Abstract
Purpose:
To investigate the mechanisms by which PD98059 and LY294002 interfere with the abnormal deposition of extracellular matrix regulated by connective tissue growth factor (CTGF) of rat pulmonary artery smooth muscle cells (PASMCs).
Methods:
Rat PASMCs were cultured and separated into a control group. Real-time fluorescence quantitative PCR was performed to detect the expression of collagen III and fibronectin mRNA. Immunohistochemistry and western blot analyses were performed to detect the expression of collagen III protein.
Results:
The expression of collagen III and fibronectin mRNA was greater in PASMCs stimulated with CTGF for 48 h, than in the control group. After 72h of stimulation, the expression of collagen III protein in the PASMCs was greater than in the control. The equivalent gene and protein expression of the CPL group were much more significant.
Conclusions:
CTGF can stimulate the gene expression of collagen III and fibronectin in PASMCs, which may be one of the factors that promote pulmonary vascular remodeling (PVR) under the conditions of pulmonary arterial hypertension (PAH). PD98059 and LY294002 can inhibit the ERK1/2 and PI3K/PKB signaling pathways, respectively, thus interfering with the biological effects of CTGF. This may be a new way to reduce PAH-PVR.
Key words:
Connective Tissue Growth Factor; Pulmonary Artery; Muscle Cells; Extracellular Matrix; Rats
Introduction
The pathological feature of Pulmonary arterial hypertension (PAH) is pulmonary vascular remodeling (PVR) in which dysfunction of endothelial cells, neointimal formation, pulmonary arterial smooth muscle cells (PASMCs) proliferation, and the abnormal deposition of extracellular matrix (ECM) play important roles.Previous studies have found that the PASMCs can produce large amounts of ECM when stimulated by various unusual factors11 Schuurhof A, Bont L, Hodemaekers HM, de Klerk A, de Groot H, Hofland RW, van de Pol AC, Kimpen JL, Janssen R. Proteins involved in extracellular matrix dynamics are associated with respiratory syncytial virus disease severity. Eur Respir J. 2012 Jun;39(6):1475-81. doi: 10.1183/09031936.00012311.
https://doi.org/10.1183/09031936.0001231...
2 Kelvin EA, Edwards S, Jedrychowski W, Schleicher RL, Camann D, Tang D, Perera FP. Modulation of the effect of prenatal PAH exposure on PAH-DNA adducts in cord blood by plasma antioxidants. Cancer Epidemiol Biomarkers Prev. 2009 Aug;18(8):2262-8. doi: 10.1158/1055-9965.EPI-09-0316.
https://doi.org/10.1158/1055-9965.EPI-09...
3 Pi L, Shenoy AK, Liu J, Kim S, Nelson N, Xia H, Hauswirth WW, Petersen BE, Schultz GS, Scott EW. CCN2/CTGFregulates neovessel formation via targeting structurally conserved cystine knot motifs in multiple angiogenic regulators. FASEB J. 2012 Aug;26(8):3365-79. doi: 10.1096/fj.11-200154.
https://doi.org/10.1096/fj.11-200154...
-44 Sakai N, Chun J, Duffield JS, Wada T, Luster AD, Tager AM. LPA1-induced cytoskeleton reorganization drives fibrosis through CTGF- dependent fibroblast proliferation. FASEB J. 2013 May;27(5):1830-46. doi: 10.1096/fj.12-219378.
https://doi.org/10.1096/fj.12-219378...
. Connective tissue growth factor (CTGF) is an bioactive factor that was recently discovered to be closely associated with the abnormal deposition of ECM. Studies had shown that the gene and protein expression of CTGF is significantly higher in the PASMCs of a PAH animal model55 Liu B, Wang XM, Zhou TF, Hua YM, Liu HM, Wei L, Qiao LN, Wang XQ, Zhao SS, Shi K. Expression of connective tissue growth factor and its down-regulation by simvastatin administration in pulmonary hypertensive rats. Zhonghua Er Ke Za Zhi. 2008 May;46(5):359-65. PMID: 19099753.. This suggests that CTGF may be a factor that can promote abnormal ECM deposition by PASMCs. The effects of extracellular regulation kinase (ERK1/2) and inositol trisphosphate kinase (PI3K/PKB) signal pathways play important roles in the proliferation of a variety of cell types and the expression of several genes in various cells types from in vivo studies66 Sbroggiò M, Bertero A, Velasco S, Fusella F, De Blasio E, Bahou WF, Silengo L, Turco E, Brancaccio M, Tarone G. ERK1/2 activation in heart is controlled by melusin, focal adhesion kinase and the scaffold protein IQGAP1. J Cell Sci. 2011 Oct 15;124(Pt 20):3515-24. doi: 10.1242/jcs.091140.
https://doi.org/10.1242/jcs.091140...
