Resumos
Para se obter elevada produtividade de grãos na cultura de milho, é preciso a utilização de práticas de manejo que reduzam o efeito negativo da infestação de plantas daninhas sobre a cultura. As relações de competição entre a cultura de milho e as plantas daninhas são usualmente influenciadas pelo arranjo espacial das plantas da cultura e pelas características morfofisiológicas dos genótipos. Esta revisão de literatura tem o objetivo de discutir a influência do arranjo espacial e das características dos genótipos das plantas de milho nas relações de competição entre a cultura e as plantas daninhas. Em geral, arranjos mais eqüidistantes entre plantas, obtidas pela redução do espaçamento entre fileiras, promovem maior competitividade à cultura. Em adição, genótipos que apresentam alta velocidade de crescimento no início do ciclo e características de planta apropriadas, especialmente estatura, sofrem menos a competição com plantas daninhas.
Zea mays; espaçamento entre fileiras; manejo cultural de plantas daninhas
Management practices contribute to reaching high grain yield in corn, which reduce negative effects of weed infestations. The competitive relationship between corn crop and weeds are usually modified by crop plant spatial arrangement and genotype morphophysiological characteristics. This literature review aims to discuss the influence of plant spatial arrangement and corn genotype characteristics in the competition relationship with weeds. Generally, equidistant plant arrangements, achieved through row width reduction, promote greater crop competitivity. Additionally, crop genotypes which present rapid initial growth, and suitable plant characteristics, particularly plant height, can tolerate interspecific plant competition better.
Zea mays; row spacing; weed cultural management
REVISÃO BIBLIOGRÁFICA
FITOTECNIA
Manejo de plantas daninhas na cultura de milho em função do arranjo espacial de plantas e características dos genótipos
Weed management in the corn crop through plant spatial arrangement and characteristics of genotypes
Alvadi Antonio Balbinot JuniorI,1 1 Autor para correspondência. ; Nilson Gilberto FleckII
IEngenheiro Agrônomo, MSc, Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina, Estação Experimental de Canoinhas, BR 280, km 219, CP 216, 89460-000, Canoinhas, Santa Catarina. E-mail: balbinot@epagri.rct-sc.br
IIEngenheiro Agrônomo, PhD, Professor do Departamento de Plantas de Lavoura da Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul (UFRGS), CP 15100, 90001-970, Porto Alegre, RS. Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico
RESUMO
Para se obter elevada produtividade de grãos na cultura de milho, é preciso a utilização de práticas de manejo que reduzam o efeito negativo da infestação de plantas daninhas sobre a cultura. As relações de competição entre a cultura de milho e as plantas daninhas são usualmente influenciadas pelo arranjo espacial das plantas da cultura e pelas características morfofisiológicas dos genótipos. Esta revisão de literatura tem o objetivo de discutir a influência do arranjo espacial e das características dos genótipos das plantas de milho nas relações de competição entre a cultura e as plantas daninhas. Em geral, arranjos mais eqüidistantes entre plantas, obtidas pela redução do espaçamento entre fileiras, promovem maior competitividade à cultura. Em adição, genótipos que apresentam alta velocidade de crescimento no início do ciclo e características de planta apropriadas, especialmente estatura, sofrem menos a competição com plantas daninhas.
Palavras-chave:Zea mays, espaçamento entre fileiras, manejo cultural de plantas daninhas.
ABSTRACT
Management practices contribute to reaching high grain yield in corn, which reduce negative effects of weed infestations. The competitive relationship between corn crop and weeds are usually modified by crop plant spatial arrangement and genotype morphophysiological characteristics. This literature review aims to discuss the influence of plant spatial arrangement and corn genotype characteristics in the competition relationship with weeds. Generally, equidistant plant arrangements, achieved through row width reduction, promote greater crop competitivity. Additionally, crop genotypes which present rapid initial growth, and suitable plant characteristics, particularly plant height, can tolerate interspecific plant competition better.
Key words:Zea mays, row spacing, weed cultural management.
INTRODUÇÃO
A convivência de plantas daninhas com lavouras pode reduzir a produtividade das culturas, obrigando o produtor de milho a utilizar práticas de manejo que significam elevação nos custos de produção (INDICAÇÕES, 2001). Plantas de milho apresentam elevada capacidade em absorver e utilizar água e nutrientes, além de possuírem elevada capacidade de utilização de radiação solar. Todavia, a presença de plantas daninhas desde a fase inicial de desenvolvimento da cultura pode acarretar perdas, inclusive totais (RAJCAN & SWANTON, 2001). Segundo FORD & PLEASANT (1994), a redução média de produtividade de grãos em seis genótipos de milho, sob interferência com plantas daninhas, foi de 70% em relação ao tratamento sem a presença das mesmas.
