Abstract
Purpose:
To evaluate DNA damage levels in pregnant rats undergoing a treadmill exercise program.
Methods:
Wistar Kyoto rats were allocated into two groups (n= 5 animals/group): non-exercise and exercise. The pregnant rats were underwent an exercise protocol on a treadmill throughout pregnancy. Exercise intensity was set at 50% of maximal capacity during maximal exercise testing performed before mating. Body weight, blood pressure and glucose levels, and triglyceride concentration were measured during pregnancy. At day 10 post-natal, the animals were euthanized and maternal blood samples were collected for DNA damage.
Results:
Blood pressure and glucose levels and biochemical measurements showed no significant differences. Increased DNA damage levels were found in exercise group compared to those of non-exercise group (p<0.05).
Conclusion:
The exercise intensity protocol used in the study might have been exhaustive leading to maternal increased DNA damage levels, demonstrating the relevance of an adequate protocol of physical exercise.
Key words:
Pregnancy; Exercise; DNA Damage; Rats
Introduction
Animal studies have shown that normal fetal growth is critically dependent on adequate intrauterine environment because maternal metabolic abnormalities impair homeostasis, which might predispose development of disease in adulthood11 Holemans K, Aerts L, Van Assche FA. Fetal growth restriction and consequences for the offspring in animal models. J Soc Gynecol Invest. 2003;10:392-99. doi: 10.1080/jmf.11.4.254.257.
https://doi.org/10.1080/jmf.11.4.254.257...
.
Pregnancy is per se an inflammatory condition with increased susceptibility to oxidative stress22 Kontic-Vucinic O, Terzic M, Radunovic N. The role of antioxidant vitamins in hypertensive disorders of pregnancy. J Perinat Med. 2008;36(4):282-90. doi: 10.1515/JPM.2008.063.
https://doi.org/10.1515/JPM.2008.063...
. Reactive oxygen and nitrogen species, derived from the inflammatory process, can cause DNA damage that might affect several physiological processes during pregnancy. As repair of DNA damage is suppressed during pregnancy, pregnant women become more susceptible to environmental and endogenous agents33 Skoner JM, Sigmon J, Larcom LL. Suppressed DNA repair capacity of peripheral lymphocytes in pregnant women. Mol Cell Endocrinol. 1995;108(1-2):179-83. PMID: 7758833.. Furthermore, modern sedentary lifestyles can also cause excess inflammation and oxidative stress, which increases the risk of diseases and DNA damage44 Furnessa DLF, Dekkerb GA, Robertsa CT. DNA damage and health in pregnancy. J Reprod Immunol. 2011;89:153-62. doi: 10.1016/j.jri.2011.02.004.
https://doi.org/10.1016/j.jri.2011.02.00...
.
Physical activity brings health benefits, so pregnant women are advised to keep active during pregnancy, although a large number of them stop exercising when they are pregnant55 Melzer K, Schutz Y, Boulvain M, Kayser B. Physical activity and pregnancy: cardiovascular adaptations, recommendations and pregnancy outcomes. Sports Med. 2010;40(6):493-507. doi: 10.2165/11532290-000000000-00000.
https://doi.org/10.2165/11532290-0000000...
. Physical fitness and regular prenatal exercise can enhance placental growth and vascularity, decrease oxidative stress and reduce maternal endothelial dysfunction66 Weissgerber TL., Wolfe LA, Davies GA, Mottola MF. Exercise in the prevention and treatment of maternal-fetal disease: a review of the literature. Appl Physiol Nutr Metab. 2006;31(6):661-74. doi: 10.1139/h06-060.
https://doi.org/10.1139/h06-060...
. Nonetheless, the American College of Obstetricians and Gynecologists77 ACOG Practice Bulletin. Chronic hypertension in pregnancy. ACOG Committee on Practice Bulletins. Obstet Gynecol. 2001;98(1):177-85. PMID: 11508256. recommends that in order to ensure safety of both mother and fetus, exercise intensity should be closely monitored during pregnancy. Better benefits are directly associated with higher levels of physical activity, but depending on its intensity, exercise can either prevent or cause DNA damage88 Cash SW, Beresford SAA, Vaughan TL, Heagerty PJ, Bernstein L, White E, Neuhouser ML. Recent physical activity in relation to DNA damage and repair using the comet assay. J Phys Act Health. 2014;11(4):770-6. doi: 10.1123/jpah.2012-0278.
https://doi.org/10.1123/jpah.2012-0278...