7 van de Laar L, van den Bosch A, Boonstra A, Binda RS, Buitenhuis M, Janssen HL, Coffer PJ, Woltman AM. PI3K-PKB hyperactivation augments human plasmacytoid dendritic cell development and function. Blood. 2012 Dec 13;120(25):4982-91. doi: 10.1182/blood-2012-02-413229.
https://doi.org/10.1182/blood-2012-02-41...
-88 Foster JG, Blunt MD, Carter E, Ward SG. Inhibition of PI3K signaling spurs new therapeutic opportunities in inflammatory/autoimmune diseases and hematological malignancies. Pharmacol Rev. 2012 Oct;64(4):1027-54. doi: 10.1124/pr.110.004051.
https://doi.org/10.1124/pr.110.004051...
. In this study rat PASMCs were cultured with CTGF in vitro, and then interfere with PD98059 or/and LY294002 to observe the roles of the ERK1/2 and PI3K/PKB signaling pathways in CTGF-regulated ECM deposition, with the aim of investigating the pathogenetic mechanisms of PAH-PVR.
Methods
Culture and identification of PASMC
Healthy male Sprague Dawley rats (approximately 120g) were killed by using a neck-breaking method. This study was conducted in accordance with the declaration of Helsinki. This study was conducted with approval from the Ethics Committee of the Affiliated Hospital of Southwest Medical University. Written informed consent was obtained from all participants.
The pulmonary artery was then extracted, the fibrous tunica externa dissected and the runica intima stripped. The myogenous tissues of the vascular middle layer of the artery were then cut into 1 mm³ tissue blocks, and seeded into the culture flasks with the density as 1cm-2. After 2 h wall-adherent cultivation at 37°C and 5% CO2, M199 medium containing 10% fetal bovine serum (FBS) was added. A week later, cells had grown outwards from the tissue blocks; when the growth covered 70% of the area of the base of the vessel, the culture was passaged. The 3rd-generation cells were taken for slice culturing and an immunohistochemical method was performed to detect the expression of cellular α-SMA and to confirm the purity of the PASMCs.
The PASMCs were then divided into five groups and cultured in M199 medium containing different additional components, and for 48 h and 72 h. The control group was cultivated in M199 medium with 10% FBS. Indeed, in this study, the control group is negative control. The CTGF group with 10% FBS + 50 ng/ml CTGF (PeproTech, USA), the CP group with 10% FBS + 50 ng/ml CTGF + 20 μmol/L PD98059 (Sigma, USA), the CL group with 10% FBS + 50 ng/ml CTGF + 10 μmol/L LY294002 (Sigma, USA) and the CPL group with 10% FBS + 50 ng/ml CTGF + 20 μmol/L PD98059 + 10 μmol/L LY294002.
RT-PCR
For the RT-PCR, the cDNA sequences of rat collagen III, fibronectin and β-actin were downloaded from NCBI gene database, then the Primer Premier 5 software was used to design appropriate PCR primers. The primer sequences were in the Table 1.
After cultivation for 48 h, the cellular total RNA was isolated and extracted by using a Trizol method (Invitrogen, USA). Then 1 μg total RNA was used as the template for reverse transcription using a Revert Aid (tm) First Strand cDNA Synthesis Kit, where the reaction parameters were: 20°C for 10 min, 42°C for 60 min, 70°C for 10 min. One microliter of the cDNA reverse transcription product was then used to perform a quantitative PCR by the RT-qPCR method, where 45 cycles with the following parameters were performed: 94°C for 2 min, 94 °C for 30 s, 55°C for 30 s, 72°C for 40 s. During the calculation of the expression of target genes, the endogenous housekeeping gene β-actin was selected as an internal standard. The comparative threshold method was used to calculate the expression level (the relative expression of the target gene=2- ΔΔ Ct), then the relative initial amounts of each sample were calculated. The relative expression value was used to determine the difference in the expression of a target gene between the experimental group and the control group, as this method directly quantifies the target gene expression relative to that of the housekeeping gene.
Immunohistochemical staining
The cells in the logarithmic growth phase were seeded into 6-well plates for the slice climbing. After wall-adherent culture for 24 h, the culture media were changed according to the experimental grouping and incubated for a further 48 h or 72 h, at which point the collagen III immunohistochemical staining was carried out. The images of the stained cells were processed using the Photoshop 10.0 (Adobe Systems; Mountain View, CA, USA) image processing software, using five randomly located sites inside the stained cytoplasm for measuring the R-value99 Takei S, Hasegawa-Ishii S, Uekawa A, Chiba Y, Umegaki H, Hosokawa M, Woodward DF, Watanabe K, Shimada A. Immunohistochemical demonstra- tion of increased prostaglandin F2a levels in the rat hippocampus following kainic acid-induced seizures. Neuroscience. 2012 Aug;218:295-304. doi: 10.1016/j. neuroscience.2012.05.013.
https://doi.org/10.1016/j...
on one colony. There are 5 different colonies used for analysis. The smaller the mean R value, the deeper the staining, and the greater the protein expression.
