Abstract
PURPOSE: To evaluate the gene expression of KGF, TNF-alpha and IL-1 beta in skin fibroblasts and keratinocytes cultured from burned patients. METHODS: Three patients with large burns and three patients with small burns, as well as two controls, were included. The cell culture was initiated by the enzymatic method. After extraction and purification of mRNA, qPCR was used to assess the gene expression of KGF, TNF-alpha and IL-1 beta. RESULTS: The expression of KGF was increased on average 220-fold in large burns and 33.33-fold in small burns in fibroblasts, and 11.2-fold in large burns and 3.45-fold in small burns in keratinocytes compared to healthy patients (p<0.05). Expression of TNF-alpha was not observed. IL-1 beta is down-regulated in fibroblasts of burned patients, and much more repressed in small burns (687-fold, p<0.05). In keratinocytes, the repression of IL-1 beta expression occurs in patients with small burns (28-fold), while patients with large burns express this gene intensively (15-fold). CONCLUSIONS: The study showed a quantitative pattern in the expression of KGF gene, which is more expressed according to the size of the burn. TNF-alpha was not expressed. A qualitative pattern in the expression of IL-1 beta gene was demonstrated.
Burns; Gene Expression; Fibroblasts; Keratinocytes; Tumor Necrosis Factor-alpha; Fibroblast Growth Factor 7; Interleukin-1
1 - ORIGINAL ARTICLE
MODELS, BIOLOGICAL
Keratinocyte growth factor, tumor necrosis factor-alpha and interleukin-1 beta gene expression in cultured fibroblasts and keratinocytes from burned patients1 1 Research performed at Translational Surgery Laboratory, Division of Plastic Surgery, Department of Surgery, Federal University of Sao Paulo, Paulista School of Medicine (UNIFESP-EPM), Brazil.
Alfredo GragnaniI; Bruno Rafael MüllerII; Ismael Dale Contrim Guerreiro da SilvaIII; Samuel Marcos Ribeiro de NoronhaIV; Lydia Masako FerreiraV
IPhD, Associate Professor, Division of Plastic Surgery, Department of Surgery, Federal University of Sao Paulo, Paulista School of Medicine (UNIFESP-EPM), Brazil. Conception, design, intellectual and scientific content of the study; analysis and interpretation of data; manuscript writing; critical revision
IIGraduate student, UNIFESP-EPM, Sao Paulo-SP, Brazil. Acquisition, analysis and interpretation of data; technical procedures; manuscript writing
IIIPhD, Associate Professor, Head, Laboratory for Molecular Gynecology, Department of Gynecology, UNIFESP-EPM, Sao Paulo-SP, Brazil. Technical procedures, conceived and designed the experiments
IVBSc, PhD, Post-doctoral student, Division of Plastic Surgery, Department of Surgery, UNIFESP-EPM, Sao Paulo-SP, Brazil. Acquisition, analysis and interpretation of data; technical procedures; manuscript writing
VPhD, Full Professor, Chairwoman, Division of Plastic Surgery, Department of Surgery, UNIFESP-EPM, Sao Paulo-SP, Brazil. Scientific and intellectual content of the study, critical revision
Correspondence Correspondence: Alfredo Gragnani Rua Napoleão de Barros, 715/4º andar 04024-002 São Paulo SP Brasil Tel.: (55 11)5576-4118 Fax: (55 11) 5571-6579 alfredogf@ig.com.br
ABSTRACT
PURPOSE: To evaluate the gene expression of KGF, TNF-alpha and IL-1 beta in skin fibroblasts and keratinocytes cultured from burned patients.
METHODS: Three patients with large burns and three patients with small burns, as well as two controls, were included. The cell culture was initiated by the enzymatic method. After extraction and purification of mRNA, qPCR was used to assess the gene expression of KGF, TNF-alpha and IL-1 beta.
RESULTS: The expression of KGF was increased on average 220-fold in large burns and 33.33-fold in small burns in fibroblasts, and 11.2-fold in large burns and 3.45-fold in small burns in keratinocytes compared to healthy patients (p<0.05). Expression of TNF-alpha was not observed. IL-1 beta is down-regulated in fibroblasts of burned patients, and much more repressed in small burns (687-fold, p<0.05). In keratinocytes, the repression of IL-1 beta expression occurs in patients with small burns (28-fold), while patients with large burns express this gene intensively (15-fold).
CONCLUSIONS: The study showed a quantitative pattern in the expression of KGF gene, which is more expressed according to the size of the burn. TNF-alpha was not expressed. A qualitative pattern in the expression of IL-1 beta gene was demonstrated.