. The underlying mechanisms are still unclear. Although some studies have evaluated the influence of exercise intensity on oxidative stress during pregnancy99 Matsuzaki M, Haruna M, Ota E, Murayama R, Yamaguchi T, Shioji I, Sasaki S, Yamaguchi T, Murashima S. Effects of lifestyle factors on urinary oxidative stress and serum antioxidant markers in pregnant Japanese women: a cohort study. Biosci Trends. 2014;8(3):176-84. PMID: 25030853., the effects of physical activity on DNA damage remain without further investigation. Thus, the aim of this study was to evaluate DNA damage levels in pregnant Wistar Kyoto rats under a treadmill exercise program.
Methods
The Ethics Committee for Animal Research of Botucatu Medical School-Universidade Estadual Paulista approved the protocols of this study (CEEA Number 889/2011).
Virgin female and male Wistar Kyoto (WKY) rats were acquired from the animal colony of Universidade de São Paulo and kept at the Laboratory of Experimental Research of the Department of Internal Medicine, Botucatu Medical School, UNESP, under controlled temperature (22 ± 2°C) and lighting (12h light/12h dark cycle) with free access to food and water.
Female rats were randomly assigned in two experimental groups (n=5 animals/group): non-exercise WKY - pregnant rats were submitted to no physical exercise, and exercise WKY- pregnant rats submitted to treadmill walking.
Adaptation to treadmill walking
At adult life (approximately at 80 days of life), the animals from the exercise group were familiarized with the exercise apparatus by being placed on the treadmill operating at zero speed as a mild electric stimulation and noise. Subsequently, familiarization to treadmill walking was performed at 5 meter (m)/minutes (min) for 10 min over a 1-week period. Electric stimulation was used to encourage and improve the ability to walk throughout the adaptation period. During pregnancy this stimulus was not carried out. Non-exercise rats were not submitted to a treadmill walking and were kept at the same inverted dark-light cycle to reproduce the circadian biological clock of animals .
Maximal exercise testing
Following familiarization to training, two days before mating, exercise animals were underwent to maximal exercise testing, which is widely used to determine exercise intensity in experimental studies1010 Rodrigues B, Figueroa DM, Mostarda CT, Heeren MV, Irigoyen MC, Angelis K. Maximal exercise test is a useful method for physical capacity and oxygen consumption determination in streptozotocin-diabetic rats. Cardiovasc Diabetol. 2007;6:38. doi: 10.1186/1475-2840-6-38.
https://doi.org/10.1186/1475-2840-6-38...
. The test consisted of graded exercise on the treadmill, starting at 5 km/h with increments of 0.3 km/h every 3 min up to exhaustion1111 Melo RM, Martinho E Jr, Michelini JC. Training-induced, pressure-lowering effect in SHR wide effects on circulatory profile of exercised and nonexercised muscles. Hypertension. 2003;42(2):851-7. doi: 10.1161/01.HYP.0000086201.27420.33.
https://doi.org/10.1161/01.HYP.000008620...
. Low intensity exercise was set at 50%-60% of the speed reached during maximal exercise test prior to pregnancy1010 Rodrigues B, Figueroa DM, Mostarda CT, Heeren MV, Irigoyen MC, Angelis K. Maximal exercise test is a useful method for physical capacity and oxygen consumption determination in streptozotocin-diabetic rats. Cardiovasc Diabetol. 2007;6:38. doi: 10.1186/1475-2840-6-38.
https://doi.org/10.1186/1475-2840-6-38...
.
Mating
At approximately 90 days of life, every morning males and females rats were put in breeding cages for mating. By late afternoon, the male was removed and vaginal smears were taken from the females for examination under a light microscope. The presence of spermatozoa in vaginal smears was positive mating indicator, and this was designated as day zero of pregnancy. Following, the animals were weighed, and the blood pressure was measured. The pregnant rats were housed in individual cages during whole pregnancy.
Treadmill exercise during pregnancy
The pregnant rats were submitted to a motorized treadmill started on day 1 of pregnancy at 10 m/min for 20 min, with increments of 10 min every two days up to 60 min. Thereafter, 60-minute sessions were held up to the end of pregnancy. Speed was gradually increased by 1m/min every day until maximum (13 m/min - determined by maximum exercise testing).