Western blot
Plasma proteins were extracted from cell lysates, and 20 μg of these proteins were added into each lane of a 6% SDS-PAGE gel, the proteins were then electrically transferred onto a polyvinylidene difluoride (PVDF) membrane; after blocking at 37°C for 1h, the monoclonal primary antibody of collagen III (1:50; Santa Cruz, United States) was added prior to overnight incubation at 4°C. The membrane was then rinsed thoroughly with TBS, and horseradish peroxidase-coated goat anti-mouse secondary antibody (1:500; Beyotime, Shanghai, China) was added prior to incubation for 1 h at 37°C. A chemiluminescence method was used to detect the presence of protein-antibody complexes by using a gel imaging system (Fujifilm, Tokyo, Japan). The Quantity One (Bio-Rad, China) analysis software was then used to scan the gray values of the various bands to calculate the relative expression levels of the various proteins relative to the internal standard (Tubulin).
Statistical analysis
The SPSS 15.0 software was used for the statistical analysis. The measurement data were expressed as a mean and standard deviation. A single-factor analysis of variance was performed, and a pairwise comparison of intergroup means was performed using the LSD method, where p<0.05 was considered to indicate statistical significance.
Results
Culture and identification of PASMC
Under an inverted phase contrast microscope, the cells were observed to be fusiform, with the nuclei being rod-shaped or oval, and with two or more nucleoli. The cells grew in parallel and were arranged in bundles, exhibiting a typical "trough-like" growth pattern. The results of immunohistochemical staining showed that >95% of cells were α-SMA positive, suggesting that they were PASMCs.
Expression of collagen III and fibronectin mRNA
After cultivation for 48 h (Figure 1) the expression levels of collagen III and fibronectin mRNA of the CTGF group were greater than those of the control group, and the differences were statistically significant (P=0.012). The expression levels of collagen III and fibronectin mRNA of the CP group, the CL group and the CPL group were lower by various degrees than those of the CTGF group. Among these differences, the difference between CPL and CTGF had the highest significance, and all the differences were statistically significant (Table 2).
Electrophoresis of Collagen III, Fibronectin. A: Electrophoresis of Collagen III gene (174 bp). B: Electrophoresis of Fibronectin gene (153 bp). C: Electrophoresis of internal standard (111 bp). M: Marker; 1: control group; 2: CTGF group; 3: CP group; 4: CL group; 5: CPL group.
Expression of collagen III protein
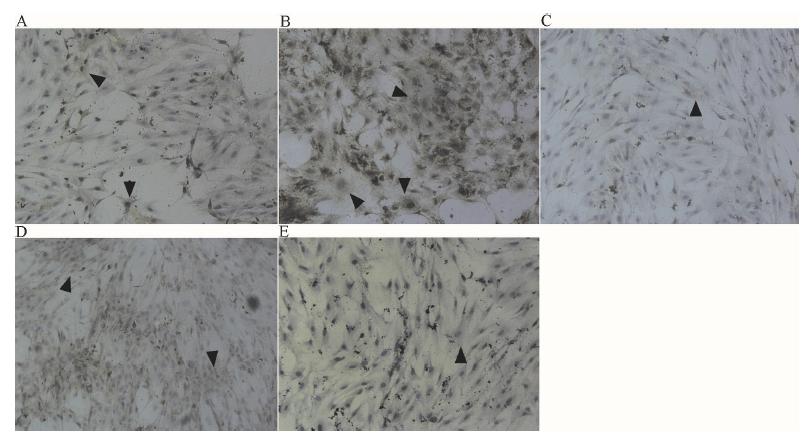
The immunohistochemical staining (Figure 2) showed that, after a 72-h intervention, the cytoplasmic staining of the CTGF group was much greater than that of the control group, while the R value was significantly decreased and the difference was statistically significant (P=0.0231; Table 3). Secondly, the cytoplasmic staining of the PD98059 and LY294002 groups was slightly greater than that of the CTGF group, while that of the CPL group was slightly lower. Here the R values were significantly higher, and the differences were statistically significant (P=0.013).
Immunohistochemical detection of Collagen III protein after slice climbing. A: Control group; B: CTGF group; C: CP group; D: CL group; E: CPL group. The positive and specific IHC signal of type III collagen were buffy in the cytoplasm of PASMCs convinced by the black arrows, ×400.
The Western-blot (Figure 3) showed that the specific protein was expressed near 130kDa in size, and the protein expression level of the CTGF group was significantly greater than in the control group (P=0.0249; Table 3). The protein expression levels of the CP and CL groups were significantly lower than those of the CTGF group, and the CPL group exhibited the lowest expression of collagen III protein; these differences were statistically significant (P=0.0187).
