Resumos
RESUMO - Os mecanismos de patogenicidade e efeitos dos molicutes (espiroplasma e fitoplasma) na fisiologia de plantas de milho são pouco conhecidos. Com esse objetivo, avaliaram-se em casa de vegetação, características fisiológicas de plantas de milho submetidas ou não à inoculação com molicutes e cultivadas sob diferentes níveis de água no solo, correspondentes à reposição de 40, 60, 80 e 100% da disponibilidade total de água. Cada parcela experimental foi constituída por um vaso com duas plantas e repetida 6 vezes. Para inoculação, cigarrinhas Dalbulus maidis, vetor dos patógenos, coletadas em campo, foram confinadas nas plântulas por 7 dias. Foram avaliados os sintomas das doenças causadas por esses patógenos nas plantas de milho, sendo feita detecção de fitoplasma por PCR e de espiroplasma por Western blotting. A inoculação aumentou significativamente o potencial hídrico e a quantidade de água por unidade de matéria seca nas plantas. Esse efeito pode ser provavelmente atribuído à interferência dos patógenos na fisiologia das plantas, mediante produção de substâncias de crescimento, causando aumento na resistência estomática, redução na transpiração e conseqüentemente maior retenção de água nos tecidos. Esse mecanismo poderia evitar alterações na concentração osmótica das células, que são prejudiciais ao desenvolvimento desses procariontes sem parede celular. A inoculação aumentou também a proliferação de espigas, efeito geralmente causado pelo desbalanço hormonal, e reduziu a área foliar, altura da planta, matéria seca da planta e de grãos.
Potencial hídrico; Zea mays L.; encurtamento de internódios
Very few studies can be found concerning the mechanisms of pathogenicity and the effects of Mollicutes (spiroplasma and phytoplasma) on maize plants physiology. With this objective it was evaluated in greenhouse, physiological characteristics of plants submitted or not to inoculation with mollicutes and cultivated under different water levels in soil, corresponding to the reposition of 40, 60, 80 and 100% of the total water availability. Each experimental plot was set up with a pot contained two plants and repeated six times. In order to accomplish the inoculation, the pathogens vectors leaf hoppers Dalbulus maydis, were collected at the field and confined on the seedlings by seven days. The plants symptoms were evaluated, as well as the detection of phytoplasma by PCR and spiroplasma by Western blotting. Inoculation increased significantly the leaf water potential and the amount of water by unity of dry matter in the plants. This effect is probably due to the interference of pathogens on plant physiology, by producing growth substances, which caused increment on stomatal resistance, leaf transpiration reduction and consequently greater water retention in the tissues. This mechanism could avoid alterations on osmotical concentrations of cells which affects the development of those procaryotes without cell wall. The inoculation increased also the number of ears, which is generally caused by hormonal imbalance and reduced the leaf area, plant height, plant dry weight and final grain weight.
Water potential; Zea mays L.; internode shortage
ASPECTOS FISIOLÓGICOS DE PLANTAS DE MILHO INFECTADAS POR MOLICUTES SOB DIFERENTES NÍVEIS DE ÁGUA NO SOLO
PAULO CÉSAR MAGALHÃES1 1 Recebido: 30/10/2000 Aceito: 14/9/2001 . Engenheiro Agrônomo, PhD, Pesquisador Embrapa Milho e Sorgo, Caixa postal 151, 35701-970 Sete Lagoas MG. E-mail: pcesar@cnpms.embrapa.br (autor para correspondência). , ELIZABETH DE OLIVEIRA1 1 Recebido: 30/10/2000 Aceito: 14/9/2001 . Engenheiro Agrônomo, PhD, Pesquisador Embrapa Milho e Sorgo, Caixa postal 151, 35701-970 Sete Lagoas MG. E-mail: pcesar@cnpms.embrapa.br (autor para correspondência). , REINALDO L. GOMIDE1 1 Recebido: 30/10/2000 Aceito: 14/9/2001 . Engenheiro Agrônomo, PhD, Pesquisador Embrapa Milho e Sorgo, Caixa postal 151, 35701-970 Sete Lagoas MG. E-mail: pcesar@cnpms.embrapa.br (autor para correspondência). , CARLOS ALBERTO VASCONCELOS1 1 Recebido: 30/10/2000 Aceito: 14/9/2001 . Engenheiro Agrônomo, PhD, Pesquisador Embrapa Milho e Sorgo, Caixa postal 151, 35701-970 Sete Lagoas MG. E-mail: pcesar@cnpms.embrapa.br (autor para correspondência). , ISABEL REGINA P. SOUZA1 1 Recebido: 30/10/2000 Aceito: 14/9/2001 . Engenheiro Agrônomo, PhD, Pesquisador Embrapa Milho e Sorgo, Caixa postal 151, 35701-970 Sete Lagoas MG. E-mail: pcesar@cnpms.embrapa.br (autor para correspondência).