Key words: Burns. Gene Expression. Fibroblasts. Keratinocytes. Tumor Necrosis Factor-alpha. Fibroblast Growth Factor 7. Interleukin-1.
Introduction
Burn injuries can influence many body functions and the consequences can be severe for these patients shortly after the accident. An intense inflammatory response is triggered in burns that affect a large total body surface area (TBSA)1. The beneficial effects of local inflammation include cleaning debris and protection against microbial agents, as well as cell growth and proliferation2.
However, the prolongation of the acute inflammatory process impairs wound healing, due to the fact that the high level of cytokines leads to degradation of collagen, apoptosis of keratinocytes, vascular commitment and production of oxygen-free radicals2.
Cytokines are the primary mediators of this inflammatory response to thermal injury. They promote communication between various types of cells via autocrine, paracrine and endocrine signaling. Thermal injury increases the serum level of cytokines1. Tumor necrosis factor-alpha (TNF-alpha) is involved in this process. The increased production of TNF-alpha is detectable in body fluids within the first twelve hours after trauma and is primarily released by macrophages in sites where it induces neutrophil recruitment and maturation3. Separately, it was shown that TNF-alpha inhibits the reepithelialization of wounds4. TNF-alpha is able to induce KGF production, which suggests that it operates indirectly in the reepithelialization process5.
The keratinocyte growth factor (KGF) or fibroblast growth factor-7 (FGF-7) is an important member of the family of FGFs involved in wound repair. Recent studies have suggested that the increase of KGF expression is a response to injuries in general. The repair of the injured epithelium depends on the mitogenic KGF power, and it has been noted that this growth factor can protect the epithelial cells of the skin against various insults in vivo and in vitro6. This factor is produced by dermal fibroblasts and acts by stimulating keratinocytes proliferation through receptors present on these cells4,7. The KGF gene expression by dermal fibroblasts is increased after cytokine signaling. And some of these cytokines are part of the Interleukin-1 family5.
The Interleukin-1 (IL-1) family cytokines are molecules that help to coordinate the early immune response against exogenous or endogenous danger. IL-1 alpha and IL-1 beta were the first members of this family to be described and now has 11 molecules of type IL-1. Due to its central role in the onset of inflammation, IL-1 has been an important therapeutic target. The IL-1 beta is a highly pyrogenic molecule and there are multiple levels of control of both its production and its effect8.
A different gene expression pattern is generated after thermal injury but no data are available in the literature. These genes are modulated during the inflammatory response in order to initiate subsequent events in wound repair9.
The aim of this study was to evaluate KGF, TNF-alpha and IL-1 beta gene expression in primary cultured dermal fibroblasts and keratinocytes from patients with large and small burns and compare it with that of the controls.
Methods
The project was approved by the Ethics Committee of Federal University of Sao Paulo (UNIFESP) (0689/11) and registered in Clinical Trials (NCT01302223).
This study had an experimental design in vitro, it was observational, analytical, controlled and it was conducted in a single center.
Samples were taken from three patients with large burns and three with small burns; two healthy patients in the control group were included in this study.
These samples were obtained from patients admitted to the Burns Treatment Unit, Plastic Surgery Division, Federal University of Sao Paulo, University Hospital (Table 1). The control group comprised two healthy, non-smoking female patients with no previous disease or medication use and undergoing aesthetic surgery. The first patient was 38 years old and underwent breast lift surgery; the second patient was 32 years old and underwent abdominoplasty.
Iinclusion, non-inclusion and exclusion criteria
Inclusion criteria for the study were patients of both genders, over 18 years old, who agreed to participate and signed a consent form, being hospitalized in Burns Unit and requiring surgery. A criterion was added to the group with large burns: having deep partial thickness or full thickness burns affecting between 25% and 50% of TBSA or which require partial skin graft in 10% TBSA. For the small burns group, the criterion added was that the TBSA be affected by 5% or less, for deep partial thickness or full thickness burns and the need of partial skin graft. To the control group was included the criterion of not having previous diseases, not smoking, and performing aesthetic surgery.
Inclusion criteria were also that the patient has not had previous skin diseases (psoriasis and similar), not had illnesses that might interfere directly in the inflammatory process (rheumatic diseases in general) and agreed to participate in the study.
Exclusion criteria were contamination of the culture flasks, low proliferation rate without achieving confluence of 80% of the cells in the flasks, insufficient quantity of extracted RNA that prevents the evaluation of patient data or loss of the extracted material.