Discussion
PAH-PVR is characterized by the formation of neointimal growth, the proliferation and hypertrophy of PASMCs, and deposition of ECM, including the elastin, collagen and fibronectin1010 Warburton D, Shi W, Xu B. TGF-ß-Smad3 signaling in emphysema and pulmonary fibrosis: an epigenetic aberration of normal development? Am J Physiol Lung Cell Mol Physiol. 2013 Jan 15;304(2):L83-5. doi: 10.1152/ajplung.00258.2012.
https://doi.org/10.1152/ajplung.00258.20...
,1111 Romero D, Kawano Y, Bengoa N, Walker MM, Maltry N, Niehrs C, Waxman J, Kypta R. Down regulation of Dickkopf-3 disrupts prostate acinar morphogenesis through TGF-ß/Smad signalling. J Cell Sci. 2013 Apr 15;126(Pt 8):1858-67. doi: 10.1242/jcs.119388.
https://doi.org/10.1242/jcs.119388...
. Collagen and fibronectin are the important ECM components of the vascular wall. Collagen is an ECM component with multiple types, among which collagen I and III are the most important components in the skin, heart, blood vessels and other organs, maintaining their normal functioning. However, when the concentrations of these collagens are too high to be effectively decomposed by the body, they accumulate and cause the tissue fibration, which leads to the arterial hardening and, in severe cases, impairment of cardiopulmonary function, and ultimately causes loss of organ function. Fibronectin is a large glycoprotein and another important component of the ECM. In addition to fibroblasts, endothelial cells, macrophages, and smooth muscle cells, a number of epithelial cell types synthesize fibronectin. Certain fibronectins can bind with other ECM components (such as collagen and proteoglycans), thus making the ECM form a network. Other fibronectins can bind with cell surface receptors, making the cells adhere to the ECM. Furthermore, fibronectin also has important roles in cell expansion, migration and phenotypic changes.
The mechanisms of abnormal ECM deposition in PAH lung tissues are not yet clear. Previous studies have found that many factors play roles in the matrix deposition process, such as hypoxia11 Schuurhof A, Bont L, Hodemaekers HM, de Klerk A, de Groot H, Hofland RW, van de Pol AC, Kimpen JL, Janssen R. Proteins involved in extracellular matrix dynamics are associated with respiratory syncytial virus disease severity. Eur Respir J. 2012 Jun;39(6):1475-81. doi: 10.1183/09031936.00012311.
https://doi.org/10.1183/09031936.0001231...
,22 Kelvin EA, Edwards S, Jedrychowski W, Schleicher RL, Camann D, Tang D, Perera FP. Modulation of the effect of prenatal PAH exposure on PAH-DNA adducts in cord blood by plasma antioxidants. Cancer Epidemiol Biomarkers Prev. 2009 Aug;18(8):2262-8. doi: 10.1158/1055-9965.EPI-09-0316.
https://doi.org/10.1158/1055-9965.EPI-09...
, ANG II33 Pi L, Shenoy AK, Liu J, Kim S, Nelson N, Xia H, Hauswirth WW, Petersen BE, Schultz GS, Scott EW. CCN2/CTGFregulates neovessel formation via targeting structurally conserved cystine knot motifs in multiple angiogenic regulators. FASEB J. 2012 Aug;26(8):3365-79. doi: 10.1096/fj.11-200154.
https://doi.org/10.1096/fj.11-200154...
, ET-144 Sakai N, Chun J, Duffield JS, Wada T, Luster AD, Tager AM. LPA1-induced cytoskeleton reorganization drives fibrosis through CTGF- dependent fibroblast proliferation. FASEB J. 2013 May;27(5):1830-46. doi: 10.1096/fj.12-219378.
https://doi.org/10.1096/fj.12-219378...
, TGF-β1010 Warburton D, Shi W, Xu B. TGF-ß-Smad3 signaling in emphysema and pulmonary fibrosis: an epigenetic aberration of normal development? Am J Physiol Lung Cell Mol Physiol. 2013 Jan 15;304(2):L83-5. doi: 10.1152/ajplung.00258.2012.
https://doi.org/10.1152/ajplung.00258.20...
,1111 Romero D, Kawano Y, Bengoa N, Walker MM, Maltry N, Niehrs C, Waxman J, Kypta R. Down regulation of Dickkopf-3 disrupts prostate acinar morphogenesis through TGF-ß/Smad signalling. J Cell Sci. 2013 Apr 15;126(Pt 8):1858-67. doi: 10.1242/jcs.119388.
https://doi.org/10.1242/jcs.119388...
