ARTIGOS ASSINADOS
Memórias
Ivan Izquierdo
O tempo flui, como um rio, aquele ao qual Heráclito disse que não podemos descer duas vezes. Há, basicamente, duas maneiras de conceber o fluxo do tempo: desde o passado em direção ao futuro, ou desde o futuro em direção ao passado (BORGES, 1960). Em qualquer um dos casos, o fluxo nos atravessa num ponto, que denominamos presente. Um ponto não tem superfície nem volume; é intangível e fugaz. É curioso que, em ambas concepções do tempo, o futuro (ou o passado) sejam conseqüências de algo quase imaterial como é o presente; de um simples ponto. Esse ponto evanescente, porém, é nossa única posse real: o futuro não existe ainda (e a palavra ainda é uma petição de princípio) e o passado não mais existe, salvo sob a forma de memórias. Não há tempo sem um conceito de memória; não há presente sem um conceito do tempo; não há realidade sem memória e sem uma noção de presente, passado e futuro.
Memória são as ruínas de Roma e as ruínas de nosso passado; memória tem o sistema imunológico, uma mola e um computador. Memória é nosso senso histórico e nosso senso de identidade pessoal (sou quem sou porque me lembro quem sou). Há algo em comum entre todas essas memórias: a conservação do passado através de imagens ou representações que podem ser evocadas. Representações, mas não realidades: as ruínas de Roma não são a Roma imperial; um disco da Nona Sinfonia gravado por Toscanini, Karayan ou Kleiber não eqüivale à sua execução, nem à Nona Sinfonia que Beethoven concebeu. Certamente não à que Beethoven tinha em mente quando, já totalmente surdo, a regeu pela primeira vez em Viena, em março de 1824: a orquestra já tinha concluído, há vários compassos, e o compositor, de olhos fechados, continuava regendo.
Quando se diz a palavra memória, a primeira que salta à evocação não é a memória das molas, dos discos ou dos computadores; é a memória das experiências individuais dos homens e dos animais, aquela que de alguma maneira se armazena no cérebro.
Desde um ponto de vista prático, a memória dos homens e dos animais é o armazenamento e evocação de informação adquirida através de experiências; a aquisição de memórias denomina-se aprendizado. As experiências são aqueles pontos intangíveis que chamamos presente.
Não há memória sem aprendizado, nem há aprendizado sem experiências. Aristóteles já disse, 2.000 anos atrás: "Nada há no intelecto que não tenha estado antes nos sentidos" (MARSHALL, 1988, p. 378). Não inventamos memórias. As memórias são fruto do que alguma vez percebemos ou sentimos. Os sonhos, que são em boa parte recombinações estranhas de memórias, provêm do que alguma vez sentimos ou percebemos (SELIGMAN, 1987, p. 46-53). Nossos planos e projetos (no fundo, uma variedade de sonhos), também. A palavra projeto vem de projetar; projetamos o passado, através do presente, em direção a um incerto futuro. Como a variedade e quantidade de experiências possíveis é enorme, a variedade de memórias possíveis é também enorme. Assim, talvez, não tenha muito sentido falar em "memória", senão em "memórias" (McGAUGH, 1988, p. 33-64). Cada uma pode ser diferente; e, em certo sentido, cada uma é diferente.
Não é possível medir memórias em forma direta (GREEN, 1964, p. 561-608). Só é possível avaliá-las medindo o desempenho em testes de evocação. Nos animais, a evocação se expressa através de mudanças comportamentais; no homem, a evocação pode também ser medida através do reconhecimento de pessoas, palavras, lugares ou fatos. É evidente que a evocação de uma alteração comportamental implica num prévio reconhecimento, e que todo reconhecimento implica numa alteração comportamental real ou potencial.
O aprendizado e a memória são propriedades básicas do sistema nervoso; não existe atividade nervosa que não inclua ou não seja afetada de alguma forma pelo aprendizado e pela memória. Aprendemos a caminhar, pensar, amar, imaginar, criar, fazer atos-motores ou ideativos simples e complexos, etc.; e nossa vida depende de que nos lembremos de tudo isso. PAVLOV (1960) e seus seguidores denominaram ao aprendizado e à memória "atividade nervosa superior".

A redução da memória a modelos, sem referencia a processos nervosos (HILGARD e BOWER, 1975; RESCORLA, 1985, p. 211-30), assim como sua redução a fenômenos puramente celulares, sem referência a processos cognitivos ou comportamentais, populares há alguns anos, tem hoje só interesse histórico (ROSE, 1988, p. 168-79; McGAUGH, 1988, p. 33-64; IZQUIERDO, 1989). Isto se deve a vários fatores. Um é o refinamento crescente das técnicas e dos conceitos neuropsicológicos que demonstraram que não é possível encaixar a enorme variedade de memórias possíveis dentro de um número limitado de esquemas ou modelos, nem reduzir seu alto grau de complexidade a mecanismos bioquímicos ou processos psicológicos únicos ou simples (MARKOWITSCH e PRITZEL, 1985, p. 189-287; IZQUIERDO, 1989). Foi-se a época em que se julgava possível explicar todos os aprendizados ou memórias por meio de reflexos condicionados, ou por meio de uma certa e determinada alteração molecular ou estrutural. Um fator importante, na queda dos modelos que pretendiam analisar a memória ignorando o sistema nervoso, é a reiterada demonstração de que é possível afetá-la profundamente, em forma às vezes muito específica, tanto por lesões nervosas precisas e localizadas (SCHÜTZ e IZQUIERDO, 1979, p. 97-105; MISHKIN et al., 1984, p. 65-77; THOMPSON, 1986, p. 941-47; SQUIRE, 1987), como por drogas ou substâncias endógenas cujo sítio e mecanismo de ação são conhecidos (IZQUIERDO, 1984 e 1989; McGAUGH, 1988, p. 33-64).
Porém, os mecanismos do armazenamento das memórias são hoje tão misteriosos como quando Ebbinghaus publicou a primeira análise experimental da memória, em 1885; ou como quando Müller e Pilzecker descreveram a consolidação da memória, em 1900. CASTELLANO (1987), numa metáfora muito vivida, comparou a memória com a luz das estrelas, "che può brillare ancora nella notte quando esse non esistono più".
GREEN (1964, p. 561-608) definiu a memória como um "estado" do cérebro que persiste além da estimulação sensorial e é capaz de influenciar sua atividade subseqüente. Disse que "o estado de memória não é acessível à investigação direta no presente momento"; porém muitas de suas conseqüências, sim; incluindo os fenômenos "transacionais" pelos quais esse "estado" pode ser adquirido ou evocado. Green sugeriu que o hipocampo pudesse estar envolvido nesses mecanismos transacionais, já que não diretamente no armazenamento. Como veremos, a evidência colhida nos 25 anos seguintes deram a razão a Green em mais de um aspecto.
A variedade de memórias possíveis é tão grande, que é evidente que a capacidade de adquirir, armazenar e evocar informações é inerente a muitas áreas ou subsistemas cerebrais, e não é função exclusiva de nenhuma delas. Aprendemos a não colocar os dedos na tomada, a reconhecer rostos ou lugares, a jogar futebol ou xadrez, a dirigir, a nos comportar nas circunstâncias mais diversas, aprendemos línguas, filosofias, medicina, música. É óbvio que diferentes sistemas sensoriais, associativos e motores participam em cada um destes aprendizados e nas correspondentes memórias. Usamos a via auditiva para aprender música, mas não para reconhecer um rosto; usamos o sistema-motor para aprender e evocar movimentos, mas não odores. Conseguimos evocar em contextos muito diferentes daqueles em que adquirimos cada informação. Aprendemos medicina nas aulas, rios livros, vendo preparados histológicos, ou estudando análises de sangue; evocamos essa informação junto a um paciente desconhecido, numa cama de hospital, ou ouvindo uma palestra em inglês, num país estrangeiro. Aprendemos tonotopias (por ex., a seqüência dó-mi-fá, numa escala muito grave, que é percebida só por certos neurônios) e podemos transpô-las, na evocação, a outros lugares da escala tonal (por ex., sol-si-dó, quatro oitavas acima, que é registrada por outros neurônios completamente diferentes ao longo da via auditiva). Podemos evocar essa tonotopia na imaginação, com a voz, ou com um violino; ou seja, utilizando neurônios totalmente distintos em cada caso. Aprendemos uma canção de Cole Porter ouvindo sua gravação pela orquestra de Nelson Riddle, na clave de sol, e podemos evocá-la no violão, com ritmo de samba, na clave de fá. Não parece digna de crédito a noção de que todas as memórias sejam explicáveis por um único e determinado processo molecular ou biofísico, nem pela constituição de novas vias nervosas ou novas sinapses ou alterações permanentes da condutância iônica em tal ou qual dendrito. Cada via nervosa é diferente, do ponto de vista dos neurotransmissores envolvidos, de suas conseqüências biofísicas, etc.; e é óbvio que algo aprendido usando determinada(s) via(s) pode ser evocado utilizando outras.