At pregnancy_maternal analysis
At days 0, 7, 14 and 21 of pregnancy, maternal body weight was assessed for weight gain evaluation, and systolic blood pressure level was measured in conscious rats by indirect tail-cuff plethysmography (LE5001 Pressure Meter; Panlab(r), Barcelona, Spain). At days 0, 14 and 20 of pregnancy, blood samples were obtained by tail cut for triglyceride level measurement (Accutrend(r) Plus, Roche Diagnostics GmbH, Mannheim, Germany) and blood glucose levels (One-Touch Ultra, Johnson & Johnson(r) ).
At day 10 post-partum_maternal analysis
At term pregnancy, the rats were closely monitored for newborn birth. Ten days after birth, mothers and pups were euthanized under sodium pentobartbital (Hypnol(r) , Cristália Chemicals, Brazil) and decapitated for collection of whole blood samples. 500µL were collected in EDTA vacutainer tubes and transferred to microtubes containing 400mL of RPMI 1640 medium and 100mL of dimethyl sulfoxide (DMSO, Sigma-Aldrich(r)). The samples were kept at -20°C for 24h, and then stored in a -80°C freezer. Thawing was achieved on a 37°C water bath, and pellets were washed with PBS. A 20-ml aliquot was mixed with 120ml low melting point (LMP) agarose, immersed into a lysis solution, electrophoresed, neutralized and stained for DNA damage assessment.
Comet assay
Chemicals
Normal Melting Point Agarose (NMPA) and Low Melting Point Agarose (LMPA)1212 Collins AR. The comet assay for DNA damage and repair: principles, applications, and limitations. Mol Biotechnol. 2004;26(3):249-61. doi: 10.1385/MB:26:3:249.
https://doi.org/10.1385/MB:26:3:249...
(Collins 2004), sodium chloride (NaCL), ethidium bromide, dimethyl sulfoxide (DMSO), sodium ethylene diamine tetra-acetic acid (EDTA), hydrochloric Acid (HCl), sodium hydroxide (NaOH), N-lauroyl sarcosine, Calcium (Ca2+) and Magnesium (Mg2+)-Free phosphate- buffered saline (PBS), Tris and Triton X-100.
Treatment with hydrogen peroxide (H 2 O 2 )
Treatment with H2O2 (400μM) was used to create positive controls, according to the protocol described by Blasiak et al.1313 Blasiak J, Arabski M, Krupa R,Wozniak K, Zadrozny M, Kaznicki J, Zurawska M, Drzewoski J. DNA damage and repair in type 2 diabetes mellitus. Mutat Res. 2004;554(1-2):297-304. doi: 10.1016/j.mrfmmm.2004.05.011.
https://doi.org/10.1016/j.mrfmmm.2004.05...
with modification regarding the molarity to the H2O2 solution (10μM).
Electrophoresis
The comet assay was conducted as described by Tice et al.1414 Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF. Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ Mol Mutagen2000;35:206-21. PMID: 10737956. with modification1515 Lima PHO, Damasceno DC, Sinzato YK, Souza MSS, Salvadori, Calderon IMP, Rudge MVC. Levels of DNA damage in blood leukocyte samples from non-diabetic and diabetic female rats and their fetuses exposed to air or cigarette smoke. Mutat Res. 2008;65:44-9. doi: 10.1016/j.mrgentox.2008.02.008.
https://doi.org/10.1016/j.mrgentox.2008....
. Slides with test-samples were placed in an electrophoresis chamber and incubated with NaOH/EDTA buffer solution for 20 min at 4oC. Electrophoresis was run at 25 volts (300 mA) for 30 min. Subsequently, the slides were neutralized in Tris (pH=7.5) for 15 min, fixed in 100% ethanol, and dried at room temperature.
Staining
All slides were stained with ethidium bromide (20mg/mL), coated with a coverslip, and immediately analyzed.
Slide analysis
Slide samples were examined considering 5 different classes of DNA damage according to a visual score system, on a 0-4 scale (0 = no damage, 4= maximum damage). A total of 100 nucleoids per sample were analyzed. Each nucleoid analyzed received a score ranging from zero to 4 according to its characteristics. In addition, comet observations were made at x400 magnification using a fluorescence microscope. One hundred nucleoids per sample (50/slide) were assessed by automated image analysis (Comet Assay IV, Perceptive Instruments, UK). Tail intensity (defined as the percentage of DNA in the tail) was used as a measure of DNA damage.