, IGF-11212 Majed BH, Khalil RA. Molecular mechanisms regulating the vascular prostacyclin pathways and their adaptation during pregnancy and in the newborn. Pharmacol Rev. 2012 Jul;64(3):540-82. doi: 10.1124/pr.111.004770.
https://doi.org/10.1124/pr.111.004770...
and TNF-α1313 Sokolove J, Strand V, Greenberg JD, Curtis JR, Kavanaugh A, Kremer JM, Anofrei A, Reed G, Calabrese L, Hooper M, Baumgartner S, Furst DE; CORRONA Investigators. Risk of elevated liver enzymes associated with TNF inhibitor utilisation in patients with rheumatoid arthritis. Ann Rheum Dis. 2010 Sep;69(9):1612-7. doi: 10.1136/ard.2009.112136.
https://doi.org/10.1136/ard.2009.112136...
, and that some of these factors can coordinate with each other1414 Rodríguez-Vita J, Sánchez-López E, Esteban V, Rupérez M, Egido J, Ruiz-Ortega M. Angiotensin II activates the Smad pathway in vascular smooth muscle cells by a transforming growth factor-beta-independent mechanism. Circulation. 2005 May 17;111(19):2509-17. PMID: 15883213.-1515 Chelladurai P, Seeger W, Pullamsetti SS. Matrix metalloproteinases and their inhibitors in pulmonary hypertension. Eur Respir J. 2012 Sep;40(3):766-82. doi: 10.1183/09031936.00209911.
https://doi.org/10.1183/09031936.0020991...
. These factors also play important roles in cell proliferation and migration, as well as the synthesis and secretion of ECM proteins in PAH-PVR. In our previous studies, we found that in PAH animal models the pulmonary artery wall and neointimae were significantly thicker, the concentration of pulmonary interstitial collagen was significantly higher, and that the expression of CTGF mRNA and protein was upregulated5. The results of this in vitro study also reveals that CTGF can increase the expression of collagen and fibronectin mRNA by PASMCs. In addition, CTGF was shown to exhibit different roles in different cells, such as by promoting the expression of fibronectin in cultured renal tubular epithelial cells while not affecting the expression of collagen I and III, whereas for renal interstitial fibroblasts (NRK-49F) CTGF increases the expression of collagen III and TSP-11515 Chelladurai P, Seeger W, Pullamsetti SS. Matrix metalloproteinases and their inhibitors in pulmonary hypertension. Eur Respir J. 2012 Sep;40(3):766-82. doi: 10.1183/09031936.00209911.
https://doi.org/10.1183/09031936.0020991...
. This study also found that CTGF can inhibit the degradation of human mesangial cellular matrix by increasing the expression of MMP-2, TIMP-1 and TIMP-3, and that anti-CTGF antibody can slow down the degradation of the matrix reduce resulting from the high sugar and TGF-β1616 Courboulin A, Barrier M, Perreault T, Bonnet P, Tremblay VL, Paulin R, Tremblay E, Lambert C, Jacob MH, Bonnet SN, Provencher S, Bonnet S. Plumbagin reverses proliferation and resistance to apoptosis in experimental PAH. Eur Respir J. 2012 Sep;40(3):618-29. doi: 10.1183/09031936.00084211.
https://doi.org/10.1183/09031936.0008421...
17 Li J, Liu NF, Wei Q. Effect of rosiglitazone on cardiac fibroblast proliferation, nitric oxide production and connective tissue growth factor expression induced by advanced glycation end-products. J Int Med Res. 2008 Mar-Apr;36(2):329-35. PMID: 18380944.-1818 Altuntas CZ, Daneshgari F, Izgi K, Bicer F, Ozer A, Sakalar C, Grimberg KO, Sayin I, Tuohy VK. Connective tissue and its growth factor CTGF distinguish the morphometric and molecular remodeling of the bladder in a model of neurogenic bladder. Am J Physiol Renal Physiol. 2012 Nov 1;303(9):F1363-9. doi: 10.1152/ajprenal.00273.2012.
https://doi.org/10.1152/ajprenal.00273.2...
. This indicates that CTGF can promote the occurrence and development of organ fibrosis through promoting ECM production and inhibiting ECM degradation. These findings suggest that CTGF may play an important role in the synthesis and secretion of ECM protein of PASMC treated with various pro-proliferation factors, as well as in the progression of PAH-PVR.