Algumas memórias consistem na inibição de respostas naturais ou inatas; outras, num aumento dessas respostas ou na geração de respostas novas; outras que não envolvem nenhuma resposta direta ou aparente. Há pacientes com falhas globais de memória, e outros com distúrbios limitados a uma única modalidade sensorial, ou ao reconhecimento de animais, por ex., mas não de objetos (MARSHALL, 1988, p. 378). Determinadas pessoas possuem uma excelente memória para números e não para faces; ou vice-versa. Tudo isto indica que diferentes memórias utilizam diferentes vias e processos tanto para sua aquisição como para sua evocação.
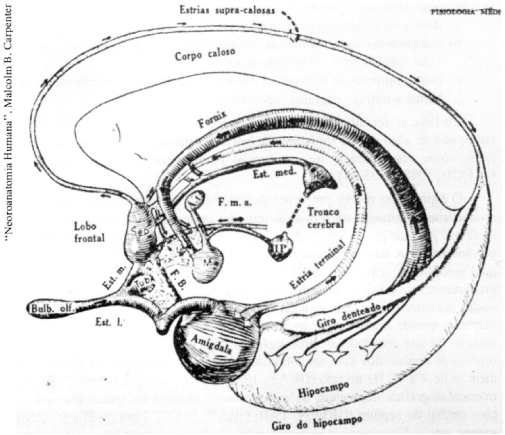
Há, não obstante, certas estruturas e vias (o hipocampo, a amígdala, e suas conexões com o hipotálamo e o tálamo) que regulam a gravação e evocação de todas, de muitas, ou pelo menos da maioria das memórias. Este conjunto de estruturas constitui um sistema modulador que influi na decisão, pelo sistema nervoso, ante cada experiência, de que deve ser gravado e de que deve ou pode ser evocado. O hipocampo e a amígdala estão interligados entre si e recebem informação de todos os sistemas sensoriais: em parte provenientes do córtex, e, em parte, de forma inespecífica quanto à modalidade sensorial, desde a formação reticular mesencefálica. O hipocampo e a amígdala projetam ao hipotálamo, e, através deste, ao tálamo e, finalmente, ao cortex (GREEN, 1964, p. 561-608; GRAY, 1982). Estas estruturas e suas conexões estão, portanto, estrategicamente localizadas para modular o processamento de informações baseadas na experiência.
Tipos de Memória
Há, talvez, tantos tipos de memória como tipos de experiência; não obstante, muitos as classificam de diversas formas. Existe um certo afã do homem em classificar, uma tendência enorme em fazê-lo constantemente, talvez como método básico para não se sentir tão profundamente ignorante. Ao classificar, reduzimos as coisas a nossa própria dimensão, não à dimensão que as coisas têm.

Classificar é, de fato, uma operação que implica em esquecer: esquecer diferenças para sustentar semelhanças (BORGES, 1960). Os orientais acham graça da tendência dos ocidentais em classificar coisas, e pôr um nome em tudo; será por isso, talvez, que em muitos lugares do Oriente não põem nome nem nas ruas (MIKES, 1970). Porém, a classificação mais ridícula que conheço é, precisamente, chinesa: a estranha classificação dos animais, constante do "Empório celestial de conhecimentos benévolos", citada por BORGES (1960):
a) pertencentes ao Imperador;
b) embalsamados;
c) amestrados;
d) leitões;
e) sereias;
f) fabulosos;
g) cachorros soltos;
h) incluídos nesta classificação;
i) que se agitam como se fossem malucos;
j) inumeráveis;
k) desenhados com um fino pincel de pêlo de camelo;
l) etcétera;
m) que vêm de quebrar um vaso;
n) que de longe parecem moscas.
As classificações da(s) memória(s) não são muito melhores.
Uma classificação habitual é de acordo com o tempo transcorrido entre sua aquisição e o momento em que são evocadas: memória imediata (segundos, minutos), recente (horas ou poucos dias), remota (semanas, meses, anos). A distinção é útil desde um ponto de vista descritivo e se usa em clínica; mas não há nenhuma evidência real de que esses tipos de memória representem processos diferentes. É provável que a distinção se deva a influências sobre a evocação de substâncias liberadas durante a aquisição b-endorfina, adrenalina, etc. (IZQUIERDO e NETTO, 1985b). É evidente que a memória imediata ocorre sob influência direta dessas substâncias; e que essa influência será menor quando medimos memória recente, minutos depois, e nula quando medimos memória remota, um mês/depois.
Outros autores preferem classificar as memórias de acordo com o tipo de informação: declarativa ("saber que") e de procedimentos ("saber como"); ou semântica (vgr., a linguagem, ou outros códigos) e episódica (memória de eventos ou episódios) (SQUIRE, 1987); ou em "hábitos" e "memórias" (MISHKBM et al., 1984, p. 65-77), como se um hábito não fosse, por definição, uma memória. Apesar de algumas tentativas iniciais (MISHKIN et al., 1984; SQUIRE, 1987), nunca foi realmente possível distinguir claramente entre estes tipos de memória. Uma memória declarativa (sabemos que pôr os dedos na tomada é perigoso) se converte, pela repetição, numa memória de procedimentos (automaticamente, sabemos como não colocar os dedos nas tomadas), ou num hábito. Um conhecimento semântico o inglês é adquirido em forma episódica (através de aulas) e pode ser evocado em forma episódica ou não. Por outro lado, não há evidência consistente de que os diversos tipos de memória correspondam a representações neuronais diferentes (ver o capítulo sobre amnésias).
Formação de Memórias
Para entender a formação de memórias a partir de experiências, é preciso considerar quatro aspectos fundamentais:
1) Recebemos informações constantemente, através de nossos sentidos; mas não memorizamos todas. Por ex., depois de ver um filme, lembramos algumas cenas; pode ser, até, muitas; mas não todas. Depois de ouvir uma aula, lembramos alguns conceitos; frases inteiras, talvez; mas não todos os conceitos nem todas as frases. Há, portanto, um processo de seleção prévio à formação de memórias, que determina quais informações serão armazenadas e quais não.
2) As memórias não são gravadas na sua forma definitiva, e são muito mais sensíveis à facilitação ou inibição logo após sua aquisição que em qualquer outro período posterior. Uma memória recente é muito mais suscetível ao efeito facilitador de certas drogas ou ao efeito amnésico de um traumatismo craniano que uma memória antiga (McGAUGH, 1988, p. 33-64). Isto indica que existe um processo de consolidação depois da aquisição (MÜLLER e PILZECKER, 1900, p. 1-288), pelo qual as memórias passam de um estado lábil a um estado estável.
3) As memórias são também muito mais sensíveis à incorporação de informação adicional nos primeiros minutos ou horas após a aquisição. Essa informação pode ser acrescentada, tanto por substâncias endógenas liberadas pela própria experiência bendorfina, adrenalina, etc. (IZQUIERDO, 1984, p. 65-77; IZQUIERDO, 1989), como por outras experiências que deixam memórias (LOFTUS e YUILLE, 1984; CAHILL et al., 1986; IZQUIERDO et al., 1988a,b; IZQUIERDO, 1989).
4) As memórias não consistem em itens isolados, senão em registros ("files") mais ou menos complexos. Não lembramos cada letra de cada palavra isoladamente; senão frases inteiras. Não lembramos cada cor ou cada odor percebido ontem como tais, senão como detalhes de "files" ou registros mais ou menos longos (o conjunto de eventos da hora do almoço; ou da tarde; ou do início da noite).
A formação ou não de uma memória depois de um determinado evento ou experiência, sua resistência à extinção, à interferência e ao esquecimento, dependem destes quatro fatores: seleção, consolidação, incorporação de mais informação, formação de registros ou "files".
Seleção
Os mecanismos que selecionam as informações que serão eventualmente armazenadas incluem o hipocampo e a amígdala. A lesão bilateral destas duas estruturas do lóbulo temporal não causa a perda de memórias preexistentes (o qual, obviamente, indica que as memórias não se armazenam nelas); mas impede a aquisição de novas memórias (SCHÜTZ e IZQUIERDO, 1979, p. 97-105; MISHKIN et al., 1984, p. 65-77; MARKOWITSCH e PRITZEL, 1985, p. 189-287).
O hipocampo intervém no reconhecimento de determinado estímulo, configuração de estímulos, ambiente ou situação, se são novos ou não, e, portanto, se merecem ou não ser memorizados (GRAY, 1982). É evidente que, para isto, o hipocampo deve ser capaz de:
a) distinguir estímulos, combinações de estímulos e ambientes;
b) compará-los com memórias preexistentes, armazenadas no cérebro (não, como vimos, no próprio hipocampo);
c) emitir informação referente à novidade ou não da situação ou do ambiente a outras estruturas (seus sítios de projeção).
De fato, só reconhecemos que "aprendemos algo" quando se trata de algo novo; não de algo que já sabíamos. A amígdala participa dos processos de seleção como conseqüência de sua função moduladora da consolidação (Mc-GAUGH, 1988, p. 33-64).