Statistical analysis
Analysis of variance (ANOVA) followed by Student's t test was used for the analysis of the study population. DNA damage levels were assessed by the Wilcoxon two-sample test. The level of statistical significance adopted was p<0.05
Results
During pregnancy, maternal body weight, systolic pressure, blood glucose and triglyceride levels, and heart weight showed no significant statistically difference between the exercise and the non-exercise groups (p>0.05). In addition, the mean number of live newborns of exercise group did no differ compared to that of non-exercise rats (Table 1).
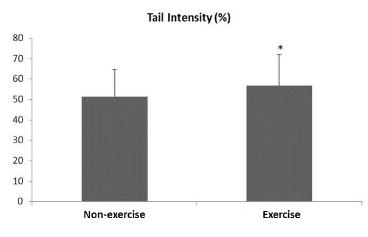
Figure 1 shows maternal DNA damage levels. The exercise group presented higher DNA damage levels compared to those of non-exercise rats (p<0.05). The DNA damage levels showed no significant correlation with body weight, systolic pressure, blood glucose and triglyceride levels) (p>0.05) (data not shown).
DNA damage levels of non-exercise and exercise Wistar Kyoto rats at day 10 post-partum. Data reported as mean ± standard deviation. *p<0.05 - compared to non-exercise group (Wilcoxon two-sample test).
Discussion
Physical training during pregnancy in laboratory animals has been shown to influence fetal growth and to improve maternal metabolism in models of voluntary exercise1616 Robinson AM, Bucci DJ. Physical exercise during pregnancy improves object recognition memory in adult offspring. Neuroscience. 2014;3;256:53-60. doi: 10.1016/j.neuroscience.2013.10.012.
https://doi.org/10.1016/j.neuroscience.2...
and swimming1717 Kim K, Chung E, Kim CJ, Lee S. Swimming exercise during pregnancy alleviates pregnancy-associated long-term memory impairment. Physiol Behav. 2012;107(1):82-6. doi: 10.1016/j.physbeh.2012.06.004.
https://doi.org/10.1016/j.physbeh.2012.0...
. However, such exercise models do not allow standardizing exercise intensity for animals, even if they are not pregnant. Therefore, the motorized treadmill has been used for establishing exercise duration/intensity, and the direct effect of the preferred intensity1818 Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
https://doi.org/10.1186/1472-6793-13-11...
.
Our results show that treadmill training did not affect pregnancy or fetal viability. Indeed, maternal body weight, systolic pressure, blood glucose and triglyceride levels showed no differences between experimental groups. Other studies have yielded conflicting results with regard to maternal body weight gain and fetal growth. These results might to be explained due to differences on exercise intensity, duration and type as well as to the presence/absence of preconditioning or familiarization with the type of exercise employed1919 Turgut S, Kaptanoglu B, Emmungil G, Turgut G. Increased plasma levels of growth hormone, insulin-like growth factor (IGF)-I and IGF-binding protein 3 in pregnant rats with exercise. Tohoku J Exp Med. 2006;208(1):75-81. PMID: 16340176.. Some animal studies have suggested that, in contrast to exercise of low to-moderate intensity, strenuous exercise may affect fetal and maternal body weight gain2020 Piçarro IC, Barros Neto TL, De Teves DC, Silva AC, Denadai DS, Tarasantchi J, Russo AK.. Effect of exercise during pregnancy, graded as a percentage of aerobic capacity: maternal and fetal responses of the rat. Comp Biochem Physiol A Comp Physiol. 1991;100(4):795-9. PMID: 1685371.. However, others have reported no effect on either maternal body weight gain2121 Nagai A, Sakamoto K, Konno K. The effect of maternal exercise on somatic growth and lung development of fetal rats: morphologic and morphometric studies. Pediatr Pulmonol. 1993;15(6):332-8. PMID: 8337010. or fetal body weight2222 Mottola MF, Bagnall KM, Belcastro AN. Effects of strenuous maternal exercise on fetal organ weights and skeletal muscle development in rats. J Dev Physiol. 1989;11(2):111-5. PMID: 2778291..