No mundo, o método de controle de plantas daninhas mais amplamente utilizado na cultura do milho é o químico, possibilitando a obtenção de elevadas produtividades (RADOSEVICH et al., 1997). Contudo, esse método de controle pode apresentar alguns problemas, tais como: possibilidade de contaminação ambiental, risco de intoxicação, aparecimento de biótipos de plantas daninhas resistentes aos herbicidas e necessidade de mão-de-obra qualificada. Por isso, há necessidade de adoção de práticas de manejo complementares que reduzam a interferência das plantas daninhas e o uso de herbicidas.
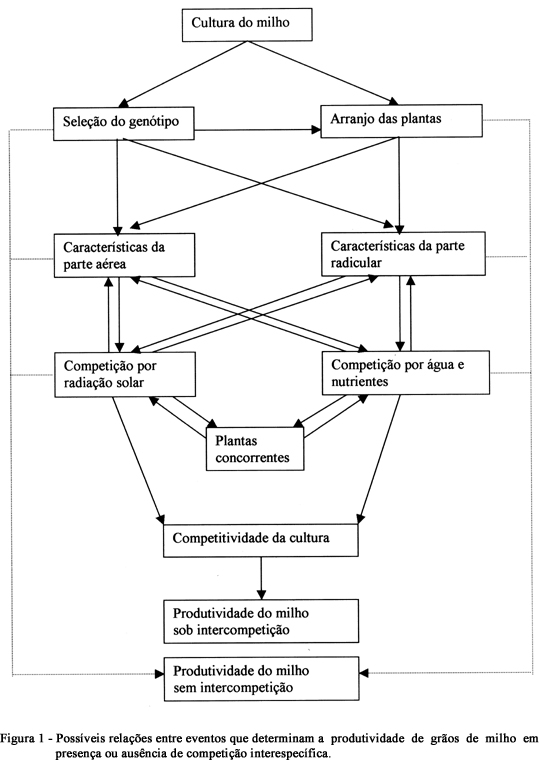
A adoção de práticas de manejo que visem posicionar a cultura em situação competitiva vantajosa em relação às plantas daninhas, constitui-se em alternativa viável para reduzir, ou até eliminar a utilização de herbicidas (TOLLENAAR et al., 1994a). É, portanto, importante no manejo integrado de plantas daninhas, o qual se baseia num conjunto de práticas que alteram as relações de competição em favor da cultura. Alterações nas relações de competição por água, luz e nutrientes entre o milho e as plantas daninhas, em favor da cultura, podem ser alcançadas por meio de práticas de manejo. Por exemplo, mudança no espaçamento entre fileiras, alterará o arranjo espacial das plantas na área. Ou ainda, a seleção de genótipos que apresentem determinadas características morfofisiológicas, como elevada estatura e rápido crescimento inicial da planta, pode elevar a competitividade com plantas daninhas. A escolha do genótipo e do arranjo espacial das plantas de milho pode influenciar vários eventos que determinam a produtividade de grãos em presença ou ausência de competição interespecífica (Figura 1).
Esta revisão discutirá a influência do arranjo espacial de plantas de milho e de características morfofisiológicas dos genótipos sobre a competitividade da cultura com plantas daninhas. Arranjo espacial das plantas de milho
A planta de milho exibe crescimento com baixa plasticidade, comparativamente às outras culturas, como arroz, trigo e soja. Isso decorre, principalmente, da ausência de afilhamento e ramificações laterais que, em geral, estão ausentes nas plantas de milho. Assim, a planta de milho apresenta reduzida variação morfológica frente às mudanças de densidade e arranjo espacial das plantas. Segundo ARGENTA et al. (2001), o arranjo espacial das plantas de milho constitui-se em prática de manejo importante para obtenção de produtividades próximas ao potencial produtivo da cultura. Modificações de ordem genética, fisiológica, bioquímica e anatômica nos genótipos de milho, realizadas pelos programas de melhoramento vegetal, fizeram com que, gradativamente, o cultivo do milho fosse realizado seguindo recomendações de alterações no arranjo espacial das plantas de milho, aumentando assim a tolerância da cultura aos estresses (SANGOI, 2001).