Embrapa Milho e Sorgo Cx. P. 151 Sete Lagoas MG 35701-970
RESUMO - Os mecanismos de patogenicidade e efeitos dos molicutes (espiroplasma e fitoplasma) na fisiologia de plantas de milho são pouco conhecidos. Com esse objetivo, avaliaram-se em casa de vegetação, características fisiológicas de plantas de milho submetidas ou não à inoculação com molicutes e cultivadas sob diferentes níveis de água no solo, correspondentes à reposição de 40, 60, 80 e 100% da disponibilidade total de água. Cada parcela experimental foi constituída por um vaso com duas plantas e repetida 6 vezes. Para inoculação, cigarrinhas Dalbulus maidis, vetor dos patógenos, coletadas em campo, foram confinadas nas plântulas por 7 dias. Foram avaliados os sintomas das doenças causadas por esses patógenos nas plantas de milho, sendo feita detecção de fitoplasma por PCR e de espiroplasma por Western blotting. A inoculação aumentou significativamente o potencial hídrico e a quantidade de água por unidade de matéria seca nas plantas. Esse efeito pode ser provavelmente atribuído à interferência dos patógenos na fisiologia das plantas, mediante produção de substâncias de crescimento, causando aumento na resistência estomática, redução na transpiração e conseqüentemente maior retenção de água nos tecidos. Esse mecanismo poderia evitar alterações na concentração osmótica das células, que são prejudiciais ao desenvolvimento desses procariontes sem parede celular. A inoculação aumentou também a proliferação de espigas, efeito geralmente causado pelo desbalanço hormonal, e reduziu a área foliar, altura da planta, matéria seca da planta e de grãos.
TERMOS ADICIONAIS PARA INDEXAÇÃO: Potencial hídrico, Zea mays L., encurtamento de internódios.
PHYSIOLOGICAL ASPECTS OF MAIZE PLANTS INFECTED BY MOLLICUTES UNDER DIFFERENT WATER LEVELS IN SOIL
ABSTRACT - Very few studies can be found concerning the mechanisms of pathogenicity and the effects of Mollicutes (spiroplasma and phytoplasma) on maize plants physiology. With this objective it was evaluated in greenhouse, physiological characteristics of plants submitted or not to inoculation with mollicutes and cultivated under different water levels in soil, corresponding to the reposition of 40, 60, 80 and 100% of the total water availability. Each experimental plot was set up with a pot contained two plants and repeated six times. In order to accomplish the inoculation, the pathogens vectors leaf hoppers Dalbulus maydis, were collected at the field and confined on the seedlings by seven days. The plants symptoms were evaluated, as well as the detection of phytoplasma by PCR and spiroplasma by Western blotting. Inoculation increased significantly the leaf water potential and the amount of water by unity of dry matter in the plants. This effect is probably due to the interference of pathogens on plant physiology, by producing growth substances, which caused increment on stomatal resistance, leaf transpiration reduction and consequently greater water retention in the tissues. This mechanism could avoid alterations on osmotical concentrations of cells which affects the development of those procaryotes without cell wall. The inoculation increased also the number of ears, which is generally caused by hormonal imbalance and reduced the leaf area, plant height, plant dry weight and final grain weight.
ADDITIONAL INDEX TERMS: Water potential, Zea mays L., internode shortage.