Although the exercise type used in our study did not affect the maternal reproductive and offspring number, there was an increased DNA damage levels in the exercise group. In this group, maximal exercise testing was performed before mating, indicating the estimated capacity for low intensity training before pregnancy might be different of that applied during pregnancy. According to Veja et al.2323 Veja CC, Reyes-Castro LA, Bautista CJ, Larrea F, Nathanielsz PW, Zambrano E. Exercise in obese female rats has beneficial effects on maternal and male and female offspring metabolism. Int J Obes (Lond). 2015 Apr;39(4):712-9. doi: 10.1038/ijo.2013.150.
https://doi.org/10.1038/ijo.2013.150...
, the pregnant rats submitted to voluntary exercise showed reduced physical capacity as pregnancy advanced. Thus, it is possible that the exercise intensity used in our study, which was classified as low at early pregnancy, was strenuous during pregnancy, and might therefore be linked to the increased maternal DNA damage levels. This hypothesis is supported by the fact that physical training of moderate intensity has been reported to provide protection against free radical generation in the long term due to an adaptive mechanism to meet energy demands24 that modulates oxidative responses, which become more effective with time. Nonetheless, in the event of overtraining, the homeostatic balance may be altered and to impair hormonal, metabolic and immune functions2525 Petibois C, Cazorla G, Poortmans JR, Déléris G. Biochemical aspects of overtraining in endurance sports: the metabolism alteration process syndrome. Sports Med. 2003;33(2):83-94. PMID: 12617688.. DNA damage levels do not increase after low-to-moderate physical exercise, but can be induced by strenuous exercise in rats1818 Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
https://doi.org/10.1186/1472-6793-13-11...
. Exhaustive exercise can lead to blood flow misdistribution, contributing to increased free radical production. Excess production of endogenous oxidants leads to tissue and DNA damage2626 Davison GW. Exercise and oxidative damage in nucleoid DNA quantified using single cell gel electrophoresis: present and future application. Front Physiol. 2016;7:249. doi: 10.3389/fphys.2016.00249.
https://doi.org/10.3389/fphys.2016.00249...
.
The increased levels of DNA damage observed in the exercise group have also been reported in other experiments. In a study of Sprague-Dawley rats exercising 30min/day on a treadmill for eight weeks, DNA damage was significantly higher as compared to sedentary controls. However, in rats receiving supplementation with vitamin E, exercise-induced DNA damage clearly was reduced, but it persisted higher than in sedentary animals2727 Hamid NAA, Hasrul MA, Ruzanna RJ, Ibrahim IA, Baruah PS, Mazlan M, Yusof YA, Ngah WZ. Effect of vitamin E (Tri E(r)) on antioxidant enzymes and DNA damage in rats following eight weeks exercise. Nutr J. 2011;10:37. doi: 10.1186/1475-2891-10-37.
https://doi.org/10.1186/1475-2891-10-37...
. Pereira et al.1818 Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
https://doi.org/10.1186/1472-6793-13-11...
examined the effects of aerobic training (60% maximal capacity) and overtraining (90% maximal capacity) in Swiss mice and found increased DNA damage in peripheral blood and skeletal muscle cells in the overtrained group as compared to the aerobic training group .
Strenuous exercise-induced DNA damage has been investigated with conflicting results. Our finding of increased DNA damage after exercising is consistent with those reported in human28 and animal studies1818 Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
https://doi.org/10.1186/1472-6793-13-11...
. Nonetheless, it is noteworthy that, in such studies, DNA damage levels were measured either immediately following exhaustive exercising or later. In the latter case, damage reversal was found to occur after 72h of rest2828 Hartmann A, Plappert U, Raddatz K, Grunert-Fuchs M, Speit, G. Does physical activity induce DNA damage? Mutagenesis. 1994;9:269-72. PMID: 7934967., whereas in our study DNA damage levels were still increased 10 days after exercising.
Tsai et al.29 assessing oxidative DNA damage in peripheral blood mononuclear cells from 12 male runners completing a 42-km marathon race (3h median running time) observed a significantly increased level of oxidative DNA damage that persisted for more than one week after the race. The authors pointed out the exact mechanisms underlying this phenomenon required elucidation as their findings, as well as ours, contradict other studies reporting decreased damage levels following a 72h post exercise rest1818 Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
https://doi.org/10.1186/1472-6793-13-11...