Espaçamento reduzido entre fileiras, em relação aos convencionalmente utilizados (0,8 a 0,9m), além de proporcionar maiores rendimentos de grãos, pode, simultaneamente, suprimir o crescimento de plantas daninhas. Mesmo que a redução do espaçamento entre fileiras, em algumas situações, não resulte em aumento do rendimento de grãos em milho, sua adoção pode justificar-se pelo aumento da competitividade com plantas daninhas, devido à maior quantidade de luz que é interceptada pelo dossel da cultura, reduzindo a dependência de herbicidas (TEASDALE, 1995). Segundo FISCHER & MILES (1973), modelos matemáticos teóricos de interferência entre plantas indicaram ser possível influenciar o resultado final da competição a favor da cultura com a utilização de arranjos espaciais quadrados ou triangulares entre plantas. Os autores estimaram que, com a redução do grau de retangularidade da distribuição entre plantas cultivadas de quatro (distância entrelinhas igual a quatro vezes a distância entre plantas na linha) para um (distância entrelinhas igual à distância entre plantas na linha), ocorreria redução de 50% na vantagem competitiva das plantas daninhas. Arranjos espaciais mais eqüidistantes promovem menor competição intraespecífica, favorecendo a cultura na competição interespecífica (NICE et al., 2001).
O arranjo espacial das plantas cultivadas constitui-se, portanto, em fator relevante na definição das relações de competição inter e intraespecífica entre plantas (FISCHER & MILES, 1973; NORRIS et al., 2001a). A redução do espaçamento entre fileiras propicia aumento da capacidade de interceptação de luz pelo dossel das plantas cultivadas. Dessa forma, a cultura ocupará o espaço de forma mais rápida, diminuindo a disponibilidade de recursos ao crescimento e desenvolvimento de plantas daninhas (THARP & KELLS, 2001). Usualmente, o termo espaço integra o conjunto de recursos necessários ao desenvolvimento das plantas (RADOSEVICH et al., 1997).
A redução do espaçamento entre fileiras do milho de 76cm para 38cm, sob mesma população de plantas, proporcionou diminuição do acúmulo de massa em Chenopodium album e Amaranthus retroflexus em até 29%, reduzindo também a quantidade de diásporos produzida pelas plantas daninhas (BEGNA et al., 2001). Observou-se, também, redução de 28% na massa acumulada em C. album, quando essa espécie competiu com milho semeado em espaçamento de 56cm em relação à semeadura com 76cm entre fileiras (THARP & KELLS, 2001). Contudo, quando o espaçamento foi reduzido de 56cm para 38cm, não houve redução de massa por unidade de área produzida por C. album. Em semeadura realizada em espaçamento de 38cm entre fileiras do milho, a cultura ocupou os espaços das entrelinhas com seu dossel uma semana antes, em relação à semeadura em espaçamento de 76cm entre fileiras (TEASDALE, 1995). Desta maneira, em espaçamentos menores, evidencia-se que a cultura de milho consegue ocupar mais rapidamente o espaço aéreo com seu dossel, reduzindo a disponibilidade de luz às plantas daninhas.
A transmitância de luz pelo dossel pode ser representada através do coeficiente de extinção luminoso (K). Quanto maior for o coeficiente K, maior será a capacidade de interceptação de luz pelo dossel analisado (FLÉNET et al., 1996). Esses autores verificaram aumento em K, durante todo ciclo da cultura, quando promoveram redução do espaçamento entre fileiras do milho, comprovando que a semeadura em arranjos espaciais mais eqüidistantes entre plantas favorece a interceptação de luz pela cultura, tornando esse recurso menos disponível às plantas daninhas. Sob reduzida quantidade de luz, a taxa fotossintética das plantas daninhas diminui, o que provoca menor potencial de dano dessas plantas às culturas (NORRIS et al., 2001b). Além disso, sob sombreamento, as plantas daninhas produzem menor quantidade de diásporos, reduzindo o abastecimento do banco de sementes no solo (RADOSEVICH et al., 1997).
Enfatiza-se, por outro lado, que alguns estudos não indicaram vantagem em alterar o arranjo espacial das plantas na competitividade da cultura. Na cultura de milho, por exemplo, a redução do espaçamento entre fileiras não influenciou a produtividade de grãos, acúmulo de massa pelas plantas daninhas e produção de diásporos por Abutilon theophrasti (TEASDALE, 1998). A variação na eqüidistância entre plantas de milho não afetou o crescimento de plantas daninhas (FERNANDEZ et al., 2002). Nesse caso, as plantas daninhas responderam ao elevado grau de sombreamento imposto pelo milho, em espaçamento reduzido, alocando maior quantidade de reservas para crescimento da parte aérea, compensando deste modo os efeitos da redução de luz. Ainda em milho, maior eqüidistância entre plantas não interferiu no crescimento e produção de diásporos de Sicyos angulatus (ESBENSHADE et al., 2001).