O hipocampo recebe projeções das diversas vias sensoriais através de vias secundárias originadas na formação reticular mesencefálica, septum, amígdala, e cortex. As vias provenientes da reticular, septum e amígdala contêm informação inespecífica, não identificada ainda pelo animal como pertencentes a tal ou qual modalidade sensorial (GREEN, 1964, p. 561-608; GRAY, 1982); as fibras provenientes do cortex, por outro lado, podem informar ao hipocampo se a ativação pertence a uma determinada modalidade sensorial, ou a várias. O hipocampo responde a ambientes, eventos ou estímulos novos com uma dessincronização de sua atividade electroencefalográfíca, e a ambientes, eventos ou estímulos já conhecidos com um ritmo lento chamado theta: de 4 a 7 Hz no homem, e de 4 a 10 Hz no rato (GRAY, 1982). Ambos os tipos de atividade electroencefalográfica dependem de marcapassos colinérgicos localizados no núcleo medial do septum (GREEN, 1964; GRAY, 1982). Muita evidência recente sugere que o hipocampo é especializado, de fato, no reconhecimento de ambientes, mais do que no reconhecimento de qualquer estímulo (O'KEEFE e NADEL, 1978; MORRIS et al., 1982, p. 681-83). A lesão do hipocampo afeta profundamente, e talvez especificamente, a capacidade de localização espacial (MORRIS et al., 1982). É provável que o reconhecimento da novidade ou não de determinadas ou de todas as situações em geral, por parte do hipocampo (GRAY, 1982), envolva, primariamente, o reconhecimento do contexto espacial de cada estímulo ou configuração de estímulos em particular (O'KEEFE e NADEL, 1978).
A ativação do hipocampo por experiências novas e/ou por ambientes novos e/ou por experiências dentro de ambientes novos tem duas conseqüências. Por um lado, o hipocampo informa a seus sítios de projeção: amígdala, hipotálamo, secundariamente através do hipotálamo a diversos núcleos talâmicos associativos (GRAY, 1982; WEISKRANTZ, 1985) que se trata realmente de uma experiência ou ambiente novo. Por outro lado, através do fórnix, o hipocampo ativa neurônios b-endorfinérgicos localizados no hipotálamo medial basal (IZQUIERDO e NETTO, 1985a). Estes neurônios, por sua vez, projetam ao septum medial (podendo, assim, fechar um círculo), à amígdala e a outros núcleos vinculados com a modulação dos processos de memória. A exposição a estímulos ou ambientes não-novidosos não é seguida de ativação do sistema b-endorfinérgico (IZQUIERDO e NETTO, 1985a; IZQUIERDO et al., 1988a; IZQUIERDO, 1989). A b-endorfina liberada pelas experiências novas incorpora informação às mesmas; pode-se, portanto, afirmar que os registros dessas experiências incluem o aprendizado respectivo somado à ação da b-endorfina (IZQUIERDO, 1984, 1989).
As memórias em que conhecimentos antigos são associados de maneira diferente se formam sem intervenção do hipocampo nem do sistema b-endorfínico. Por ex., todo animal sabe que ao sofrer um estimulo doloroso deve fugir ou reagir; aprende isso na sua infância, se já não nasce sabendo-o. Mas pode aprender a fugir melhor, ou a reagir mais conveniente ou rapidamente, perante a repetição do estímulo doloroso. Este tipo de aprendizado, que consiste no aperfeiçoamento ou no melhor uso de conhecimentos prévios, pode ser denominado "aprendizado de hábitos" (IZQUIERDO e NETTO, 1985b, p. 249-65) e é insensível a lesões hipocâmpicas e à ação da b-endorfina ou de antagonistas da b-endorfina.
Consolidação
Podemos perceber muitos estímulos ao mesmo tempo, em diversas combinações; e podemos até formar várias memórias novas simultaneamente (Id. ibid.); algumas, porém, serão melhor consolidadas que outras. Por ex., vemos várias pessoas que avançam em nossa direção; distinguimos seus rostos, expressões, roupas, gestos. Uma delas carrega uma arma. Gravaremos sua imagem vividamente; as das outras, não. A maior ou menor consolidação de determinadas experiências ou fragmentos de experiências, simultâneas ou consecutivas, influi grandemente no processo seletivo (McGAUGH, 1988). Uma memória bem-consolidada é difícil de extinguir (IZQUIERDO e PEREIRA, 1989); uma memória malconsolidada se extingue ou se esquece facilmente.
A consolidação é modulável. Os processos de modulação são importantes porque são muito melhor conhecidos que os sistemas de armazenamento ou evocação; e porque, de fato, a única forma disponível de afetar a memória quantitativamente, ou até qualitativamente, é através de variáveis que agem sobre os sistemas moduladores. As únicas bases biológicas da memória que conhecemos são, na verdade, as bases biológicas da modulação; e, embora não nos sirvam para entender como as memórias se armazenam, servem-nos para tratá-las quando se encontram diminuídas, como por ex., nos diversos tipos de amnésia.
As memórias adquiridas em estado de alerta e com certa carga emocional ou afetiva são melhor lembradas que as memórias de fatos inexpressivos ou adquiridas em estado de sonolência. Os estados de alerta, afetivos e emocionais se acompanham da liberação de hormônios periféricos e neurotransmissores centrais. Várias dessas substâncias afetam a memória. Numerosos experimentos com drogas que liberam, mimetizam ou bloqueiam sua ação demonstraram que as mesmas não atuam durante a aquisição, senão no período imediatamente posterior, afetando a consolidação (McGAUGH, 1988; IZQUIERDO et al., 1988a; IZQUIERDO e PEREIRA, 1989; IZQUIERDO, 1989). Os tratamentos são efetivos quando aplicados após a aquisição (no período denominado pós-treinamento).
Em nível central, os principais sistemas moduladores são o sistema colinérgico septo-hipocampal, o sistema colinérgico nucleus basalis-amígdala, e o sistema noradrenérgico locus ceruleus-amígdala. Existem projeções do sistema b-endorfínico ao septum, locus ceruleus e amígdala, assim como sinapses GABAérgicas na amígdala e no hipocampo. Há, portanto, possibilidade de interação de todos estes sistemas neuromoduladores em diversos níveis. Evidência recente indica que o principal sítio de interação é a amígdala (McGAUGH, 1988; IZQUIERDO, 1989).
A lesão bilateral da amígdala, a secção bilateral da stria terminalis (principal via aferente e eferente da amígdala), ou a injeção de bloqueadores b-noradrenérgicos na amígdala bloqueiam o efeito facilitador sobre a consolidação da adrenalina, de estimulantes colinérgicos, e de antagonistas da b-endorfina, dados após o treino. A lesão da stria terminalis bloqueia também o próprio efeito da b-endorfina. A injeção intra-amigdalina de bicuculina ou picrotoxina (bloqueadores do GABA) facilita a consolidação (McGAUGH, 1988). Sabe-se há anos que o diazepam, uma droga que potencia a transmissão GABAérgica, deprime a formação de novas memórias (CAHILL et al., 1986; IZQUIERDO et al., 1988a,b). A injeção de agonistas noradrenérgicos ou colinérgicos na amígdala facilita a consolidação (McGAUGH, 1988). A ativação moderada dos sistemas noradrenérgico, colinérgico e GABAérgico no período imediatamente posterior à aquisição de memórias parece, assim, necessária para que haja consolidação; e a amígdala parece ser o principal sítio de ação e interação destes sistemas entre si e com a b-endorfina (McGAUGH, 1988; IZQUIERDO, 1989).
Vários sistemas hormonais também modulam a consolidação. Os hormônios mais estudados são a adrenalina, adrenocorticotrofina e vasopressina, três hormônios liberados em resposta ao alerta e/ou ao estresse. A noradrenalina circulante, produzida pelo sistema simpático, é liberada geralmente ao mesmo tempo e/ou em resposta aos mesmos estímulos alertantes ou estressantes que a adrenalina da medula supra-renal e afeta a consolidação de forma similar. Uma hipersecreção moderada destas substâncias no período imediatamente posterior à aquisição, como a provocada por experiências moderadamente alertantes, ou a injeção de uma dose pequena das mesmas, facilita a consolidação. O grau de facilitação depende da concentração sangüínea e/ou da dose de cada um desses hormônios (GOLD e DELANOY, 1981, p. 79-98; McGAUGH, 1988; IZQUIERDO, 1989). Uma hipersecreção exagerada, ou a administração de uma dose alta, de qualquer uma dessas substâncias tem, pelo contrário, efeitos de-pressores que podem ser facilmente revertidos por uma nova injeção ou uma nova liberação das mesmas no momento da evocação (IZQUIERDO, 1984, p. 65-77; IZQUIERDO e McGAUGH, 1987, p. 778-81; IZQUIERDO et al., 1988a, p. 9-25). Os efeitos destes hormônios são mediados por neurônios noradrenérgicos na amígdala; a lesão deste núcleo, ou a injeção de bloqueadores b-adrenérgicos no mesmo, cancela os efeitos dos hormônios (McGAUGH, 1988). As vias reflexas, através das quais ações periféricas da adrenalina, adrenocorticotrofina ou vasopressina eventualmente afetam a atividade do núcleo amigdalino, não são conhecidas; só sabemos que essas vias incluem a formação reticular mesencefálica (GOLD e DELANOY, 1981; IZQUIERDO et al., 1988a; IZQUIERDO, 1989).