. According to those authors, the duration and extent of DNA effects seemed to increase synchronously with the amount and intensity of exercise. Furthermore, the fact that substantial increase in DNA damage still occurred in the subjects of their study, who undertook regular running training contradicts the notion that regular training confers protection against further exercise-induced DNA damage caused by adaptive responses2929 Tsai K, Hsu TG, Hsu KM, Cheng H, Liu TY, Hsu CF, Kong CW. Oxidative DNA damage in human peripheral leukocytes induced by massive aerobic exercise. Free Radic Biol Med. 2001;31(11):1465-72. PMID: 11728819..
Gandhi & Gunjan2626 Davison GW. Exercise and oxidative damage in nucleoid DNA quantified using single cell gel electrophoresis: present and future application. Front Physiol. 2016;7:249. doi: 10.3389/fphys.2016.00249.
https://doi.org/10.3389/fphys.2016.00249...
in their study review suggested the delayed onset of DNA damage might be explained by the fact that the initial injury is mechanical (muscular) in origin and results in physiological changes similar to those seen in acute inflammation, as well as in activation of polymorphonuclear neutrophils3030 Benoni G, Bellavite P, Adami A, Chirumbolo S, Lippi G, Cuzzolin LB. Effect of acute exercise on some haematological parameters and neutrophil functions in active and inactive subjects. Eur J Appl Physiol Occup Physiol. 1995;70:187-91. PMID: 7768243.. These polymorphonuclear neutrophils infiltrated in muscle damage sites release oxygen radicals and other reactive agents as part of the phagocytic process, leading to secondary tissue damage3131 Ebbeling CB, Clarkson PM. Exercise-induced muscle damage and adaptation. Sports Med. 1989;7:207-34. PMID: 2657962.. The infiltration of macrophages at the injury site rises steadily and is maintained through several days after exercise3232 Smith LL. Acute inflammation: the underlying mechanism in delayed onset muscle soreness? Med Sci Sports Exerc. 1991;23:542-51. PMID: 2072832..
Conclusions
The comet assay is a sensitive tool for assessing the effects of exercise during pregnancy on DNA damage and exercise has no adverse effects on both maternal and fetal health. In addition, no interference in body weight gain, pregnancy progress, delivery, and fetal viability were observed after treadmill training. However, an adequate intensity protocol is relevant to conduct an exercise program for laboratory animals and human.
Acknowledgements
The staff of Laboratory for Experimental Research in Gynecology and Obstetrics; to Mr. José Carlos Georgete, Mr. Mário Batista Bruno, Giovana Vesentini, Nathália Macedo and Thyego Santos for their technical contribution and to the Research Support Office of Botucatu Medical School, UNESP, especially Dr Jose Eduardo Corrente, for the statistical analysis.
References
-
1Holemans K, Aerts L, Van Assche FA. Fetal growth restriction and consequences for the offspring in animal models. J Soc Gynecol Invest. 2003;10:392-99. doi: 10.1080/jmf.11.4.254.257.
» https://doi.org/10.1080/jmf.11.4.254.257 -
2Kontic-Vucinic O, Terzic M, Radunovic N. The role of antioxidant vitamins in hypertensive disorders of pregnancy. J Perinat Med. 2008;36(4):282-90. doi: 10.1515/JPM.2008.063.
» https://doi.org/10.1515/JPM.2008.063 -
3Skoner JM, Sigmon J, Larcom LL. Suppressed DNA repair capacity of peripheral lymphocytes in pregnant women. Mol Cell Endocrinol. 1995;108(1-2):179-83. PMID: 7758833.
-
4Furnessa DLF, Dekkerb GA, Robertsa CT. DNA damage and health in pregnancy. J Reprod Immunol. 2011;89:153-62. doi: 10.1016/j.jri.2011.02.004.
» https://doi.org/10.1016/j.jri.2011.02.004 -
5Melzer K, Schutz Y, Boulvain M, Kayser B. Physical activity and pregnancy: cardiovascular adaptations, recommendations and pregnancy outcomes. Sports Med. 2010;40(6):493-507. doi: 10.2165/11532290-000000000-00000.
» https://doi.org/10.2165/11532290-000000000-00000 -
6Weissgerber TL., Wolfe LA, Davies GA, Mottola MF. Exercise in the prevention and treatment of maternal-fetal disease: a review of the literature. Appl Physiol Nutr Metab. 2006;31(6):661-74. doi: 10.1139/h06-060.