Surgery
The debridement of burned tissue with deep partial thickness or full thickness burns occurred three to five days after injury. Normal skin around the lesion is removed because of the surgical procedure itself.
Keratinocyte culture
Keratinocytes derived from a skin fragment of burn patient were isolated and cultured according to the standard method, adapted in laboratory with changes as described below7.
In a 60 mm culture plate, the skin fragment was placed and sectioned into small pieces with size of about 0.5 cm3. These fragments were placed in a 50 ml sterile tube with 30 ml of dispase (Boeringer Mannheim, Indianapolis, IN; cat. no. 165859) and kept refrigerated at 4°C until the next morning, totaling 15 hours of action of dispase. After this period, each fragment was gently detached epidermis from the dermis with forceps; the epidermis was intended for isolation of keratinocytes, while the dermis underwent enzymatic dissociation with collagenase as described below.
Fibroblast culture
The culture was initiated by the enzymatic method using collagenase10. The dermis was placed in sterile Petri dishes. Collagenase solution sterile type-2 (Gibco, Grand Island, NY, USA - 17101-015) and diluted in PBS (3000 units/ml) and 3 ml of the solution for every gram of tissue was placed on the plate. This was made overnight at 37°C. After digestion of the tissue, the suspension was filtered through a 100micron filter and centrifuged at 400g for 10 min. The cells were centrifuged and placed in a flask with fibroblast culture medium11.
RNA isolation and purification
RNA was extracted from the cells using Trizol Reagent (Invitrogen, Foster City, CA) following the manufacturer's instructions. After complete dissociation of nucleoprotein complexes, phase separation was achieved with chloroform and centrifugation. The precipitated RNA from the aqueous phase was washed with 75% ethanol. The RNA was dried and dissolved in RNase-free water. Total RNA was then purified with QiagenRNeasyMiniKit (Qiagen, Valencia, CA) and submitted to DNAse treatment. The amount and quality of the extracted RNA were assessed by spectrophotometry using NanoDrop v3.3.0 (NanoDrop Technologies Inc., Rockland, DE).
qPCR
Commercially available TaqMan® Gene Expression Assay kits (Applied Biosystems, Foster City, CA) were used to assess TNF-alpha (Hs00174128_m1), FGF7 (KGF) (Hs00384281_m1), IL-1 beta (Hs01555410_m1) and B-actin (Hs99999903_m1) gene expressions on all samples. B-actin was used as housekeeping gene12. All qPCR reactions were performed on an ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, CA) using the fluorescent Taqman methodology (TaqMan One Step qPCR Master Mix Reagentes, Applied Biosytems). Total RNA (50 ng, 100 ng or 300 ng per well) was used for each qPCR reaction in a total volume of 25μL according to the manufacturer's protocol. Thermal cycling conditions were as follow: 30 s at 48ºC, 10 min at 95ºC, 40 cycles of 15 s, denaturation at 95ºC, and 60 s annealing at 60ºC. Quantification Cycle (Cq) values were used as endpoint defined as the PCR cycle number in which the fluorescence generated by the amplification crosses the threshold. Three replicate reactions per sample were run to ensure statistical significance.
Statistical analysis
Comparative Cq method (also known as the 2-DDqC method)13 was used for data analysis in order to calculate relative quantities (RQs) of gene expression among the samples. DataAssistTM, software developed for quick analysis of TaqMan® real-time PCR (Applied Biosystems, Foster City, CA), was used to confirm the data calculations performed in Microsoft® Office Excel. Whenever receptor quantification exceeded 36 amplification rounds (qC), quantities were considered to be undetectable or very low, and measurements were taken as invalid. Statistical significance was assessed using the Student's t-test. Statistical significance was defined at p<0.05.
Results
Extraction, purification and measurement of RNA
Samples of fibroblasts and keratinocytes were subjected to RNA extraction. Both the data from dosing solutions as well as their purity are presented in Table 2. The solutions presented RNA 260nm/280nm ratio values between 1.9 and 2.1, i.e., a purity range considered suitable for gene expression analysis.
KGF gene expression in fibroblasts
The KGF gene was significantly upregulated in fibroblasts from burned patients when compared to the control group. The small burns group presented a fold change value of 33.33, while in the large burns group this value was 220. There is no statistical difference between small and large burns groups (Figure 1).