Currently, certain studies have found that CTGF has a role in promoting cell proliferation, migration and ECM deposition in various cell types, and the question of whether or not the CTGF ligand integrin is involved has become a research hotspot. CTGF can promote the expression of α5β1 on the cell surface and its adhesion with fibronectin1919 Weston BS, Wahab NA, Mason RM. CTGF mediatesTGF-beta induced fibronectin matrix deposition by up-regulating active alpha5betal integrin in human mesangial cells. J Am Soc Nephrol. 2003 Mar;14(3):601-10. PMID: 12595495.. The binding of α5β1 and fibronectin plays an important role in mediating ECM deposition. CTGF can increase its binding with fibronectin through its CT structural domain at the C-terminal, and integrin αVβ1 mediates this binding2020 Hoshijima M, Hattori T, Inoue M, Araki D, Hanagata H, Miyauchi A, Takigawa M. CT domain of CCN2/CTGF directly interacts with fibronectin and enhances cell adhesion of chondrocytes through integrin alpha5beta1. FEBS Lett. 2006 Feb 20;580(5):1376-82. doi: 10.1016/j.febslet.2006.01.061
https://doi.org/10.1016/j.febslet.2006.0...
. CTGF involves in the deposition of collagen when bound with the integrin α6β1 in structural domain 3 of fibroblast CTGF2121 Heng EC, Huang Y, Black SA Jr, Trackman PC. CCN2, connective tissue growth factor, stimulates collagen deposition by gingival fibroblasts via module 3 and alpha6- and beta1 integrins. J Cell Biochem. 2006 May 15;98(2):409-20. PMID: 16440322.. This finding suggests that some integrin receptors may be binding sites of CTGF. However, it is not yet clear how the signal transduction pathways are launched, and how to affect the abnormal deposition of ECM, after the binding of CTGF to integrin receptors of PASMCs.
The present study found that, when inhibitors of the ERK1/2 signaling pathway (PD98059) and of the PI3K/PKB signaling pathway (LY294002) acted on PASMCs, the mRNA expression levels of cellular collagen III and fibronectin were significantly lower than those of the CTGF group, as was the expression level of collagen III protein. Studies of cancer pathogenesis have shown that the activation of the ERK1/2 and PI3K/PKB signaling pathways plays an important role in the process of ECM deposition of turmor cells. Inhibitors such as PD98059 and LY294002 can promote apoptosis, reduce their biological activities and inhibit the conversion of cellular phenotype by inhibiting the deposition of ECM in malignant mesothelioma cells66 Sbroggiò M, Bertero A, Velasco S, Fusella F, De Blasio E, Bahou WF, Silengo L, Turco E, Brancaccio M, Tarone G. ERK1/2 activation in heart is controlled by melusin, focal adhesion kinase and the scaffold protein IQGAP1. J Cell Sci. 2011 Oct 15;124(Pt 20):3515-24. doi: 10.1242/jcs.091140.
https://doi.org/10.1242/jcs.091140...
,2222 Liu YY, Zhang X, Ringel MD, Jhiang SM. Modulation of sodium iodide symporter expression and function by LY294002, Akti-1/2 and Rapamycin in thyroid cells. Endocr Relat Cancer. 2012 May;19(3):291-304. doi: 10.1530/ ERC-11-0288.,2323 Collo G, Bono F, Cavalleri L, Plebani L, Mitola S, Merlo Pich E, Millan MJ, Zoli M, Maskos U, Spano P, Missale C. Nicotine-induced structural plasticity in mesencephalic dopaminergic neurons is mediated by dopamine D3 receptors and Akt-mTORC1 signaling. Mol Pharmacol. 2013 Jun;83(6):1176-89. doi: 10.1124/mol.113.084863.
https://doi.org/10.1124/mol.113.084863...
. Another study found that the increased expression of fibronectin protein in rat mesangial cells was related to glomerular fibrosis and sclerosis77 van de Laar L, van den Bosch A, Boonstra A, Binda RS, Buitenhuis M, Janssen HL, Coffer PJ, Woltman AM. PI3K-PKB hyperactivation augments human plasmacytoid dendritic cell development and function. Blood. 2012 Dec 13;120(25):4982-91. doi: 10.1182/blood-2012-02-413229.
https://doi.org/10.1182/blood-2012-02-41...
, and also closely related to the ERK1/2 signaling pathway88 Foster JG, Blunt MD, Carter E, Ward SG. Inhibition of PI3K signaling spurs new therapeutic opportunities in inflammatory/autoimmune diseases and hematological malignancies. Pharmacol Rev. 2012 Oct;64(4):1027-54. doi: 10.1124/pr.110.004051.
https://doi.org/10.1124/pr.110.004051...
. These findings strongly suggest that the activation of the ERK1/2 and PI3K/PKB signaling pathways play an important role in the CTGF-promoted deposition of ECM by PASMCs during PAH.
Conclusions
The results of this study suggest that CTGF can stimulate the gene expression of collagen III and fibronectin in PASMC ECM, which may be one of the important factors that promote the matrix deposition in PAH-PVR. PD98059 and LY294002 can inhibit the ERK1/2 and PI3K/PKB pathways, respectively, thus interfering with the biological effects of CTGF. The ERK1/2 and PI3K/PKB signaling pathways may be involved in the ECM deposition of PASMCs stimulated by CTGF, which may be a new target for the therapeutic interference of with PAH-PVR.