INTRODUÇÃO
As doenças do milho causadas por molicutes (fitoplasma e espiroplasma) destacam-se em importância para a cultura em conseqüência da alta incidência e aos prejuízos que causam na produção de grãos (Massola, 1999). Esses patógenos infectam o floema das plantas, sendo transmitidos de forma persistente pela cigarrinha Dalbulus maidis (Nault, 1980). Ocorrem simultaneamente nas mesmas áreas e, freqüentemente, nas mesmas plantas (Oliveira et al. 1998).
De forma geral, os sintomas dessas doenças têm sido associados ao encurtamento de internódios, a alterações na coloração das folhas e à proliferação de espigas improdutivas (Nault, 1980), efeitos que evidenciam interferência desses patógenos na fisiologia das plantas de milho.
O efeito causado pelos molicutes no encurtamento dos internódios pode reduzir uma importante reserva de fotoassimilados, que normalmente é utilizada pela planta durante o período de enchimento de grãos. Essa reserva se localiza no colmo e é de fudamental importância para satisfazer a demanda da planta no período reprodutivo (Magalhães et al. 1998). O fato de os patógenos alterarem a coloração das folhas igualmente afeta o desenvolvimento e a produção de grãos, uma vez que as folhas são consideradas a principal fonte de fotoassimilados para a planta e o mais importante órgão fotossintetizante do milho (Magalhães e Jones 1990; Magalhães et al. 1995). Já a proliferação de espigas improdutivas causa um desbalanceamento na relação fonte/dreno da planta e uma competição entre drenos por fotoassimilados disponíveis (Magalhães et al. 1999).
Os efeitos dessas doenças em milho têm sido avaliados principalmente com relação às reduções que causam na produção de grãos (Massola, 1999). Porém, as interações dos patógenos com a planta hospedeira, em relação aos seus mecanismos de patogenicidade e aos mecanismos de defesa da planta, são ainda pouco conhecidas (Daniels, 1983). Também não são conhecidos os efeitos da disponibilidade de água no solo sobre o desenvolvimento das doenças causadas por molicutes, e que têm sido encontradas em alta incidência em áreas irrigadas (Oliveira et al. 1998). Portanto, é importante estudar essa interação água no solo x molicutes, na tentativa de se identificar os mecanismos utilizados pelas plantas de milho quando infectadas por fitoplasma e/ou por espiroplasma.
Neste trabalho, objetivou-se verificar os efeitos da infecção por molicutes e da disponibilidade de água no solo sobre características fisiológicas em plantas de milho.
MATERIAL E MÉTODOS
Foi conduzido um experimento, em casa-de-vegetação, em vasos contendo 18 Kg de latossolo vermelho-escuro, fase cerrado, com os seguintes atributos químicos: pHágua 6,6; Ca e Mg extraído com KCl 1N, respectivamente 6,78 e 0,87 cmolc kg-1, o K e o P extraído pelo Mehlich 1, respectivamente 65 e 8 mg kg-1, Vettori (1969). O experimento seguiu o esquema fatorial 2x4 com os tratamentos: inoculado; não inoculado e níveis de 40, 60, 80 e 100% da água disponível no solo. Cada parcela experimental foi constituída por um vaso com duas plantas e cada tratamento foi repetido seis vezes, em delineamento experimental inteiramente ao acaso. No plantio, usaram-se 98 mg kg-1 de N; 150 mg kg-1 de P2O5, 197 mg kg-1 de K2O e 10 mg kg-1 de Zn. A adubação de cobertura foi efetuada semanalmente aplicando-se o equivalente a 20 mg kg-1 de N com sulfato de amônio. Em cada vaso foram semeadas 5 sementes de milho pipoca, cultivar Pop Zélia e, após germinação, foi feita a inoculação dos molicutes. Para inoculação, foram confinadas, sob gaiolas de tela, em cada vaso, 10 espécimes de cigarrinha Dalbulus maidis, coletadas em campo cultivado com milho pipoca, com elevada incidência de enfezamentos. Após uma semana, foi feita pulverização com o inseticida DDVP para eliminação das cigarrinhas, retiradas as gaiolas de tela e feito desbaste deixando-se duas plantas por vaso. Aos 30 dias pós semeadura, foram estabelecidos os tratamentos com os diferentes níveis de água no solo, correspondentes à reposição de 40, 60, 80 e 100% da disponibilidade total de água. A determinação dos níveis de água foi feita com base no balanço de água do solo, obtido individualmente para cada vaso, com o auxílio de uma balança eletrônica digital, com capacidade máxima de 32000,0g e sensibilidade de 0,1g. As irrigações foram realizadas de modo a repor integralmente a água consumida pela cultura, no intervalo entre duas irrigações sucessivas, visando a elevar a umidade do solo até o respectivo nível de reposição hídrica de cada tratamento.