KGF gene expression in keratinocytes
The KGF gene was upregulated in keratinocytes from burned patients when compared to the control group. The small burns group presented a fold change value of 3.45, while in the large burned group this value was 11.2. There is a significant difference between small and large burns groups (p=0.019) (Figure 2). KGF gene expression was modulated by the burn intensity in keratinocytes.
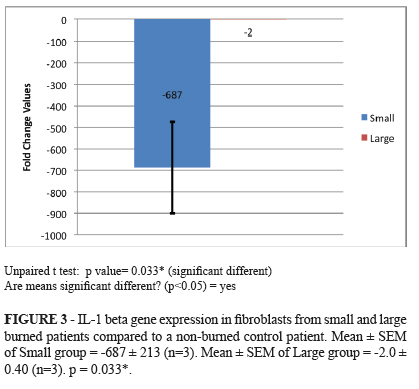
IL-1 beta gene expression in fibroblasts
The IL-1 beta gene was down-regulated in fibroblast of the burned patients when compared with the control group, and much more repressed in small burns. The fold change value of the small burned group was -687, while in the large group this value was -2.0 (p=0.033) (Figure 3).
Nevertheless, all patients continue to express the gene.
IL-1 beta gene expression in keratinocytes
Repression of IL-1 beta gene was high in small burns patients, while large burns patients expressed this gene intensively when compared to control (Figure 4). Burn intensity is controlling which intracellular pathways should be activated and deciding if IL-1B gene should be repressed or activated according to the changes in the microenvironment of the cells.
TNF-alpha gene expression in fibroblasts and keratinocytes
TNF-alfa gene expression was either not detected in fibroblasts and keratinocytes or occured at very low levels (Figure 5).
Discussion
Studies that evaluate gene expression of cytokines and growth factors by epidermal and dermal cells of the skin from burned patients do not exist in the literature. The few studies published on gene expression in burns were realized in blood cells and offered a vision of systemic inflammation in the body. Nedelec et al.14 demonstrated that a differential effect of burn injury occurs on fibroblasts from wounds and normal skin. Systemic mediators markedly alter the proliferation potential of dermal fibroblasts, but not fibroblasts from burn injuries. The wound environment substantially alters both the proliferation rate and the responsiveness of fibroblasts to cytokines. Thus, the data support the value of using burn wound fibroblasts in vitro experiments to investigate the change of wound healing by cytokine manipulation14.
The present study is the first to evaluate the gene expression of KGF, TNF-alpha and IL-1 beta, mediators with a central role in the inflammation process in human skin cells from burned patients.
KGF is an important growth factor involved in the reepithelialization of lesions. This factor is produced by dermal fibroblasts, and for this reason KGF gene expression is increased in these cells during the acute phase of burns in order to accelerate cleaning and repair of damaged tissues. Our results showed that this gene is upregulated, on average, 125-fold in skin fibroblasts and 7.5-fold in keratinocytes from burned patients than in control patients. This suggests that thermal injury is an event with very strong signal to stimulate the expression, and perhaps the production of KGF. This also indicated a quantitative pattern of expression. The greater the extension of burned area higher the KGF expression by skin cells of the patient.
Chedid et al.15 studied cytokines and growth factors that influenced the production of KGF by fibroblasts. An embryonic fibroblast culture (M426), a postectomy skin fragment of a newborn and an adult dermis fragment were used in this study. It demonstrated that IL-1 induced a marked increase in KGF RNA levels. This effect was dose dependent, blocked in the presence of anti-IL-1 and accompanied by an increase of active KGF protein. The results suggested that other growth factors such as PDGF-beta, IL-6 and TGF-alpha moderately stimulate KGF RNA. These data suggested that a variety of growth factors and cytokines can influence the production of KGF in vivo15.
The increase in KGF gene expression for injured tissue has already been demonstrated by Brauchle et al.16, who used Western blot analysis to show an increased KGF expression on surgical specimens of patients suffering from Crohn's disease and ulcerative colitis.
Li et al.17, through analysis by qPCR, demonstrated that KGF is expressed in healthy periodontal tissue, and also found that the expression levels of KGF increased significantly in the injured periodontal tissues.
The present study also found this KGF overexpression by fibroblasts and keratinocytes in an injured tissue. In this case, the skin of burned victims was examined, and it was observed that thermal injury significantly increased the gene expression of KGF by dermal fibroblasts and keratinocytes that are around the burn.
Analysis of IL-1 beta showed a qualitative pattern on gene expression in the present study. This cytokine expression was substantially suppressed in patients with small burns, both in dermal fibroblasts and in keratinocytes. In keratinocytes of large burns there is an increase in gene expression.