References
-
1Schuurhof A, Bont L, Hodemaekers HM, de Klerk A, de Groot H, Hofland RW, van de Pol AC, Kimpen JL, Janssen R. Proteins involved in extracellular matrix dynamics are associated with respiratory syncytial virus disease severity. Eur Respir J. 2012 Jun;39(6):1475-81. doi: 10.1183/09031936.00012311.
» https://doi.org/10.1183/09031936.00012311 -
2Kelvin EA, Edwards S, Jedrychowski W, Schleicher RL, Camann D, Tang D, Perera FP. Modulation of the effect of prenatal PAH exposure on PAH-DNA adducts in cord blood by plasma antioxidants. Cancer Epidemiol Biomarkers Prev. 2009 Aug;18(8):2262-8. doi: 10.1158/1055-9965.EPI-09-0316.
» https://doi.org/10.1158/1055-9965.EPI-09-0316 -
3Pi L, Shenoy AK, Liu J, Kim S, Nelson N, Xia H, Hauswirth WW, Petersen BE, Schultz GS, Scott EW. CCN2/CTGFregulates neovessel formation via targeting structurally conserved cystine knot motifs in multiple angiogenic regulators. FASEB J. 2012 Aug;26(8):3365-79. doi: 10.1096/fj.11-200154.
» https://doi.org/10.1096/fj.11-200154 -
4Sakai N, Chun J, Duffield JS, Wada T, Luster AD, Tager AM. LPA1-induced cytoskeleton reorganization drives fibrosis through CTGF- dependent fibroblast proliferation. FASEB J. 2013 May;27(5):1830-46. doi: 10.1096/fj.12-219378.
» https://doi.org/10.1096/fj.12-219378 -
5Liu B, Wang XM, Zhou TF, Hua YM, Liu HM, Wei L, Qiao LN, Wang XQ, Zhao SS, Shi K. Expression of connective tissue growth factor and its down-regulation by simvastatin administration in pulmonary hypertensive rats. Zhonghua Er Ke Za Zhi. 2008 May;46(5):359-65. PMID: 19099753.
-
6Sbroggiò M, Bertero A, Velasco S, Fusella F, De Blasio E, Bahou WF, Silengo L, Turco E, Brancaccio M, Tarone G. ERK1/2 activation in heart is controlled by melusin, focal adhesion kinase and the scaffold protein IQGAP1. J Cell Sci. 2011 Oct 15;124(Pt 20):3515-24. doi: 10.1242/jcs.091140.
» https://doi.org/10.1242/jcs.091140 -
7van de Laar L, van den Bosch A, Boonstra A, Binda RS, Buitenhuis M, Janssen HL, Coffer PJ, Woltman AM. PI3K-PKB hyperactivation augments human plasmacytoid dendritic cell development and function. Blood. 2012 Dec 13;120(25):4982-91. doi: 10.1182/blood-2012-02-413229.
» https://doi.org/10.1182/blood-2012-02-413229 -
8Foster JG, Blunt MD, Carter E, Ward SG. Inhibition of PI3K signaling spurs new therapeutic opportunities in inflammatory/autoimmune diseases and hematological malignancies. Pharmacol Rev. 2012 Oct;64(4):1027-54. doi: 10.1124/pr.110.004051.
» https://doi.org/10.1124/pr.110.004051 -
9Takei S, Hasegawa-Ishii S, Uekawa A, Chiba Y, Umegaki H, Hosokawa M, Woodward DF, Watanabe K, Shimada A. Immunohistochemical demonstra- tion of increased prostaglandin F2a levels in the rat hippocampus following kainic acid-induced seizures. Neuroscience. 2012 Aug;218:295-304. doi: 10.1016/j. neuroscience.2012.05.013.
» https://doi.org/10.1016/j -
10Warburton D, Shi W, Xu B. TGF-ß-Smad3 signaling in emphysema and pulmonary fibrosis: an epigenetic aberration of normal development? Am J Physiol Lung Cell Mol Physiol. 2013 Jan 15;304(2):L83-5. doi: 10.1152/ajplung.00258.2012.
» https://doi.org/10.1152/ajplung.00258.2012 -
11Romero D, Kawano Y, Bengoa N, Walker MM, Maltry N, Niehrs C, Waxman J, Kypta R. Down regulation of Dickkopf-3 disrupts prostate acinar morphogenesis through TGF-ß/Smad signalling. J Cell Sci. 2013 Apr 15;126(Pt 8):1858-67. doi: 10.1242/jcs.119388.
» https://doi.org/10.1242/jcs.119388 -
12Majed BH, Khalil RA. Molecular mechanisms regulating the vascular prostacyclin pathways and their adaptation during pregnancy and in the newborn. Pharmacol Rev. 2012 Jul;64(3):540-82. doi: 10.1124/pr.111.004770.