É provável que os efeitos decorrentes de mudanças no arranjo espacial das plantas de milho sobre a competitividade com plantas daninhas, sejam dependentes de diversos fatores, como características morfofisiológicas dos genótipos, espécies daninhas presentes na área, grau de infestação e condições de ambiente, principalmente em termos de fertilidade do solo, disponibilidade de água, radiação solar e temperatura do ar. Semeaduras de milho realizadas sob condições de baixas temperaturas do ar e do solo e reduzida intensidade luminosa, provocam baixa velocidade de crescimento das plantas no início do ciclo de desenvolvimento. Especialmente, nessa situação, a distância entre fileiras exerce importância na velocidade de ocupação do espaço pelas plantas de milho (BULLOCK et al., 1988) e, conseqüentemente, sobre a competitividade da cultura.
O arranjo espacial mais eqüidistante das plantas também pode facilitar o controle químico das plantas daninhas. Pesquisas têm demonstrado que a adoção de práticas de manejo que aumentem a competitividade da cultura, associadas com aplicações de herbicidas, pode resultar em melhor controle de plantas daninhas (SWANTON & WEISE, 1991). De outro modo, quando as fileiras estão muito afastadas entre si, a maior proximidade de plantas da cultura nas filas pode prejudicar a interceptação do herbicida pelas plantas daninhas que ocorrem junto às fileiras, reduzindo o controle. A utilização de práticas de manejo que propiciem vantagem competitiva à cultura, como arranjo eqüidistante das plantas, permite reduções na dose e, em algumas situações, no número de aplicações de herbicidas, reduzindo o custo de produção da lavoura e a contaminação do ambiente (BUHLER et al., 1992).
Competitividade de genótipos de milho com plantas daninhas
Nas últimas décadas, pesquisas com melhoramento vegetal visaram, predominantemente, a melhoria da produtividade e das características de qualidade dos grãos. Mas, nos últimos anos, pesquisas relacionadas à habilidade competitiva de genótipos de culturas com plantas daninhas ganharam importância, principalmente porque a adoção de genótipos competitivos constitui-se em prática cultural que promove reduções de custo e impacto ambiental. Neste aspecto, a utilização de genótipos de milho de elevada capacidade competitiva representa ferramenta importante para o manejo cultural de plantas daninhas (RAJCAN & SWANTON, 2001).
A habilidade competitiva de culturas pode ser expressa de duas maneiras (GOLDBERG, 1990; BUSSAN et al., 1997; JANNINK et al., 2001). A primeira se refere à habilidade da cultura em suprimir o crescimento e a produção de diásporos das plantas daninhas. A segunda se relaciona à habilidade da cultura em tolerar a presença de plantas daninhas, mantendo inalterada a produtividade. Para JORDAN (1993), a habilidade supressora deve ter preferência em programas de manejo de plantas daninhas, pois reduz a produção de diásporos de plantas daninhas que germinarão em cultivos subseqüentes.
Com a evolução do melhoramento genético em milho, através da obtenção de híbridos a partir da década de 1930, ocorreram mudanças acentuadas nas características das plantas desse cereal. Em geral, houve redução da estatura das plantas, diminuição do tamanho de folhas, aumento do ângulo foliar em relação ao solo, redução da área foliar por planta e aumento da área foliar acima da espiga (SLAFER, 1994). Essas mudanças morfológicas reduziram a capacidade competitiva de cada planta individualmente, permitindo semeaduras em maiores densidades, sem que a competição intraespecífica limitasse a produtividade de grãos. Por outro lado, as mudanças morfológicas favorecem a penetração de luz no dossel, o que, potencialmente, poderia diminuir a capacidade competitiva dos genótipos de milho modernos em competir por luz com plantas daninhas (TOLLENAAR et al., 1997). Sabe-se que o grau de sombreamento imposto pela cultura causa alterações em densidade, crescimento, morfologia e produção de diásporos por plantas daninhas (TOLLENAAR et al., 1994b).