Hu et al.18 investigate the role of IL-1 in incisional wound biology and its effect on wound chemokine production in vivo and whether this mechanism could be active in human subjects. A well-characterized murine model of incisional wounding was used to assess the in vivo role of IL-1 in wound biology. The amount of seven different cytokines/chemokines produced within an experimentally induced skin incision on a mouse paw and the nociceptive response was analyzed in mice treated with an IL-1 inhibitor. They also investigated whether human IL-1 beta or IL-1 alpha stimulated the production of chemokines by primary human keratinocytes in vitro, and whether there was a correlation between IL-1 beta and chemokine levels in two experimental human wound paradigms. Administration of an IL-1 receptor antagonist to mice decreased the nociceptive response to an incisional wound, and reduced the production of multiple inflammatory mediators, including keratinocyte-derived chemokine (KC) and macrophage inhibitory protein (MIP)-1 alpha, within the wounds. IL-1 alpha and IL-1 beta stimulated IL-8 and GRO-alpha (human homologues of murine keratinocyte-derived chemokine) production by primary human keratinocytes in vitro. IL-1 beta levels were highly correlated with IL-8 in human surgical wounds, and at cutaneous sites of human ultraviolet B-induced sunburn injury. IL-1 plays a major role in regulating inflammatory mediator production in wounds through a novel mechanism; by stimulating the production of multiple cytokines and chemokines, it impacts clinically important aspects of wound biology. These data suggest that administration of an IL-1 receptor antagonist within the perioperative period could decrease postsurgical wound pain18.
With the same approach, in our results, the gene expression of IL-1 beta in human keratinocytes were more expressed in large burns, so if an IL-1 receptor antagonist would be used, we will can control the inflammatory process exaggerated by the stimulus to cytokine production and determine an improved clinical evolution.
This study demonstrated that TNF-alpha was not expressed by dermal fibroblasts and keratinocytes, both in burned patients as the control group, suggesting that during the acute inflammatory process other cells must express this gene and thus secrete the growth factor.
The effects of exogenous TNF-alpha depend on the concentration and duration of exposure, and this emphasizes the importance of balancing pro-inflammatory signals that control wound healing. At low concentrations, TNF-alpha can stimulate the healing process since it indirectly induces inflammation and releases growth factors4.
Cytokines produced during acute inflammation, such as IL-1 and TNF-alpha, stimulate fibroblasts to synthesize other pro-inflammatory cytokines, creating a loop that is responsible for the magnification and even for the perpetuation of the inflammatory process19.
Finnerty et al.20, after realizing that burned adults have higher morbidity and mortality than burned children, compared the temporal profile of cytokine concentrations in plasma of burned adults and children. The temporal profile of cytokines was different between burned children and adults, but no significant differences were detected between TNF-alpha concentrations between the two groups. Other pro-inflammatory and anti-inflammatory cytokines had large differences in concentrations in adults and children with extensive burns. Adults had a hyper-inflammatory pattern compared to children. Plasma concentrations of cytokines were analyzed using a multiplex assay kit array system that evaluated 22 cytokines20.
It was demonstrated that these mediators have a central role in inflammation, and they are extremely important to understand the response to thermal injury6,8,18. It has also been shown that there is an interaction between IL-1 beta, TNF-alpha and KGF during inflammation and wound repair21.
The originality of this article lies in the fact that is the first one to assess the gene expression of TNF-alpha, KGF and IL-1 beta in epidermal and dermal cultured cells of burned patients and compare them with those of healthy people.
Conclusions
The study showed a quantitative pattern in the expression of KGF gene, which is more expressed accord to the size of the burn. TNF-alpha was not expressed by dermal fibroblasts and keratinocytes, suggesting that during the acute inflammatory process other cells must express this gene. A qualitative pattern in the expression of IL-1 beta gene was demonstrated.
Acknowledgments
To Sao Paulo Research Foundation (FAPESP), National Council of Technological and Scientific Development (CNPq), Institutional Program of Scientific Initiation Scholarships (PIBIC).
Received: April 15, 2013
Review: June 17, 2013
Accepted: July 18, 2013
Conflict of interest: none
Financial sources: FAPESP (number 2011/12.945-4) and CNPq
- 1. Gauglitz GG, Song J, Herndon DN, Finnerty CC, Boehning D, Barral JM, Jeschke MG. Characterization of the inflammatory response during acute and post-acute phases after severe burn. Shock. 2008;30(5):503-7.