É importante salientar que a consolidação, mediada, como vimos, em boa parte por mecanismos b-noradrenérgicos na amígdala, é decisiva para estabelecer se uma determinada memória será gravada bem ou mal, e, portanto, se será forte ou fraca, e assim mais ou menos suscetível à perda, deterioração ou alteração no futuro. Uma consolidação fraca ou ineficiente causa amnésia anterógrada, como se observa, de fato, em pacientes ou animais com lesão bilateral da amígdala (SQUIRE, 1987). Se, à lesão da amígdala, acrescenta-se uma lesão hipocâmpica (MISHKIN et al., 1984; WEISKRANTZ, 1985; MARKOWITSCH e PRITZEL, 1985; SQUIRE, 1987), ou das projeções de ambas estruturas, o quadro é mais grave. O hipocampo, como vimos acima, é crucial para estabelecer se uma determinada memória é nova e deverá ser logo consolidada (gravada) ou não.
Incorporação de Informação Adicional: informação farmacológica
Enquanto estão sendo consolidadas, ou ainda depois, as memórias podem incorporar informação adicional.
Um tipo de informação adicional provém da ação da b-endorfina liberada por experiências novas (IZQUIERDO, 1984; IZQUIERDO e NETTO, 1985a,b; IZQUIERDO et al., 1988a,b). Esta se incorpora às memórias como mais um estímulo: a injeção intracerebral ou sistêmica de b-endorfina, ou uma segunda liberação cerebral da mesma por outra experiência nova, antes da evocação, tem um efeito facilitante similar àquele que teria a representação de qualquer estímulo próprio dessa tarefa (reminder effect) (IZQUIERDO e McGAUGH, 1985; IZQUIERDO e NETTO, 1985b). A administração de um excesso de (b-endorfina depois da adquisição requer a nova administração de uma quantidade maior de b-endorfina antes da evocação para que esta seja possível (IZQUIERDO e NETTO, 1985a).
No caso em que duas memórias são processadas ou adquiridas simultaneamente, uma envolvendo mais novidade do que a outra, a b-endorfina, miméticos da b-endorfina como a Leu-encefalina, liberadores da b-endorfina cerebral como o eletrochoque, e antagonistas da b-endorfina como a naloxona, só afetam a mais nova das duas (NETTO et al., 1986).

Doses ou concentrações sangüíneas elevadas de adrenalina, adrenocorticotrofina ou vasopressina no período posterior à adquisição, como vimos, podem também inibir a evocação. Seu efeito é também revertido por uma nova administração das mesmas substâncias antes da evocação (IZQUIERDO, 1984, p. 65-77; IZQUIERDO e McGAUGH, 1987, p. 778-81), o qual indica que, como a b-endorfina, estes hormônios, em doses altas, passam a se incorporar às diversas experiências como informação adicional, cuja reiteração no momento da evocação é necessária para que a mesma possa ter lugar (IZQUIERDO et al., 1988a,b; IZQUIERDO, 1989).
A incorporação de informação farmacológica às memórias ocorre só até os primeiros 5 a 10 minutos após cada experiência; a liberação ou injeção das substâncias além desse período não é mais efetiva (IZQUIERDO, 1984, 1989). A incorporação de informação farmacológica às memórias pode também ocorrer durante a adquisição (HO et al., 1978; IZQUIERDO, 1984). Diz-se, nesse caso, que a memória correspondente foi adquirida durante um estado induzido pela droga em questão; isto costuma fazer com que o sujeito precise voltar a esse estado, através de uma nova ingestão da droga, para poder evocar corretamente. Esta dependência de estados induzidos por drogas tem sido descrita, para uma grande variedade de fármacos de ação central, fundamentalmente drogas depressoras (HO et al., 1978).
Relações entre Consolidação e Incorporação de Informação Farmacológica
A consolidação e a incorporação de informação farmacológica às experiências (por ex., do efeito de uma injeção pós-treino de b-endorfina, ou de uma dose alta de adrenalina) coincidem no tempo. Os agentes que alteram a consolidação só são efetivos quando administrados nos primeiros segundos ou minutos, geralmente não mais de 10 minutos, após o treino, como também as substâncias que acrescentam informação (McGAUGH, 1988; IZQUIERDO, 1984, 1989). Isto sugere uma relação entre os dois fenômenos. É importante salientar que, da conjunção destes dois fenômenos no momento imediatamente posterior à aquisição de uma memória, dependerá, em boa parte, toda a vida futura dessa memória: se será firme ou fraca, se será muito ou pouco dependente para sua evocação de fatores neurohumorais, etc.
Outro dado que sugere uma inter-relação entre esses dois processos é o fato de que o isolamento cirúrgico da amígdala, por secção da stria terminalis (McGAUGH et al., 1987, p. 839-44), ou o bloqueio b-noradrenégico da amígdala (McGAUGH, 1988, p. 33-64), interferem tanto com o efeito de adrenalina e outros agentes sobre a consolidação, como com o efeito pós-treino da b-endorfina ou de seu antagonista, a naloxona, que age bloqueando os efeitos "informáticos" da b-endorfina (IZQUIERDO, 1989).
É possível que, além de acrescer informação, a b-endorfina também exerça um efeito modulador sobre a memória; e que além de bloquear o acréscimo de informação produzido pela b-endorfina, a naloxona também atue favorecendo a consolidação (McGAUGH, 1988). Dada a variedade de projeções do sistema b-endorfinérgico central: ao septum, amígdala, locus ceruleus, regiões periventriculares do hipotálamo e substância cinzenta periaquedutal (IZQUIERDO, 1989), é possível que esse peptídeo exerça efeitos diferentes segundo o lugar. BOSTOCK et al. (1988) observaram efeitos da injeção pós-treino de b-endorfina e naloxona no núcleo medial do septum, comparáveis aos obtidos pela injeção sistêmica ou intra-amigdalina dessas substâncias. É provável que a b-endorfina e a naloxona atuem modulando a consolidação em alguns lugares do cérebro, e atuem produzindo ou bloqueando, respectivamente, informação pós-aquisição, agindo sobre outros lugares. De fato, a injeção pós-treino de morfina ou de dinorfina em camundongos atua inibindo a consolidação, não através do acréscimo pós-treino de informação: contrariamente à b-endorfina, o efeito pós-treino da morfina ou da dinorfina não é reversível por uma nova injeção das mesmas substâncias antes do teste (McGAUGH, 1988; CASTELLANO, 1987).
Incorporação de Informação Adicional: informação cognitiva
Além da incorporação de informação proveniente da influência de substâncias endógenas liberadas pelas próprias experiências, estas podem incorporar, depois de terem sido adquiridas, informação proveniente de outros aprendizados (IZQUIERDO et al., 1988b, p. 105-206; IZQUIERDO e CHAVES, 1988; IZQUIERDO, 1989).
As memórias não se adquirem isoladamente, senão uma após a outra: a vida diária pode ser descrita como uma continua experiência, ou como uma continuidade de experiências consecutivas. Muitas vezes, memórias adquiridas em forma consecutiva podem se interligar de tal maneira que, depois, o indivíduo as recorda como uma experiência única (LOFTUS e YUILLE, 1984, p. 163-83). A conexão entre memórias consecutivas pode resultar numa alteração qualitativa de ambas, ou numa alteração quantitativa da primeira. Este último caso é muito mais freqüente, e pode consistir numa facilitação retrógrada, que é relativamente rara, ou numa interferência retrógrada, que é muito comum. Um exemplo típico é a apresentação de comentários derrogatórios referentes a um evento ou um conhecimento adquirido minutos ou poucas horas antes: o comentário interfere com a memória desse conhecimento ou evento (IZQUIERDO et al., 1988a,b; IZQUIERDO, 1989).
Em 1900, Müller e Pilzecker descreveram a interferência retrógrada, explicando-a, erroneamente, por uma interferência com a consolidação. Recentemente, foi determinado fora de qualquer questão que a consolidação de cada item de memória é independente, e que a consolidação, como um todo, é independente da interferência retrógrada (IZQUIERDO e PEREIRA, 1989; IZQUIERDO, 1989) ou de qualquer tipo de acréscimo post hoc de informação às experiências (IZQUIERDO, 1989).
A interferência retrógrada se observa em animais e humanos (CAHILL et al., 1986, p. 454-56; IZQUIERDO et al., 1988a,b; PEREIRA et al., 1988, p. 402-404). Não é causada por qualquer associação entre dois aprendizados consecutivos; algumas associações podem causá-la, outras não. Em animais, a segunda experiência deve reunir certas características: ser alertante, mas menos alertante que a experiência precedente a ser interferida (IZQUIERDO et al., 1989). Por ex., uma habituação a um tom depois de um aprendizado aversivo o interfere, mas o inverso não ocorre (NETTO et al., 1985, p. 515-20). A tarefa ou experiência interferente não tem por que ter relação cognitiva com a tarefa interferida (Idem; CAHILL et al., 1986, p. 454-56; PEREIRA et al., 1988, p. 402-404; IZQUIERDO, 1989), mas uma relação cognitiva inversa pode contribuir à sua ação interferente: por ex., urna experiência num ambiente muito amplo e iluminado pode interferir com um aprendizado prévio num ambiente muito restrito e escuro (IZQUIERDO et al., 1989). No homem, um comentário pejorativo, não-factual, pode interferir sensivelmente com a memória de fatos concretos aprendidos poucos minutos ou horas antes (IZQUIERDO e CHAVES, 1988).