» https://doi.org/10.1139/h06-060 -
7ACOG Practice Bulletin. Chronic hypertension in pregnancy. ACOG Committee on Practice Bulletins. Obstet Gynecol. 2001;98(1):177-85. PMID: 11508256.
-
8Cash SW, Beresford SAA, Vaughan TL, Heagerty PJ, Bernstein L, White E, Neuhouser ML. Recent physical activity in relation to DNA damage and repair using the comet assay. J Phys Act Health. 2014;11(4):770-6. doi: 10.1123/jpah.2012-0278.
» https://doi.org/10.1123/jpah.2012-0278 -
9Matsuzaki M, Haruna M, Ota E, Murayama R, Yamaguchi T, Shioji I, Sasaki S, Yamaguchi T, Murashima S. Effects of lifestyle factors on urinary oxidative stress and serum antioxidant markers in pregnant Japanese women: a cohort study. Biosci Trends. 2014;8(3):176-84. PMID: 25030853.
-
10Rodrigues B, Figueroa DM, Mostarda CT, Heeren MV, Irigoyen MC, Angelis K. Maximal exercise test is a useful method for physical capacity and oxygen consumption determination in streptozotocin-diabetic rats. Cardiovasc Diabetol. 2007;6:38. doi: 10.1186/1475-2840-6-38.
» https://doi.org/10.1186/1475-2840-6-38 -
11Melo RM, Martinho E Jr, Michelini JC. Training-induced, pressure-lowering effect in SHR wide effects on circulatory profile of exercised and nonexercised muscles. Hypertension. 2003;42(2):851-7. doi: 10.1161/01.HYP.0000086201.27420.33.
» https://doi.org/10.1161/01.HYP.0000086201.27420.33 -
12Collins AR. The comet assay for DNA damage and repair: principles, applications, and limitations. Mol Biotechnol. 2004;26(3):249-61. doi: 10.1385/MB:26:3:249.
» https://doi.org/10.1385/MB:26:3:249 -
13Blasiak J, Arabski M, Krupa R,Wozniak K, Zadrozny M, Kaznicki J, Zurawska M, Drzewoski J. DNA damage and repair in type 2 diabetes mellitus. Mutat Res. 2004;554(1-2):297-304. doi: 10.1016/j.mrfmmm.2004.05.011.
» https://doi.org/10.1016/j.mrfmmm.2004.05.011 -
14Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF. Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ Mol Mutagen2000;35:206-21. PMID: 10737956.
-
15Lima PHO, Damasceno DC, Sinzato YK, Souza MSS, Salvadori, Calderon IMP, Rudge MVC. Levels of DNA damage in blood leukocyte samples from non-diabetic and diabetic female rats and their fetuses exposed to air or cigarette smoke. Mutat Res. 2008;65:44-9. doi: 10.1016/j.mrgentox.2008.02.008.
» https://doi.org/10.1016/j.mrgentox.2008.02.008 -
16Robinson AM, Bucci DJ. Physical exercise during pregnancy improves object recognition memory in adult offspring. Neuroscience. 2014;3;256:53-60. doi: 10.1016/j.neuroscience.2013.10.012.
» https://doi.org/10.1016/j.neuroscience.2013.10.012 -
17Kim K, Chung E, Kim CJ, Lee S. Swimming exercise during pregnancy alleviates pregnancy-associated long-term memory impairment. Physiol Behav. 2012;107(1):82-6. doi: 10.1016/j.physbeh.2012.06.004.
» https://doi.org/10.1016/j.physbeh.2012.06.004 -
18Pereira BC, Pauli JR, Antunes LM, de Freitas EC, de Almeida MR, de Paula Venâncio V, Ropelle ER, de Souza CT, Cintra DE, Papoti M, da Silva AS. Overtraining is associated with DNA damage in blood and skeletal muscle cells of Swiss mice. BMC Physiol. 2013;13:11. doi: 10.1186/1472-6793-13-11.
» https://doi.org/10.1186/1472-6793-13-11 -
19Turgut S, Kaptanoglu B, Emmungil G, Turgut G. Increased plasma levels of growth hormone, insulin-like growth factor (IGF)-I and IGF-binding protein 3 in pregnant rats with exercise. Tohoku J Exp Med. 2006;208(1):75-81. PMID: 16340176.