- 2. Shupp JW, Nasabzadeh TJ, Rosenthal DS, Jordan MH, Fidler P, Jeng JC. A review of the local pathophysiologic bases of burn wound progression. J Burn Care Res. 2010;31(6):849-73.
- 3. Rumalla VK, Borah GL. Cytokines, growth factors, and plastic surgery. Plast Reconstr Surg. 2001;108(3):719-33.
- 4. Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008;16:585-601.
- 5. Brauchle M, Angermeyer K, Hübner G, Werner S, Large induction of keratinocyte growth factor expression by serum growth factors and pro-inflammatory cytokines in cultured fibroblasts. Oncogene. 1994;9:3199-204.
- 6. Ferreira LM, Gragnani A, Furtado F, Hochman B. Control of the skin scarring response. An Acad Bras Cienc. 2009;81:623-9.
- 7. Gragnani A, Rossi MB, Albuquerque JC, Brito GS, Aloise AC, Ferreira LM. Keratinocyte growth factor protected cultured human keratinocytes exposed to oxidative stress. Acta Cir Bras. 2010;25:93-7.
- 8. Goldbach-Mansky R, Kastner DL. Autoinflammation: the prominent role of IL-1 in monogenic autoinflammatory diseases and implications for common illnesses. J Allergy Clin Immunol. 2009;124:1141-9.
- 9. Greco 3rd JA, Pollins AC, Boone BE, Levy SE, Nanney LB. A microarray analysis of temporal gene expression profiles in thermally injured human skin. Burns. 2010;36:192204.
- 10. Hirsch T, von Peter S, Dubin G, Mittler D, Jacobsen F, Lehnhardt M, Eriksson E, Steinau HU, Steinstraesser L. Adenoviral gene delivery to primary human cutaneous cells and burn wounds. Mol Med. 2006;12(9-10):199-207.
- 11. Campaner AB, Ferreira LM, Gragnani A, Cusick JL, Morgan JR. Up-regulation of TGF-β1 expression may be necessary but is not sufficient for excessive scarring. J Invest Dermatol. 2006;5:1168-76.
- 12. Greer S, Honeywell R, Geletu M, Arulanandam R, Raptis L. Housekeeping genes; expression levels may change with density of cultured cells. J Immunol Methods. 2010;355(1-2):76-9.
- 13. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3(6):1101-8.
- 14. Nedelec B, De Oliveira A, Saint-Cyr M, Garrel DR. Differential effect of burn injury on fibroblasts from wounds and normal skin. Plast Reconstr Surg. 2007;119(7):2101-9.
- 15. Chedid M, Rubin JS, Csaky KG, Aaronson SA. Regulation of keratinocyte growth factor gene expression by interleukin 1. J Biol Chem. 1994;269:10753-7.
- 16. Brauchle M, Madlener M, Wagner AD, Angermeyer K, Lauer U, Hofschneider PH, Gregor M, Werner S. Keratinocyte growth factor is highly overexpressed in inflammatory bowel disease. Am J Pathol. 1996;149(2):521-9.
- 17. Li M, Firth JD, Putnins EE. Keratinocyte growth factor-1 expression in healthy and diseased human periodontal tissues. J Periodont Res. 2005;40(2):118-28.
- 18. Hu Y, Liang D, Li X, Liu HH, Zhang X, Zheng M, Dill D, Shi X, Qiao Y, Yeomans D, Carvalho B, Angst MS, Clark JD, Peltz G. The role of interleukin-1 in wound biology. Part II: In vivo and human translational studies. Anesth Analg. 2010;111(6):1534-42.
- 19. Jordana M, Särnstrand B, Sime PJ, Ramis I. Immune-inflammatory functions of fibroblasts. Eur Resp J. 1994;7:2212-22.
- 20. Finnerty CC, Jeschke MG, Herndon DN, Gamelli R, Gibran N, Klein M, Silver G, Arnoldo B, Remick D, Tompkins RG; Investigators of the Inflammation and the Host Response Glue Grant. Temporal cytokine profiles in severely burned patients: a comparison of adults and children. Mol Med. 2008;14:553-60.
- 21. Maas-Szabowski N, Stark HJ, Fusenig NE. Keratinocyte growth regulation in defined organotypic cultures through IL-1-induced keratinocyte growth factor expression in resting fibroblasts. J Invest Dermatol. 2000;114(6):1075-84.
Publication Dates
-
Publication in this collection
26 July 2013 -
Date of issue
Aug 2013
History
-
Received
15 Apr 2013 -
Accepted
18 July 2013 -
Reviewed
17 June 2013