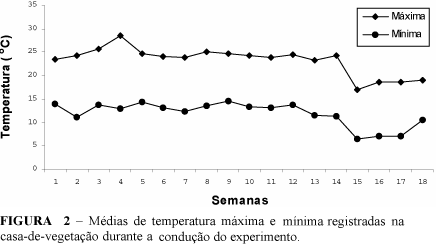
O experimento foi conduzido no período de abril a setembro, sendo a temperatura da casa-de-vegetação registrada em termohigrógrafo (Figura 2). Aos 60 dias após semeadura, uma das plantas de cada vaso foi utilizada para determinações de temperatura da folha, resistência estomática, transpiração, área foliar, altura da planta, potencial hídrico foliar e peso da matéria seca da parte aérea. A temperatura da folha, resistência estomática, e transpiração foliar foram determinadas com o uso de um porômetro LICOR 1600, (Steady State Porometer). A folha amostrada foi a primeira totalmente expandida, de cima para baixo, sendo as leituras realizadas sempre às 9 horas (horário solar) por ser esse considerado um horário em que a abertura estomática não era limitada pela luz e condições atmosféricas. O potencial hídrico foi determinado com a bomba de pressão de Schollander (Schollander et al. 1965). Para determinação do peso da matéria seca as plantas foram cortadas ao nível do solo, secas em estufa com ventilação forçada até peso constante a 75ºC. A outra planta foi cultivada até a produção, sendo avaliada aos 90 dias para determinações de: proliferação de espigas e peso de grãos. A manifestação dos sintomas de enfezamentos foi visualmente avaliada nas plantas aos 60 e aos 90 dias, sendo utilizada uma amostra de folha de cada uma para testes de detecção de fitoplasma e de espiroplasma.
A detecção de fitoplasma foi feita por PCR (Polymerase Chain Reaction), utilizando-se primers específicos para maize bushy stunt phytoplasma: MSB-F1 5AATGTCGAACTAA CAGGCGG e MBS-R1 3GGTTTTGGTTT AGGGGTT e condições para a reação descritas por Harrison et al. (1996). A detecção de espiroplasma foi feita por Western blotting, em membrana de nitrocelulose 0,45m (Hampton et al. 1990), utilizando-se IgG policlonal antiespiroplasma.
RESULTADOS E DISCUSSÃO
As plantas de milho submetidas à inoculação com molicutes apresentaram sintomas de enfezamento pálido e/ou de enfezamento vermelho, em intensidade variável. Particularmente após o florescimento, observaram-se reduções em crescimento, descolorações internervais e/ou avermelhamento das folhas e proliferação de espigas. Por meio do teste PCR, foi confirmada a presença de fitoplasma apenas nas plantas com sintomas acentuados do enfezamento vermelho. No caso do teste Western blotting, foi detectada a presença de espiroplasma apenas nas plantas com sintomas acentuados de enfezamento pálido. Nessas plantas constatou-se predominância de fitoplasma em relação ao espiroplasma. A limitação para detecção dos molicutes em todas as plantas apresentando sintomas pouco acentuados de enfezamentos pode ser provavelmente atribuída ao baixo título e à distribuição desuniforme dos patógenos. Limitações para detecção desses patógenos em plantas de milho que apresentavam sintomas de enfezamentos já foram observadas anteriormente (Oliveira et al. 1998).