Tanto em animais como em humanos, a interferência retrógrada é inibida pelo diazepam (CAHILL et al., 1986; PEREIRA et al., 1988; IZQUIERDO et al., 1988a,b; IZQUIERDO, 1989), que age favorecendo a neurotransmissão GABAérgica. É provável que sinapses GABAérgicas sejam responsáveis pelo fechamento do registro de memória correspondente a cada experiência, fazendo com que não se possa incorporar mais informação post hoc às mesmas.
A interferência retrógrada tem relação com os problemas de memória que se observam em sujeitos ansiosos e em pacientes dementes com agitação. A amnésia retrógrada imediata (para fatos recém-aprendidos) que se observa nesses pacientes é empíricamente combatida com o diazepam e/ou com outras benzodiazepinas. (GREEN (1964, p. 561-608) foi o primeiro em levantar a possibilidade de que a amnésia retrograda imediata, causada pela interferência retrógrada, fosse uma síndrome "postdistracional". Considerou óbvio que a expressão "perda da memória recente" é "inapropriada"; disse que "parece simplesmente bobo e confuso chamá-la perda de memória intermediária": sugeriu, por isso, o termo "amnésia postdistracional". Com isto, Green estabeleceu as bases do conceito moderno de interferência retrógrada desenvolvido em nosso laboratório (IZQUIERDO et al., 1988a,b; IZQUIERDO, 1989).
A incorporação de informação cognitiva às experiências ocorre fundamentalmente nas primeiras 3 horas após cada experiência, tanto no homem como em animais (IZQUIERDO et al., 1988a; IZQUIERDO, 1989). Este fenômeno, portanto, segue um curso de tempo diferente do tempo da consolidação ou do tempo da incorporação de informação farmacológica, que se estende, no máximo, por 10 minutos depois de cada experiência.
Formação de "Registros"
As seqüências de memórias que armazenamos como registros ou capítulos (e.g., a memória de um filme, a memória da manhã de ontem, a memória do jornal de hoje, etc.) se originam justamente pela peculiaridade que tem cada memória isolada de incorporar informação adicional. Vimos acima como esta incorporação ocorre predominantemente no periodo das poucas horas seguintes a cada evento ou experiência (IZQUIERDO, 1989).
Porém, as memórias podem ser também modificadas (os registros podem ser ampliados) posteriormente, às vezes anos depois (IZQUIERDO et al., 1988b). Uma nova experiência, ou evento, ou detalhe, acrescenta ou tira informação de uma memória velha (NEISSER, 1982). Assim, a imagem de nosso companheiro de aula, "X", a quem não víamos há 30 anos, pode ser inteiramente mudada pelo encontro desse mesmo "X" anteontem; a imagem do novo "X", cinqüentão, careca, se junta com, e modifica a imagem do "X" de antigamente.
É possível que a evocação também altere as memórias (IZQUIERDO et al., 1988b). Há muita evidência disso (NEISSER, 1982). A memória (o registro) que guardamos de determinado fato pode não ser a do fato em si, senão da terceira ou da vigésima vez que o evocamos. Quem é, para nós, George Washington? O Libertador dos Estados Unidos, como nos ensinaram no colégio, ou a evanescente figura que ilustra essas cobiçadas notas de um dólar que vimos meia hora atrás?
Em suma: os registros se formam basicamente durante e depois de cada experiência ou evento memorizado, amalgamando várias memórias consecutivas (LOFTUS e YUILLE, 1984, p. 163-83; IZQUIERDO et al., 1988b; IZQUIERDO, 1989). Porém, podem ser reacondicionados, alterados ou ampliados tempos depois, pela evocação ou por novas memórias interpoladas (IZQUIERDO et al., 1988). Não guardamos itens isolados como memórias, sensações ou percepções avulsas: guardamos e evocamos registros, memórias complexas.
Esquecimento
O esquecimento é "a outra cara" da memória (CASTELLANO, 1987), ou o aspecto mais saliente da memória: é muito mais o que esquecemos que o que recordamos.
Vários fatores causam esquecimento. Um, clássico, é a extinção, ou perda geralmente gradativa de uma memória por sua evocação reiterada sem reforço; ou seja, sem aquele(s) componente(s) que a fizeram marcante quando adquirida: um choque elétrico no caso de um aprendizado aversivo, comida no caso de um aprendizado alimentício, a resposta no caso de um número telefônico, o pagamento no caso de um serviço prestado. A simples passagem do tempo é outro fator no esquecimento. Influi mais, claramente, quanto pior tenha sido a gravação original: é mais fácil esquecer um número telefônico de um cinema, aprendido ontem, que um fato marcante da infância. A interferência retrógrada é outro fator.
Não há dúvida que algum grau de esquecimento é necessário para poder ter uma vida útil. É preciso esquecer para poder pensar; para poder fazer generalizações, sem as quais é impossível desenvolver qualquer atividade cognitiva (BORGES, 1960). É difícil conceber a criação sem esquecimento; o esquecimento diferencia a criação da clonagem.
O esquecimento é normal. O excesso de esquecimento, a perda real de memórias que não queremos perder, é patológico, denomina-se amnésia, e será analisada mais adiante.
Regulação da Evocação
Vimos acima que as memórias se evocam, via de regra, em blocos, como partes de registros ("files"). A primeira palavra de uma canção nos faz lembrar do resto da letra; as primeiras quatro notas da Terceira Sinfonia nos fazem lembrar de toda a Sinfonia e de seu autor.
Há uma exceção à regra anterior. Os sonhos consistem na evocação desorganizada de memórias em combinações muito variadas e de diferente grau de complexidade, geralmente incluindo interações entre memórias muito antigas e muito recentes, e entre ambas e o estado anímico e o emocional (e, portanto, neurohumoral e hormonal) do sujeito, antes ou durante o sono (SELIGMAN, 1987, p. 46-53). Não se conhecem os mecanismos da evocação utilizados nos sonhos; é provável que incluam fatores neurohumorais e hormonais e, ao mesmo tempo, estímulos externos eventualmente percebidos durante o sono (IZQUIERDO et al., 1964, p. 277-96; SELIGMAN, 1987).
Na vigília, a evocação não ocorre espontaneamente; é sempre desencadeada por algum estímulo ou estímulos iguais ou semelhantes àqueles inerentes a cada experiência (WEISKRANTZ, 1985, p. 380-415). Ninguém evoca corretamente movimentos natatórios sem cair na água. O exemplo típico são as dicas ("cues") utilizadas pelos atores ou professores para lembrar longas tiradas. Uma palavra evoca uma cena completa, no caso do ator, ou boa parte de uma aula, no caso do professor. Algumas memórias requerem muitos estímulos para serem lembradas: é o caso das memórias de atos-motores elaborados ou complexos, ou de procedimentos com muitas regras (IZQUIERDO, 1989). Os estímulos capazes de promover ou induzir evocação incluem substâncias endógenas, como a b-endorfina e os hormônios do stress (IZQUIERDO e McGAUGH, 1985, 1987; IZQUIERDO et al., 1988a,b).
Dados recentes (BRIONI e IZQUIERDO, 1988, p. 151-53; BRIONI et al., 1989) sugerem a participação de um mecanismo colinérgico muscarínico na regulação da evocação. Memórias enfraquecidas por um tratamento subagudo com etanol podem ser reativadas pelo agonista muscarínico, oxotremorina (BRIONI et al., 1989). O efeito não se deve à reiteração de atributos neurohumorais da experiência presentes durante sua aquisição (IZQUIERDO et al., 1989), nem por uma influência sobre processos de consolidação (BRIONI et al., 1989). O efeito da oxotremorina é antagonizado pela escopolamina (BRIONI e IZQUIERDO, 1988). É provável que o efeito benéfico dos agonistas colinérgicos às vezes observado no tratamento empírico das demências possa ser explicado em boa parte por uma influência preponderante sobre a evocação (Idem IZQUIERDO, 1989). Há dados que sugerem uma conexão, na regulação da evocação, entre o sistema b-endorfínico e o mecanismo colinérgico sensível à oxotremorina e escopolamina (BRIONI e IZQUIERDO, 1988). Um vínculo possível entre ambos os sistemas é o núcleo septal mediai, que é colinérgico, sensível à b-endorfina, e recebe projeções b-endorfínicas (IZQUIERDO et al., 1988a; IZQUIERDO, 1989).