Genótipos de milho diferem em estatura de planta, índice de área foliar, ângulo das folhas e taxa de crescimento (LINDQUIST et al., 1998). Desse modo, híbridos de milho que apresentam diferentes características morfofisiológicas demonstram variação em habilidade competitiva com plantas daninhas (BEGNA et al., 2001). Híbridos precoces, possuidores de folhas horizontais e reduzida estatura de planta permitem maior crescimento de plantas daninhas em relação aos híbridos de maior estatura com folhas eretas (MOOMAW & MARTIN, 1984), indicando efeito da estatura de planta sobre a competitividade da cultura. Entretanto, segundo ROGGENKAMP et al. (2000), genótipos de milho não apresentam efeito direto sobre a densidade das plantas daninhas, porém, interferem significativamente no crescimento e produção de diásporos por essas plantas.
Sob competição com plantas daninhas, híbridos de milho modernos apresentam menores reduções em acúmulo de massa em relação aos híbridos antigos (TOLLENAAR et al., 1997). Além disso, aqueles genótipos apresentam maior eficiência na utilização de nitrogênio e luz; por isso, em geral, são mais competitivos que os genótipos antigos (TOLLENAAR et al., 1994b). Adicionalmente, deve-se considerar o maior grau de heterose observado em híbridos modernos em relação aos híbridos antigos, pois o primeiro grupo é representado, principalmente, por híbridos dos tipos simples e triplo, enquanto o segundo grupo é representado, em geral, por híbridos duplos. Por exemplo, o híbrido moderno Pioneer 3902 apresenta maior taxa de crescimento, em comparação ao híbrido antigo Pride 5 (TOLLENAAR & AGUILERA, 1992). Segundo esses autores, isso decorre, principalmente, da maior eficiência no uso da radiação solar pelo híbrido Pioneer 3902.
Entretanto, híbridos antigos apresentaram elevada velocidade de incremento do índice de área foliar até a fase de enchimento de grãos (LINDQUIST & MORTENSEN, 1998). Assim, esses genótipos foram mais hábeis em suprimir o crescimento de Abutilon theophrasti, apresentando menor redução de produtividade de grãos sob condição de interferência, comparativamente aos híbridos modernos. É provável que isso tenha decorrido do fato dos dois grupos de híbridos terem sido semeados na mesma densidade, 42 mil plantas ha-1. Talvez essa densidade estivesse abaixo daquela considerada ideal para que os híbridos modernos expressassem plenamente seu potencial produtivo e competitivo. Nesse sentido, estudos que visem comparar a habilidade competitiva de genótipos de milho devem ser conduzidos em densidades de plantas recomendadas para cada tipo de material.
Grande parte das pesquisas já desenvolvidas para investigar a competitividade de genótipos de culturas foi realizada em arroz, trigo e soja. O incremento da capacidade competitiva de plantas tem sido atribuído a diversos fatores, como emergência precoce (ROMAN et al., 1999), elevado vigor de plântulas (HORAK & LOUGHIN, 2000; BALBINOT JR. et al., 2001; FLECK et al., 2003), rapidez de expansão foliar (BENNETT & SHAW, 2000), formação de dossel denso (BALBINOT JR. et al., 2003), elevada estatura de planta (DIARRA et al., 1985), ciclo de desenvolvimento longo (SMITH JR., 1974) e rápido crescimento do sistema radicular (SEIBERT & PEARCE, 1993).
Pesquisas associadas à determinação das variações em arquitetura de plantas de milho em competição com plantas daninhas mereceram pouca atenção, apesar da indicação de que a habilidade das plantas em alterar sua arquitetura seja importante na determinação das relações de competição por luz (REGNIER & HARRISON, 1993). Em milho, uma característica importante na definição do potencial da cultura em competir por luz é a capacidade de interceptação de luz pelas folhas localizadas acima da espiga, levando a cultura a sombrear plantas concorrentes que se posicionam abaixo desse nível (LOOMIS et al., 1968). A estatura das plantas de milho, de outra forma, demonstra ser uma característica fundamental em definir a competitividade dos genótipos (ROGGENKAMP et al., 2000).
Segundo esses autores, a produção de diásporos por Abutilon theophrasti e Setaria viridis foi cerca de 23% inferior quando essas espécies competiram com híbridos de porte elevado, comparativamente aos genótipos com menor estatura de planta.
Enfatiza-se que o aumento em habilidade competitiva do milho com plantas daninhas não deve ser vinculado à redução do potencial de rendimento de grãos.