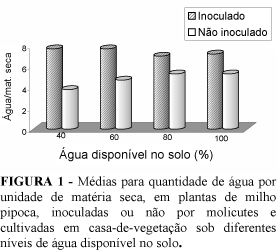
As plantas de milho com enfezamentos apresentaram maior potencial hídrico foliar (Tabela 1) e maior quantidade de água por unidade de matéria seca (Figura 1), independentemente do nível de água disponível no solo. Observa-se também na Tabela 1 que as plantas doentes apresentaram maior resistência estomática, o que pode explicar a menor transpiração e, conseqüentemente, o acúmulo de água nessas plantas. Esse fato pode ser possivelmente atribuído à interferência dos molicutes na fisiologia das plantas, de forma a manter a turgescência das células hospedeiras, sendo adequado à sua sobrevivência e multiplicação. Os molicutes são procariontes sem parede celular (Bascopé e Galindo 1981; Davis e Worley 1973) e, portanto, sensíveis a alterações na concentração osmótica do meio (Bascopé e Galindo 1981). Esse resultado chamou a atenção, uma vez que apesar de as plantas submetidas à inoculação estarem mais túrgidas, não tiveram maior crescimento e tampouco produziram mais grãos em relação às plantas sadias. Evidentemente, para plantas de milho sadias, fisiologicamente normais e apresentando maior potencial hídrico e maior turgescência, seria esperado maior produção de grãos em relação a plantas menos túrgidas, conforme já demonstrado em vários estudos (Boyer 1982, Taiz e Zeiger 1991). A presença dos molicutes foi o fator diferencial, pois, as plantas não utilizaram o potencial de água nos diversos processos de crescimento e desenvolvimento. Isso pode ser mais bem observado na Figura 1, por meio da qual verifica-se a quantidade de água/unidade de matéria seca nas plantas submetidas ou não a inoculação, nos 4 níveis de umidade do solo. Nota-se que praticamente não há diferença entre plantas doentes, com relação à quantidade de água por unidade de matéria seca, nos diferentes níveis de água disponível no solo ao passo que as plantas sadias mostram uma tendência de aumento na quantidade de água/matéria seca, à medida que aumenta a disponibilidade de água no solo.
A patogenese dos molicutes em outras espécies vegetais caracteriza-se por alterações na produção de substâncias de crescimento pelo hospedeiro (Chang 1998). O nível de ácido abscísico, em plantas de milho submetidas a condições de deficiência hídrica no solo, afeta diretamente a resistência estomática, e, conseqüentemente, a transpiração (Taiz e Zeiger 1991). Considerando-se esses aspectos, pode-se supor, então, que o espiroplasma e o fitoplasma patogênicos ao milho estejam causando aumento na concentração do ácido abscísico, resultando em aumento na resistência estomática, redução na transpiração e acúmulo de água nos tecidos.
Verifica-se ainda na Tabela 1 que a temperatura foliar não foi significativamente afetada pela doença. Ressalta-se, entretanto, que reduções na taxa de transpiração, que contribuam para o resfriamento foliar, podem causar aumento na temperatura da folha.
Outros resultados obtidos nesse estudo sugerem alterações hormonais nas plantas de milho, decorrentes da infecção por molicutes, como a redução em altura pelo encurtamento de internódios e a proliferação de espigas (Tabela 2). Chang (1998), estudando o efeito de molicutes em Catharanthus roseus, concluiu que a infecção por Spiroplasma citri reduziu a produção de substâncias de crescimento. É possível que efeitos semelhantes tenham ocorrido nas plantas de milho infectadas pelos molicutes. Alterações na produção de substâncias de crescimento podem ter causado a redução em altura das plantas infectadas. Por outro lado, a proliferação de espigas nessas plantas pode ter sido causada pelo desbalanço hormonal resultante da ação dos patógenos. O aumento no número de espigas improdutivas, além de causar um desbalanceamento da relação fonte/dreno, provoca uma queda na produtividade, uma vez que a maioria, se não todas, não terão seus grãos cheios, representando, assim, um gasto desnecessário de energia e uma competição entre drenos por fotoassimilados disponíveis (Magalhães et al. 1999). Foram observadas no presente trabalho várias plantas apresentando típico perfilhamento na base, provavelmente também causado por desbalanço hormonal.