-
20Piçarro IC, Barros Neto TL, De Teves DC, Silva AC, Denadai DS, Tarasantchi J, Russo AK.. Effect of exercise during pregnancy, graded as a percentage of aerobic capacity: maternal and fetal responses of the rat. Comp Biochem Physiol A Comp Physiol. 1991;100(4):795-9. PMID: 1685371.
-
21Nagai A, Sakamoto K, Konno K. The effect of maternal exercise on somatic growth and lung development of fetal rats: morphologic and morphometric studies. Pediatr Pulmonol. 1993;15(6):332-8. PMID: 8337010.
-
22Mottola MF, Bagnall KM, Belcastro AN. Effects of strenuous maternal exercise on fetal organ weights and skeletal muscle development in rats. J Dev Physiol. 1989;11(2):111-5. PMID: 2778291.
-
23Veja CC, Reyes-Castro LA, Bautista CJ, Larrea F, Nathanielsz PW, Zambrano E. Exercise in obese female rats has beneficial effects on maternal and male and female offspring metabolism. Int J Obes (Lond). 2015 Apr;39(4):712-9. doi: 10.1038/ijo.2013.150.
» https://doi.org/10.1038/ijo.2013.150 -
24Møller P, Knudsen LE, Loft S, Wallin H. The comet assay as a rapid test in biomonitoring occupational exposure to DNA-damaging agents and effect of confounding factors. Cancer Epidemiol Biomarkers Prev. 2000;9(10):1005-15. PMID: 11045781.
-
25Petibois C, Cazorla G, Poortmans JR, Déléris G. Biochemical aspects of overtraining in endurance sports: the metabolism alteration process syndrome. Sports Med. 2003;33(2):83-94. PMID: 12617688.
-
26Davison GW. Exercise and oxidative damage in nucleoid DNA quantified using single cell gel electrophoresis: present and future application. Front Physiol. 2016;7:249. doi: 10.3389/fphys.2016.00249.
» https://doi.org/10.3389/fphys.2016.00249 -
27Hamid NAA, Hasrul MA, Ruzanna RJ, Ibrahim IA, Baruah PS, Mazlan M, Yusof YA, Ngah WZ. Effect of vitamin E (Tri E(r)) on antioxidant enzymes and DNA damage in rats following eight weeks exercise. Nutr J. 2011;10:37. doi: 10.1186/1475-2891-10-37.
» https://doi.org/10.1186/1475-2891-10-37 -
28Hartmann A, Plappert U, Raddatz K, Grunert-Fuchs M, Speit, G. Does physical activity induce DNA damage? Mutagenesis. 1994;9:269-72. PMID: 7934967.
-
29Tsai K, Hsu TG, Hsu KM, Cheng H, Liu TY, Hsu CF, Kong CW. Oxidative DNA damage in human peripheral leukocytes induced by massive aerobic exercise. Free Radic Biol Med. 2001;31(11):1465-72. PMID: 11728819.
-
30Benoni G, Bellavite P, Adami A, Chirumbolo S, Lippi G, Cuzzolin LB. Effect of acute exercise on some haematological parameters and neutrophil functions in active and inactive subjects. Eur J Appl Physiol Occup Physiol. 1995;70:187-91. PMID: 7768243.
-
31Ebbeling CB, Clarkson PM. Exercise-induced muscle damage and adaptation. Sports Med. 1989;7:207-34. PMID: 2657962.
-
32Smith LL. Acute inflammation: the underlying mechanism in delayed onset muscle soreness? Med Sci Sports Exerc. 1991;23:542-51. PMID: 2072832.
-
Financial source:
CAPES -
1
Research performed at Laboratory of Experimental Research on Gynecology and Obstetrics, Department of Gynecology and Obstetrics, Botucatu Medical School, Universidade Estadual Paulista (UNESP), Botucatu-SP, Brazil. Part of Master degree thesis, Postgraduate Program in Ginecology, Obstetrics and Mastology. Tutor: Prof. José Carlos Peraçoli.
Publication Dates
-
Publication in this collection
May 2017
History
-
Received
09 Jan 2017 -
Reviewed
08 Mar 2017 -
Accepted
10 Apr 2017