É evidente que os sistemas moduladores, por sua ação, tanto na hora da gravação como da evocação das memórias, exercem poderosa influência para decidir sobre a intensidade que terão ambos os processos. Uma memória cuja consolidação ocorre sob a influência de um nível moderadamente elevado de adrenalina, por ex., será gravada com mais intensidade que outra adquirida sem essa influência (McGAUGH, 1988, p. 33-64). Uma memória gravada sob a influência de um nível exagerado de adrenalina será mais fraca: só poderá ser evocada se o animal libera essa substância ou recebe uma injeção da mesma na hora da evocação (IZQUIERDO, 1984, p. 65-77).
Pode-se afirmar que as memórias dependem, para sua correta evocação, do estado hormonal e neurohumoral presente depois da aquisição (que faz com que se gravem melhor ou pior), e do estado hormonal e neurohumoral presente no momento da evocação (IZQUIERDO, 1984, 1989). É evidente que uma disfunção generalizada dos sistemas moduladores neurohumorais e hormonais pode provocar um déficit da capacidade de evocação e uma conseqüente perda ou falha da memória.
Amnésias
Denomina-se amnésia a perda de memórias preexistentes (amnésia retrógrada), ou a incapacidade de formar memórias novas (amnésia anterógrada). A perda total de memórias, anterógrada e retrógrada, se denomina amnésia global, esta é, geralmente, transitória e reversível, e se observa após grandes traumatismos cranianos ou acidentes vasculares cerebrais com edema cerebral (SQUIRE, 1987).
A lesão bilateral da amígdala e do hipocampo provoca amnésia anterógrada pura, explicável pela supressão do principal núcleo modulador e do principal mecanismo seletor, respectivamente. Esta amnésia foi descrita pela primeira vez, e posteriormente estudada em detalhe, no paciente H. M., a quem foram extirpados, em 1953, bilateralmente, a amígdala, o hipocampo e o córtex temporal adjacente, para o tratamento de epilepsia temporal. H. M. lembra bem fatos e experiências anteriores à operação; mas não conseguiu, desde então, adquirir praticamente nenhuma memória nova, com exceção de algumas muito simples (MARKOWITSCH e PRITZEL, 1985, p. 189-287). (Esse tipo de cirurgia radical não se usa mais no tratamento da epilepsia temporal desde o lamentável incidente com H. M. e suas seqüelas.) Pacientes com lesões bilaterais da amígdala e do hipocampo, ou de seus sítios de projeção diencefálicos, apresentam graus variáveis de amnésia anterógrada (WEISKRANTZ, 1985, p. 380-415; SQUIRE, 1987).
A amnésia retrógrada ocorre quase que invariavelmente associada com outros múltiplos déficits cognitivos (desorientação, alterações da percepção e da inteligência, diversos graus de amnésia anterógrada, etc.) em quadros com lesões cerebrais múltiplas e difusas: as demências. Denomina-se demência justamente a perda global, geralmente irreversível, das faculdades cognitivas em humanos.
Dos casos de demência, 50% são devidos à Doença de Alzheimer, antigamente chamada demência senil ou arterosclerótica (MOOS et al., 1988, p. 353-91). Sua causa é desconhecida e se acompanha de lesões muito disseminadas, tanto em áreas corticais como em núcleos e projeções colinérgicas, noradrenérgicas, dopaminérgicas serotoninérgicas e b-endorfínicas (IZQUIERDO, 1987, 1989). A Doença de Alzheimer inicia de forma insidiosa depois dos 50 anos, mais comumente depois dos 70; em pacientes portadores da Síndrome de Down, sua incidência é muito alta e seu começo pode ser numa idade muito menor (40 anos). Demências com lesões também difusas podem se observar em pacientes com infartos cerebrais múltiplos por embolia ou hemorragia (10 a 30% das demências). As lesões vasculares podem ser secundárias à arterosclerose (metade ou mais dos casos), ou a traumatismos cranianos reiterados (por ex., nos boxeadores), ao alcoolismo crônico (10% das demencias) ou outras intoxicações (solventes orgânicos, alumínio, monóxido de carbono, etc.). Um tipo particular de demência pós-alcoólica é a Síndrome de Korsakoff, em que as lesões se circunscrevem em grande parte aos sítios de projeção hipotalâmicos da amígdala e do hipocampo, e a regiões talâmicas associativas; o quadro às vezes se acompanha também de lesões em regiões motoras cerebelares e mesencefálicas, em cujo caso denomina-se Síndrome de Wernicke-Korsakoff (WEISKRANTZ, 1985, p. 380-415; MARKOWHSCH e PRITZEL, 1985, p. 189-287). Em muitos casos (10 a 20% das demências), coexistem lesões vasculares e/ou quadros pós-alcoólicos, ou outras alterações neurológicas, como a Doença de Alzheimer. Em várias síndromes neurológicas (Parkinson, Creutzfeld-Jakob, Coréia de Huntington, hidroencefalia com pressão ventricular normal, etc.), uma síndrome demencial pode aparecer como complicação, muitas vezes terminal, da doença. As lesões, nesses casos, são também disseminadas e múltiplas. A Síndrome da Imunodeficiência Adquirida inclui, muitas vezes, um quadro demencial, que às vezes é manifestação precoce da doença, e cuja patologia não é ainda bem conhecida.
Pode-se observar amnésia retrógrada ou global leve, sem outros transtornos cognitivos, em humanos ou animais com idade muito avançada (amnésia senil benigna). A explicação habitual é a perda neuronal progressiva que se inicia, no homem, no primeiro ano de vida, mas pode atingir proporções importantes depois dos 70 ou 80 anos.
Na amnésia retrógrada humana, inclusive nas demências, o déficit se deve em boa parte a um déficit de evocação, e não de armazenamento. O déficit de evocação é corrigível em parte, nas fases iniciais: a apresentação de estímulos apropriados, cognitivamente relacionados com as memórias a serem evocadas (WEISKRANTZ, 1985), ou a simples exposição a situações ou ambientes novos, que libera a b-endorfina (IZQUIERDO et al., 1988a,b), pode melhorar a memória em pacientes com profunda amnésia retrógrada. Com o avanço da doença, isto, como é óbvio, acaba sendo impossível. É provável que as lesões cerebrais múltiplas das demências causem uma disfunção dos sistemas neurohumorais envolvidos na evocação, e que seja esta a principal causa do déficit observado (IZQUIERDO, 1987, p. 325-27).
Na depressão grave, pode ocorrer um quadro que costuma ser denominado pseudodemência, com profunda amnésia retrógrada e anterógrada e outros transtornos cognitivos. Um tratamento medicamentoso adequado (geralmente com drogas que estimulam sistemas noradrenérgicos centrais, como o Lítio e os antidepressivos tricíclicos) reverte o quadro, recuperando a memória em níveis normais. Obviamente, não houve perda de material gravado, senão um problema de evocação: recuperado o estado neurohumoral do indivíduo, desaparece o quadro depressivo, e recuperam-se memórias que julgava ter perdido. É importante salientar que a depressão acompanha, e complica, muitas doenças crônicas, incluindo as demências, e que é bastante comum na idade senil. O diagnóstico diferencial entre depressão e demência é fundamental, pois a depressão tem tratamento definido e, se tratada, bom prognóstico; e a demência, não.
Nas fases avançadas e terminais das demências, que são invariavelmente progressivas e se agravam com a idade, principalmente a Doença de Alzheimer, a amnésia retrógrada não responde mais a estímulos facilitadores da evocação (SQUIRE, 1987). Nestes casos, não há dúvida de que o déficit consiste numa perda real de memórias, por lesão generalizada de sítios de armazenagem; e não há mais como corrigi-lo (IZQUIERDO, 1987).
Não há, no momento, um tratamento apropriado para a amnésia das síndromes demenciais. A exposição dos pacientes a estímulos novos (CHAVES e IZQUIERDO, 1986, p. 211-19; IZQUIERDO, 1989), ou a fragmentos das memórias a serem evocadas (WEISKRANTZ, 1985) é um paliativo para o componente retrógrado. Se houver agitação ou ansiedade, com a resultante presença de interferência retrógrada, o diazepam ou outras benzodiazepinas podem ser outro paliativo (CAHILL et al., 1986; IZQUIERDO, 1989). A amnésia anterógrada, devida à lesão dos aparelhos seletores e consolidadores, não pode ser corrigida. Pode ser minorada, em parte, e, dependendo da gravidade do caso (da lesão), por drogas como estimulantes colinérgicos, derivados do ACTH ou da vasopressina, etc. Várias destas drogas atuam também, como vimos, favorecendo a evocação, e seu efeito moderadamente benéfico, nas síndromes demenciais, pode ser em boa parte devido a este último efeito (IZQUIERDO, 1989). Nos últimos anos, tem se usado, com sucesso exíguo e muito variável, uma família de drogas muito atóxicas chamadas nootrópicos (piracetam, vincamina e derivados). Em geral, seu efeito não é melhor que o de um placebo; mas as empresas que as produzem advogam entusiásticamente por seu emprego (MOOS et al., 1988). O futuro da farmacologia da memória, porém, não parece passar por essa família heterogênea de drogas (IZQUIERDO, 1989).