A redução em altura é um dos fatores que pode contribuir de forma expressiva na disponibilização de fotoassimilados para o enchimento dos grãos. O colmo contém considerável reserva de fotoassimilados (Duncan et al. 1965; Tollenaar, e Daynard 1978 a), os quais podem ser translocados para a espiga, (Francis et al. 1978; Allison e Watson 1996, Jones e Simmons 1983). Há evidências de que uma mobilização considerável de fotoassimilados ocorra durante o período de enchimento dos grãos (Adelana e Milbourn 1972; Jones e Simmons 1983; Lucas 1981). Essa mobilização ocorre especialmente quando os requerimentos de carboidratos para os grãos excedem a produção de fotoassimilados pela planta (Tollenaar e Daynard 1978b). Portanto, o encurtamento dos internódios compromete o potencial dessa fonte extra de fotoassimilados localizada no colmo da planta de milho.
Notase que a inoculação reduziu significativamente a área foliar, a matéria seca da planta e a produção de grãos, especialmente no menor nível de água (Tabela 2). A folha é a principal fonte de fotoassimilados para a planta de milho (Magalhães et al. 1995) e redução na sua área pode comprometer decisivamente o desenvolvimento e a produção de grãos, uma vez que a área fotossinteticamente ativa é diminuída (Magalhães e Jones 1990). A redução na matéria seca da parte aérea e de grãos é uma conseqüência de todo o processo de crescimento e desenvolvimento da planta, comprometido pelo efeito da infecção pelos molicutes espiroplasma e fitoplasma (Tabela 2). As alterações na fisiologia das plantas de milho, causadas por esses patógenos, resultam em redução na absorção de N, P, K, Ca, Mg, Zn e S, como demonstrado por (Oliveira et al. 2001).
As médias de temperatura máxima registradas na casa-de-vegetação, durante o período de condução do experimento, variaram entre 16,9 e 28,600C e as médias de temperatura mínima entre 7,1 e 14,50C (Figura 2).
Essas condições de temperatura podem ser consideradas amenas em relação àquelas favoráveis ao desenvolvimento dos molicutes. Isso pode ter contribuído para amenizar os efeitos das doenças causadas por esses patógenos sobre o desenvolvimento das plantas de milho na casa-de-vegetação, em relação a condições de altas temperaturas, como aquelas que predominam em várias regiões do Brasil onde o milho é cultivado e essas doenças constituem problema (Oliveira et al. 1998).
- ADELANA, B.O. & MILBOURN, G.M. The growth of maize. II. Dry matter partition of three maize hybrids. Journal Agricultural Science, 78:73-78, 1972.
- ALLISON, J.C.S. & WATSON, D.J. The production and distribution of dry matter maize after flowering. Annual of Botany, 30:3655-381, 1966.
- BASCOPÉ, Q.B. & GALINDO A. J. Naturaleza micoplásmica de la raza mesa central del achaparramiento del maiz. Revista Fitopatologia, 16: 28-33, 1981.
- BOYER, J.S. Plant productivity and environment. Science, 218: 443-448, 1982.
- CHANG, J. C. Pathogenicity of Aster Yellow Phytoplasma and Spiroplasma citri on periwinkle. Phytopathology, 88: 1347-1350, 1998.
- DANIELS, M.J. Mechanisms of spiroplasma pathogenicity. Annual Review of Phytopathology,. 31: 29-43, 1983
- DAVIS, R.E. & WORLEY, J.F. Spiroplasma: motile, helical microorganism associated with corn stunt diseases. Phytopathology, 63: 403-408, 1973.
- DUNCAN. W.G.; HATFIELD, A.L. & RAGLAND, J.L. The growth and yield of corn. II. Daily growth of corn Kernels. Agronomy Journal, 57:221-223, 1965.
- FRANCIS, C.A.; TEMPLE, S.R.; FLOR, C.A. & GROGAN, C.L. Effects of competition on yield and dry matter distribution in maize. Field Crop Research, 1:51-63, 1978.
- GRANT, R. F.; JACKSON, B. S.; KINIRY, J. R. & ARKIN, G. F. Water deficit timing effects on yield components in maize. Agronomy Journal, 81: 61-65, 1989.
