasticity. Acta Cir Bras. 1998;13(1):08-17.
- 28. Silva TP, Silveira GA, Fior-Chadi DR, Chadi G. Effects of ethanol consumption on vasopressin and neuropeptide Y immunoreactivity and mRNA expression in peripheral and central areas related to cardiovascular regulation. Alcohol. 2004;32(3):213-22.
- 29. Rodrigues RW, Gomide VC, Chadi G. Striatal injection of 6-hydroxydopamine induces retrograde degeneration and glial activation in the nigrostriatal pathway. Acta Cir Bras. 2003;18 (4):272-82.
- 30. Murray M, Wang SD, Goldberger ME, Levitt P. Modification of astrocytes in the spinal cord following dorsal root or peripheral nerve lesions. Exp Neurol. 1990;110(3):248-57.
- 31. Fior-Chadi DR, Varella TC, Maximino JR, Chadi G. Aortic coarctation hypertension induces fibroblast growth factor-2 immunoreactivity in the stimulated nucleus tractus solitarii. J Mol Histol. 2007;38(4):285-94.
- 32. Tetzlaff W, Graeber MB, Bisby MA, Kreutzberg GW. Increased glial fibrillary acidic protein synthesis in astrocytes during retrograde reaction of the rat facial nucleus. Glia. 1988;1(1):90-5.
- 33. Klein RD, Borchers AH, Sundareshan P, Bougelet C, Berkman MR, Nagle RB, et al. Interleukin-1beta secreted from monocytic cells induces the expression of matrilysin in the prostatic cell line LNCaP. J Biol Chem. 1997;272(22):14188-92.
- 34. Laskawi R, Wolff JR. Changes in glial fibrillary acidic protein immunoreactivity in the rat facial nucleus following various types of nerve lesions. Eur Arch Otorhinolaryngol. 1996;253(8):475-80.
- 35. Ruan RS, Leong SK, Yeoh KH. Glial reaction after facial nerve compression in the facial canal of the albino rat. Acta Otolaryngol. 1994;114(3):271-7.
- 36. Chadi G, Fuxe K. Analysis of trophic responses in lesioned brain: focus on basic fibroblast growth factor mechanisms. Braz J Med Biol Res. 1998;31(2):231-41.
- 37. Gomide VC, Silveira GA, Chadi G. Transient and widespread astroglial activation in the brain after a striatal 6-OHDA-induced partial lesion of the nigrostriatal system. Int J Neurosci. 2005;115(1):99-117.
- 38. Gomide VC, Chadi G. The trophic factors S-100beta and basic fibroblast growth factor are increased in the forebrain reactive astrocytes of adult callosotomized rat. Brain Res. 1999;835(2):162-74.
- 39. Firkins SS, Bates CA, Stelzner DJ. Corticospinal tract plasticity and astroglial reactivity after cervical spinal injury in the postnatal rat. Exp Neurol. 1993;120(1):1-15.
- 40. Echeverry S, Shi XQ, Zhang J. Characterization of cell proliferation in rat spinal cord following peripheral nerve injury and the relationship with neuropathic pain. Pain. 2007.
- 41. Hammarberg H, Risling M, Hokfelt T, Cullheim S, Piehl F. Expression of insulin-like growth factors and corresponding binding proteins (IGFBP 1-6) in rat spinal cord and peripheral nerve after axonal injuries. J Comp Neurol. 1998;400(1):57-72.
- 42. Zhang J, De Koninck Y. Spatial and temporal relationship between monocyte chemoattractant protein-1 expression and spinal glial activation following peripheral nerve injury. J Neurochem. 2006;97(3):772-83.
- 43. Sweitzer S, Martin D, DeLeo JA. Intrathecal interleukin-1 receptor antagonist in combination with soluble tumor necrosis factor receptor exhibits an anti-allodynic action in a rat model of neuropathic pain. Neuroscience. 2001;103(2):529-39.
- 44. Raghavendra V, Tanga F, DeLeo JA. Inhibition of microglial activation attenuates the development but not existing hypersensitivity in a rat model of neuropathy. J Pharmacol Exp Ther. 2003;306(2):624-30.
- 45. Sweitzer SM, Schubert P, DeLeo JA. Propentofylline, a glial modulating agent, exhibits antiallodynic properties in a rat model of neuropathic pain. J Pharmacol Exp Ther. 2001;297(3):1210-7.
- 46. Sweitzer SM, Medicherla S, Almirez R, Dugar S, Chakravarty S, Shumilla JA, et al. Antinociceptive action of a p38alpha MAPK inhibitor, SD-282, in a diabetic neuropathy model. Pain. 2004;109(3):409-19.
- 47. Molina-Holgado E, Ortiz S, Molina-Holgado F, Guaza C. Induction of COX-2 and PGE(2) biosynthesis by IL-1beta is mediated by PKC and mitogen-activated protein kinases in murine astrocytes. Br J Pharmacol. 2000;131(1):152-9.
- 48. Ma W, Quirion R. Partial sciatic nerve ligation induces increase in the phosphorylation of extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK) in astrocytes in the lumbar spinal dorsal horn and the gracile nucleus. Pain. 2002;99(1-2):175-84.
- 49. Yoon YW, Sung B, Chung JM. Nitric oxide mediates behavioral signs of neuropathic pain in an experimental rat model. Neuroreport. 1998;9(3):367-72.
- 50. Xu M, Bruchas MR, Ippolito DL, Gendron L, Chavkin C. Sciatic nerve ligation-induced proliferation of spinal cord astrocytes is mediated by kappa opioid activation of p38 mitogen-activated protein kinase. J Neurosci. 2007;27(10):2570-81.
Publication Dates
-
Publication in this collection
22 Jan 2008 -
Date of issue
Dec 2007
History
-
Reviewed
14 June 2007 -
Received
12 Apr 2007 -
Accepted
18 July 2007