Armazenagem das Memórias
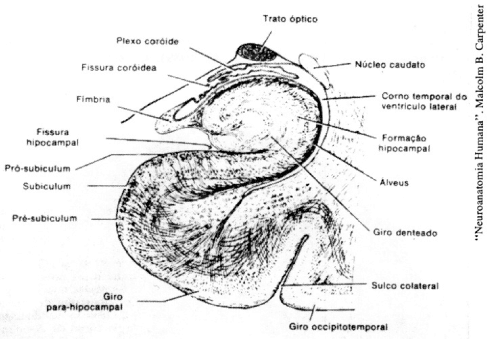
A necessidade de que existam lesões cerebrais difusas, principalmente corticais, para que se observe amnésia retrógrada (LASHLEY, 1930, p. 1-24), como nas demências (MARKOWITSCH e PRITZEL, 1985; SQUIRE, 1987) sugere que a armazenagem das diversas memórias também é sumamente disseminada, preponderantemente cortical e provavelmente muito redundante. Admite-se, de fato, que as memórias se armazenam no córtex e dependem para sua integração, no momento da evocação, de numerosas conexões aferentes e eferentes dessas regiões. Claramente, o hipocampo e a amígdala não são sítios de armazenamento; nem o paciente H. M., nem outros similares (MARKOWITSCH e PRITZEL, 1985), nem animais de experimentação com lesão de ambas as estruturas (SQUIRE, 1987; IZQUIERDO, 1989), apresentam amnésia retrógrada.

Claramente, também, não há um único sítio de armazenagem para todas as memórias nem para cada tipo possível de memória; senão, cada lesão vascular ou cada morte de um grupo neuronal causaria a perda isolada de um fragmento determinado de uma memória; o que não acontece nunca. Ninguém lembra sua primeira namorada, com exceção do olho direito, por exemplo. É, porém, possível que certas variedades ou categorias de registros se conservem em regiões separadas: há casos de amnésia para objetos e não para animais, ou vice-versa , ou de perda seletiva de alguma língua, mas não de outras, por ex. (MARSHALL, 1988, p. 378). Evidência em animais sugere que certas memórias-motoras muito simples se armazenam num circuito que inclui o cerebelo (THOMPSON, 1986, p. 941-47), que as memórias visuais se guardam no córtex visual (MISHKIN et al., 1984, p. 65-77), e que algumas discriminações complexas se armazenam no córtex pré-frontal (SQUIRE, 1987). É possível que muitas memórias se armazenem através de várias representações, em diversos lugares, e que algumas representações simples, incompletas, secundárias ou elementares se conservem em estruturas subcorticais, diencefálicas, mesencefálicas ou cerebelares. A grande maioria das memórias, sem dúvida, se armazena em circuitos múltiplos e redundantes, já que sobrevive a lesões muito amplas e difusas (LASHLEY, 1929; MARKOWITSCH e PRITZEL, 1985), e é possível registrar alterações difusas da atividade neuronal unitária relacionadas com sua aquisição ou evocação em diversas áreas cerebrais (JOHN, 1967).
Não se conhecem os mecanismos pelos quais as memórias se mantêm armazenadas durante tanto tempo, às vezes décadas. O único fenômeno eletrofisiológico conhecido que pode durar meses é a potenciação de longa duração. Consiste no aumento possivelmente permanente da eficiência de um determinado grupo de sinapses depois de sua estimulação repetida. Foi descrita originalmente no hipocampo que, como vimos, não pode ser um sítio de armazenamento de memórias , mas foi observada mais recentemente no cortex (IZQUIERDO, 1989). Pode ser considerada um modelo, ou urna forma eletrofisiológica de memória; mas não há evidência de que seja um mecanismo da memória. Outra possibilidade para explicar a persistência das memórias é a eliminação progressiva de sinapses ou neurônios desnecessários e potencialmente interferentes. De fato, ocorre uma marcada diminuição do número de neurônios em muitas regiões cerebrais no primeiro ano de vida (um período certamente de muito aprendizado), que continua em menor grau ao longo dos anos (à medida que vamos aprendendo mas também envelhecendo). Uma possibilidade sugerida no início do século por Cajal para explicar a memória é a formação de novas sinapses; a formação de novas sinapses depois de experiências complexas tem sido descrita em animais por GREENOUGH (1985, p. 77-103). Não é impossível que algumas memórias obedeçam a um desses mecanismos, outras a outro, e muitas, ou todas, a nenhum dos três. O certo é que, até agora, das dezenas de hipóteses bioquímicas e eletrofisiológicas da memória que foram propostas ao longo dos anos, em espécies que variam desde moluscos até o homem, nenhuma sobreviveu (alterações do RNA, da síntese proteica, do código genético, da permeabilidade iônica das membranas, etc.). Nenhuma dessas hipóteses conseguiu, de fato, o que é realmente essencial: a postulação de um esquema realista de tradução em nível celular de alterações cognitivas e/ou de comportamento (McGAUGH, 1988; ROSE, 1988; IZQUIERDO, 1989).
Conhecemos, pois, bastante sobre a modulação das memórias durante e depois de sua aquisição e na hora da evocação; mas não sabemos de que estão feitas as memórias, se é que estão feitas de alguma coisa.
CASTELLANO (1987) comparou as memórias com a luz das estrelas, que ainda brilham na noite quando já não existem mais. É possível que o complicado e ainda muito desconhecido hardware cerebral possa fazer, alterar e desfazer memórias sem alterar o funcionamento de nenhuma sinapse ou processo bioquímico em forma estrutural ou permanente. Já em 1964, Green afirmou que a "memória pode ser considerada em termos de modelos ou abstrações matemáticas, e é até concebível que o estado de memória em si seja algum tipo de abstração sem realidade tangível".
A hipótese, impensável até poucos anos atrás, sustenta atualmente dezenas de centros de pesquisa no mundo todo: aqueles que analisam o funcionamento cerebral desde o ponto de vista de redes neurais. Nesses centros, físicos, engenheiros, matemáticos e até alguns biólogos procuram uma lógica ou uma logística que, até agora, os biólogos sozinhos não acharam (ver, por ex., DELACOUR e LEVY, 1988).
Às vezes o espectador de um jogo de xadrez vê o xeque-mate que os próprios jogadores não enxergam. Se conhece bem as regras, é claro.
Agradecimentos
Trabalho desenvolvido com apoio da Financiadora de Estudos e Projetos (FINEP). O autor agradece a John Davis Green (1917-1964) e Jorge Luis Borges (1899-1986), os espectadores do jogo da memória que melhor entenderam suas regras, entre os muitos que conheceu.
Referências Bibliográficas
BORGES, J. L. 1960. Otras inquisiciones. Buenos Aires, Emece.
BOSTOCK, E., GALLAGHER, M. & KING, R. A. 1988. Effects of opioid microinjections into the medial septal area on spatial memory in rats. Behav. Neurosci., 102: 643-652.
BRIONI, J. D. & IZQUIERDO, I. 1988. Retention enhancement by pre-test b-endorphin and oxotremorine and its reversal by scopolamine. Behav. Neural Biol., 50: 151-153.
BRIONI, J. D., McGAUGH, J. L. & IZQUIERDO, L 1989. Amnesia induced by short-term treatment with ethanol: attenuation by pre-test oxotre-morine. Pharmacology, Biochemistry & Behavior, no prelo.
CAHILL, L., BRIONI, J. D. & IZQUIERDO, I. 1986. Retrograde memory enhancement by diazepam: its relation to anterograde amnesia and some clinical implications. Psychopharmacology, 90: 454-456.
CASTELLANO, C. 1987. La memoria. Roma, Editori Reuniti.
CHAVES, M. L. F. & IZQUIERDO, I. 1986. Previous exposure to a novel experience enhaces performance in two simple memory tests in humans. Braz. J. Med. Biol. Res., 19: 211-219.
EBBINGHAUS, H. 1885. Über das Gedächtniss. Leipzig, Drucker & Humblatt.
GOLD, P. E. & DELANOY, R. 1981. ACTH modulation of memory storage processing. In: MARTINEZ, J. L., JENSEN, R. B., MESSING, R. B., RIGTER, H. & McGAUGH, J. L., eds. Endogenous peptides and learning and memory processes. New York, Academic Press, p. 79-98.
GRAY, J. A. 1982. The neuropsychology of anxiety: an enquiry into the function of the septo-hippocampal system. Oxford, Clarendon.
GREEN, J. D. 1964. The hippocampus. Physiol. Rev., 44: 561-608.
GREENOUGH, W. T. 1985. The possible role of experience-dependent synaptogenesis, or synapses on demand, in the memory process. In: WEINBERGER, N. M., McGAUGH, J. L. & LYNCH, G. eds. Memory systems of the brain. New York, Guilford Press, p. 77-103.
HILGARD, E. R. e BOWER, G. H. 1975. Teorias del aprendizaje. México, Trillas.
HO, B., CHUTE, D. & RICHARDS, D. eds. 1978. Drug discrimination and state-dependent learning. New York, Academic Press.