- HAMPTON,R., BALLE, E. & DE BOER, S. (Eds). Serological methods for detection and identification of viral and bacterial plant patogens A Laboratory Manual, St. Paul: APS press,1990. 384 p.
- HARRISON, N. A., RICHARDSON, P. A. & TSAI, J.H. PCR assay for detection of the phytoplasma associated with maize bushy stunt disease. Plant Disease, 80: 263-269, 1996.
- JONES, R.J. & SIMMONS, S.R. Effects of altered source sink relation on growth of maize kernels. Crop Science, 23: 129-134, 1983.
- LUCAS, E.O. Remobilization of stem assimilates in maize varieties grown under tropical conditions. Maydica, 26: 287-292, 1981.
- LUDLOW M. & MUCHOW, R. C. A critical evaluation of traits for improving crop yields in water-limited environments. Advance in Agronomy, 43: 107-153, 1990.
- MAGALHÃES P. C. & JONES, R. Aumento de fotoassimilados na taxa de crescimento e peso final dos grãos de milho. Pesquisa Agropecuária Brasileira, 25: 1747-1754, 1990.
- MAGALHÃES, P. C.; DURÃES, F.O.M. & PAIVA, E. Fisiologia da planta de milho Sete Lagoas, EMBRAPA-CNPMS, 1995. 27 p. (EMBRAPA-CNPMS. Circular Técnica, 20).
- MAGALHÃES, P. C.; DURÃES, F.O.M, & OLIVEIRA, A. C. Efeitos do quebramento do colmo no rendimento de grãos de milho. Ciência e Agrotecnologia, 22: 279-289, 1998.
- MAGALHÃES, P. C.; DURÃES, F.O.M; OLIVEIRA, A. C; & GAMA, E.E.G. Efeitos de diferentes técnicas de despendoamento na produção de milho. Scientia Agrícola, 56: 77-82, 1999.
- MASSOLA, JUNIOR, N.S.; BEDENDO, I. P.; AMORIM,L. & LOPES, J. R. S. Quantificação de danos causados pelo enfezamento vermelho e enfezamento pálido do milho em condições de campo. Fitopatologia Brasileira, 24: 136-142, 1999.
- NAULT, L.R. Maize bushy stunt and corn stunt: a comparison of disease symptoms, pathogen host ranges, and vectors. Phytopathology, 70: 659-662, 1980.
- OLIVEIRA, E.; WAQUIL, J.M.; FERNANDES, F.T.; PAIVA, E. RESENDE, R.O. & KITAJIMA, W.E. Enfezamento pálido e enfezamento vermelho na cultura do milho no Brasil Central. Fitopatologia Brasileira,.23: 45-47, 1998.
- OLIVEIRA, E.; MAGALHÃES, P.C.; GOMIDE, R.L.; VASCONCELOS, C.A.; SOUZA, I.R.P., OLIVEIRA, C. M. & CRUZ, I. Growth and nutrition of mollicute infected maize. Plant Disease, 2001. (No prelo).
- SCHOLLANDER, R. F.; HAMELL. H. T.; BRADSTREET, E. D. & HEMMINGSEN, E. A. Sap pressure in vascular plants. Science, 148: 339-346, 1965.
- TAIZ, L. & ZEIGER, E. Stress Physiology In: TAIZ, L. AND ZEIGER, E. Plant Physiology, California, The Benjamin / Cummings,. p. 346-370, 1991.
- TOLLENAAR. M. & DAYNARD, T.B. Effect of defoliation on Kernel development in maize. Canadian Journal of Plant Science, 58: 207.212, 1978a.
- TOLLENAAR. M. & DAYNARD, T.B. Relationship between assimilate source and reproductive sink in maize grown in a short season environment. Agronomy Journal, 70: 219-223, 1978b.
- VETTORI, L. Métodos da análise do solo . Rio de Janeiro, EPE, 1969. 24p ( Boletim Técnico, 7).
Datas de Publicação
-
Publicação nesta coleção
16 Jul 2002 -
Data do Fascículo
2001
Histórico
-
Aceito
14 Set 2001 -
Recebido
30 Out 2000