» https://doi.org/10.1124/pr.111.004770 -
13Sokolove J, Strand V, Greenberg JD, Curtis JR, Kavanaugh A, Kremer JM, Anofrei A, Reed G, Calabrese L, Hooper M, Baumgartner S, Furst DE; CORRONA Investigators. Risk of elevated liver enzymes associated with TNF inhibitor utilisation in patients with rheumatoid arthritis. Ann Rheum Dis. 2010 Sep;69(9):1612-7. doi: 10.1136/ard.2009.112136.
» https://doi.org/10.1136/ard.2009.112136 -
14Rodríguez-Vita J, Sánchez-López E, Esteban V, Rupérez M, Egido J, Ruiz-Ortega M. Angiotensin II activates the Smad pathway in vascular smooth muscle cells by a transforming growth factor-beta-independent mechanism. Circulation. 2005 May 17;111(19):2509-17. PMID: 15883213.
-
15Chelladurai P, Seeger W, Pullamsetti SS. Matrix metalloproteinases and their inhibitors in pulmonary hypertension. Eur Respir J. 2012 Sep;40(3):766-82. doi: 10.1183/09031936.00209911.
» https://doi.org/10.1183/09031936.00209911 -
16Courboulin A, Barrier M, Perreault T, Bonnet P, Tremblay VL, Paulin R, Tremblay E, Lambert C, Jacob MH, Bonnet SN, Provencher S, Bonnet S. Plumbagin reverses proliferation and resistance to apoptosis in experimental PAH. Eur Respir J. 2012 Sep;40(3):618-29. doi: 10.1183/09031936.00084211.
» https://doi.org/10.1183/09031936.00084211 -
17Li J, Liu NF, Wei Q. Effect of rosiglitazone on cardiac fibroblast proliferation, nitric oxide production and connective tissue growth factor expression induced by advanced glycation end-products. J Int Med Res. 2008 Mar-Apr;36(2):329-35. PMID: 18380944.
-
18Altuntas CZ, Daneshgari F, Izgi K, Bicer F, Ozer A, Sakalar C, Grimberg KO, Sayin I, Tuohy VK. Connective tissue and its growth factor CTGF distinguish the morphometric and molecular remodeling of the bladder in a model of neurogenic bladder. Am J Physiol Renal Physiol. 2012 Nov 1;303(9):F1363-9. doi: 10.1152/ajprenal.00273.2012.
» https://doi.org/10.1152/ajprenal.00273.2012 -
19Weston BS, Wahab NA, Mason RM. CTGF mediatesTGF-beta induced fibronectin matrix deposition by up-regulating active alpha5betal integrin in human mesangial cells. J Am Soc Nephrol. 2003 Mar;14(3):601-10. PMID: 12595495.
-
20Hoshijima M, Hattori T, Inoue M, Araki D, Hanagata H, Miyauchi A, Takigawa M. CT domain of CCN2/CTGF directly interacts with fibronectin and enhances cell adhesion of chondrocytes through integrin alpha5beta1. FEBS Lett. 2006 Feb 20;580(5):1376-82. doi: 10.1016/j.febslet.2006.01.061
» https://doi.org/10.1016/j.febslet.2006.01.061 -
21Heng EC, Huang Y, Black SA Jr, Trackman PC. CCN2, connective tissue growth factor, stimulates collagen deposition by gingival fibroblasts via module 3 and alpha6- and beta1 integrins. J Cell Biochem. 2006 May 15;98(2):409-20. PMID: 16440322.
-
22Liu YY, Zhang X, Ringel MD, Jhiang SM. Modulation of sodium iodide symporter expression and function by LY294002, Akti-1/2 and Rapamycin in thyroid cells. Endocr Relat Cancer. 2012 May;19(3):291-304. doi: 10.1530/ ERC-11-0288.
-
23Collo G, Bono F, Cavalleri L, Plebani L, Mitola S, Merlo Pich E, Millan MJ, Zoli M, Maskos U, Spano P, Missale C. Nicotine-induced structural plasticity in mesencephalic dopaminergic neurons is mediated by dopamine D3 receptors and Akt-mTORC1 signaling. Mol Pharmacol. 2013 Jun;83(6):1176-89. doi: 10.1124/mol.113.084863.
» https://doi.org/10.1124/mol.113.084863
-
Financial source:
Science and Technology Research Foundation of Sichuan Provincial Health Department (nº 100264) -
1
Research performed at Laboratory Center of Southwest Medical University, Luzhou, China. Part of Master degree thesis, Postgraduate Program in Pediatrics Cardiology. Tutor: Bin Liu.
Publication Dates
-
Publication in this collection
May 2017
History
-
Received
15 Jan 2017 -
Reviewed
18 Mar 2017 -
Accepted
19 Apr 2017