IZQUIERDO, I. 1984. Endogenous state dependency: memory depends on the neurohumoral and hormonal states present after training and at the time of testing. In: LYNCH, G., McGAUGH, J. L. e WEINBERGER, N. M., eds. Neurobiology of learning and memory. New York, Guilford Press, p. 65-77.
__________. 1987. The pathogenesis of the amnesia of Alzheimer's disease. Trends Pharmacol.Sci., 8: 325-327.
__________. 1989. Different forms of posttraining memory processing. Behav. Neural Biol., no prelo.
IZQUIERDO, L, BARCIK, N. R. & BRIONI, J. D. 1989. Pre-test b-endorphin and epinephrine, but not oxotremorine, reverse retrograde interference of a conditioned emotional response in mice. Pharmacol. Biochem. Behav., no prelo.
IZQUIERDO I. & CHAVES, M. L. F. 1988. The effect of a non-factual posttraining negative comment on the recall of verbal information. Journal of Psychiatric Research, no prelo.
IZQUIERDO, I. & McGAUGH, J. L. 1985. Effect of a novel experience prior to training or testing on retention of an inhibitory avoidance task in mice: involvement of an opioid systrem. Behav. Neural Biol., 44: 228-238.
__________. 1987. Retention impairment by posttraining epinephrine: role of endogenous state dependency and of endogenous opioid mechanisms. Behav. Neurosci., 101: 778-781.
IZQUIERDO, I. & NETTO, C. A. 1985a. The role of b-endorphin in memory regulation. Ann. N. Y. Acad. Sci., 444: 162-177.
__________. 1985b. The brain b-endorphin system and behavior: the modulation of consecutively and simultaneously processed memories. Behav. Neural Biol., 44: 249-265.
IZQUIERDO, I et al. 1964. Établissement de reflexes de trace pendant le sommeil naturel chez le chat. Actual. NeurophysioL, 6:277-96.
IZQUIERDO, L, et al. 1988a. Construction and reconstruction of memories. Braz. J. Med. Biol. Res., 21: 9-25.
__________. 1988b. The organization of memories into "files". In: DELACOUR, J. & LEVY, J. C. C. eds. Systems with learning and memory abilities. Amsterdam, North Holland, p. 105-126.
IZQUIERDO, I. & PEREIRA, M.E. 1989. Posttraining memory facilitation prevents extinction but not retroactive interference. Behav. Neural Biol., no prelo.
JOHN, E. R. 1967. Mechanisms of memory. New York, Academic Press.
LASHLEY, K. C. 1930. Basic neural mechanisms in behavior. Psychol. Rev., 37: 1-24.
LOFTUS, E. F. & YUILLE, J. C. 1984. Departures from reality in human perception and memory. In: WEINGARTNER, H. & PARKER, E. S., eds. Memory consolidanon Psychobiology of cognition. Hillsdale, Lawrence Erlbaum. p. 163-183.
MARKOWITSCH, H. J. & PRITZEL, M. 1985. The neuropathology of amnesia. Progr. Neurobiol., 25: 189-287.
MARSHALL, J. C. 1988. Sensation and semantics. Nature, 334: 378.
McGAUGH, J. L. 1988. Modulation of memory storage processes. In: SOLOMON, P. R. et al. Perspectives of memory research. New York, Springer Verlag. p. 33-64.
McGAUGH, J. L. et al. 1987. Stria terminalis lesions attenuate the effects of posttraining naloxone and b-endorphin on retention. Behavioral Neuroscience, 100: 839-844.
MIKES, G. 1970. The land of the Rising Yen. London, Penguin.
MISHKIN, M. et al. 1984. Memories & habits: two neural systems. In: LYNCH, G. et al. Neurobiology of learning & memory. New York, Guilford Press, p. 65-77.
MOOS, W. H. et al. 1988. Cognition activators. Med. Res. Reviews, 8: 353-391.
MORRIS, R. G. M. et al. 1982. Place navigation impaired in rats with hippocampal lesions. Nature, 297: 681-683.
MÜLLER, G. E. & PILZECKER, A. 1900. Experimented Beitrage zur Lehre vom Gedächtniss. Zeitschr. Psychol., 1: 1-288.
NEISSER, U. 1982. Memory observed. San Francisco, Freeman.
NETTO, C. A., DIAS, R. D. & IZQUIERDO, I. 1985. Interaction between consecutive learnings: Inhibitory avoidance and habituation. Behavioral & Neural Biology, 44: 515-520.
__________. 1986. Training in an open field: Simultaneous learning of habituation and of a water finding task, and differential effect of posttraining naloxone, b-endorphin, lenenkephalin, and electroconvulsive shock. Psycho-neuroendocrinology, 11: 437-446.
O'KEEFE, J. & NADEL, L. 1978. The hippocampus as a cognitive map. Oxford, Oxford University Press.
PAVLOV, I. P. 1960. Conditioned reflexes. New York, Dover.
PEREIRA, M. L. et al. 1988. Diazepam blocks the interfering effect of posttraining behavioral manipulations on retention of a shuttle avoidance task. Psychopharmacology, 94: 402-404.
RESCORLA, R. 1985. Associative learning: some consequences of contiguity. In: WEINBERGER, N. M. et al. Memory systems of the brain. New York, Guilford. p. 211-230.
ROSE, S. P. R. 1988. Mind and memory between metaphor and molecule. In: DELACOUR, J. & LEVY, J. C. C., eds. Systems with learning & memory abilities. Amsterdam, Elsevier. p. 168-179.
SCHÜTZ, R. A. & IZQUIERDO, I. 1979. Effect of brain lesions on rat shuttle behavior in four different tests. Physiol. Behav., 23: 97-105.
SELIGM AN, M. E. P. 1987. A reinterpretation of dreams. The Sciences, 27 (5): 46-53.
SQUIRE, L.R. 1987. Memory and brain. Oxford, Oxford University Press.
THOMPSON, R. F. 1986. The neurobiology of learning and memory. Science, 233: 941-947.
WEISKRANTZ, L. 1985. On issues and theories of the human amnesic syndrome. In: WEINBERGER, N. M. et al. Memory systems of the brain. New York, Guilford. p. 380-415.
Bibliografia
AJMONE-MARSAN, C. & MATTHIES, H., eds. Neuronal plasticity and memory formation. IBRO Monograph Series, nº 9. Raven, New York, 1982.
DELACOUR, J. & LEVY, J. C. C., eds. Systems with learning & memory abilities. Amsterdam, Elsevier, 1988.
LYNCH, G. et al. Neurobiology of learning & memory. New York, Guilford Press, 1984.
McGAUGH, J. L. & HERZ, A. Memory consolidation. San Francisco, Albion, 1972.
OLTON, D. S. et al. Memory dysfunctions: an integration of animal and human research from preclinical and clinical perspectives. Annals of the New York Academy of Sciences, vol. 444, 1985.
SOLOMON, P. R. et al. Perspectives of memory research. New York, Springer Verlag, 1988.
WEINBERGER, N. M. et al. Memory systems of the brain. New York, Guilford, 1985.
WEINGARTNER, H. & PARKER, E. S., eds. Memory consolidation Psychobiology of cognition. Hillsdale, Lawrence Erlbaum, 1984.
Ivan Izquierdo é professor titular do Depto. de Bioquímica do Instituto de Biociências da UFRS e conferencista do mês de outubro, 1988, do IEA.
... os mecanismos do armazenamento das memórias são hoje tão misteriosos como quando Ebbinghaus publicou a primeira análise experimental da memória, em 1885.
Algumas memórias consistem na inibição de respostas naturais ou inatas; outras, num aumento dessas respostas ou na geração de respostas novas; outras, que não envolvem nenhuma resposta direta ou aparente.
De fato, só reconhecemos que "aprendemos algo" quando se trata de algo novo; não de algo que já sabíamos. A amígdala participa dos processos de seleção como conseqüência de sua função moduladora da consolidação.
As memórias adquiridas em estado de alerta e com certa carga emocional ou afetiva são melhor lembradas que as memórias de fatos inexpressivos ou adquiridas em estado de sonolência.
Vários sistemas hormonais também modulam a consolidação. Os hormônios mais estudados são a adrenalina, adrenocorticotrofina e vasopressina, três hormônios liberados em resposta ao alerta e/ou ao stress.
É possível que, além de acrescer informação, a b-endorfina também exerça um efeito modulador sobre a memória...
O esquecimento é "a outra cara" da memória, ou o aspecto mais saliente da memória: é muito mais o que esquecemos que o que recordamos.
É evidente que os sistemas moduladores, por sua ação, tanto na hora de gravação como da evocação das memórias, exercem poderosa influência para decidir sobre a intensidade que terão ambos os processos.
É importante salientar que a depressão acompanha, e complica, muitas doenças crônicas, incluindo as demencias, e que é bastante comum na idade senil.
Conhecemos, pois, bastante sobre a modulação das memórias durante e depois de sua aquisição e na hora da evocação; mas não sabemos de que estão feitas as memórias, se é que estão feitas de alguma coisa.
Datas de Publicação
-
Publicação nesta coleção
22 Mar 2006 -
Data do Fascículo
Ago 1989