Entretanto, alguns trabalhos demonstram existir correlação negativa entre capacidade produtiva e características morfológicas que conferem elevada supressão de plantas daninhas (SIDDIQUE et al., 1989). Isso ocorre porque, provavelmente, mudanças adaptativas podem ser alcançadas às expensas de outros processos ou características (JANNINK et al., 2001). Ou seja, ocorrência de aclimatação e/ou adaptação para elevar a capacidade competitiva pode estar relacionada à redução da capacidade de produção de órgãos reprodutivos, inclusive grãos. Incrementos em estatura de planta, massa vegetal e área foliar podem propiciar aumentos em taxa respiratória, autosombreamento e alocação de fotoassimilados em órgãos vegetativos, reduzindo o índice de colheita e a produtividade de grãos (FISCHER et al., 1997).
Por outro lado, outros estudos têm apontado que é possível aliar adequada produtividade de grãos com elevada habilidade competitiva (FISCHER et al., 1997; JANNINK et al., 2001). Em pesquisa realizada por BUSSAN et al. (1997), em que foram utilizados 16 genótipos de soja, constatou-se que a elevada competitividade da cultura não necessariamente foi alcançada às expensas de redução do potencial de rendimento da cultura. Assim, realizando trabalhos de modo integrado por cientistas das plantas daninhas e melhoristas de plantas, será possível obterem-se genótipos que aliem elevado potencial competitivo com manutenção da produtividade de grãos.
Perspectivas futuras
Há tendência em se intensificarem as pesquisas relacionadas à identificação de práticas culturais que coloquem a cultura do milho em situação competitiva vantajosa, sem diminuir o seu potencial de rendimento. Esse fato decorre da elevada importância das plantas daninhas em cultivos de milho e à necessidade de redução da aplicação de herbicidas em sistemas agrícolas que visam racionalizar o uso de agroquímicos. A adoção dessas práticas culturais pode minimizar o efeito negativo provocado por plantas daninhas, diminuindo a dependência de herbicidas e os custos da lavoura, preservando elevados patamares de produtividade de grãos.
Nesse contexto, a escolha de genótipo competitivo de milho aliada ao uso de arranjo espacial eqüidistante de plantas, constituem-se em alternativas essenciais para aumentar a competitividade da cultura com plantas daninhas. Cabe salientar que a utilização de espaçamentos entre fileiras abaixo de 80cm depende da disponibilidade de equipamentos que apresentem custo compensatório e adequada eficiência tanto na semeadura como na colheita. Por outro lado, é fundamental que a escolha de um genótipo competitivo não seja acompanhada de perda acentuada de potencial de rendimento de grãos.
CONCLUSÕES
A adoção de espaçamentos mais reduzidos entre fileiras na cultura de milho, com maior grau de eqüidistância entre plantas, aliada à utilização de genótipos mais competitivos, pode reduzir a interferência de plantas daninhas e facilitar o manejo das mesmas e, assim, diminuir a utilização de herbicidas, preservando o potencial de produtividade de grãos. A adoção destas práticas pode tornar o manejo de plantas daninhas na cultura do milho mais econômico, racional e ambientalmente correto.
Recebido para publicação 15.10.03
Aprovado em 14.07.04
- ARGENTA, G. et al. Arranjo de plantas em milho: análise do estado-da-arte. Ciência Rural, Santa Maria, v.31, n.6, p.1075-1084, 2001.
- BALBINOT JR. A.A. et al. Competitividade de cultivares de arroz irrigado com cultivar simuladora de arroz-vermelho. Pesquisa Agropecuária Brasileira, Brasília, v.38, n.1, p.53-59, 2003.
- BALBINOT JR. A.A. et al. Velocidade de emergência e crescimento inicial de cultivares de arroz irrigado influenciando a competitividade com as plantas daninhas. Planta Daninha, Viçosa, v.19, n.3, p.305-316, 2001.
- BEGNA, S.H. et al. Morphology and yield response to weed pressure by corn hybrids differing in canopy architecture. European Journal of Agronomy, Amsterdam, v.14, n.4, p.293-302, 2001.
- BENNETT, A.C.; SHAW, D.R. Effect of Glycine max cultivar and weed control on weed seed characteristics. Weed Science, Lawrence, v.48, n.4, p.431-435, 2000.
- BUHLER, D.D. et al. Integrated weed management techniques to reduce herbicide inputs. Agronomy Journal, Madison, v.84, n.6, p.973-978, 1992.
- BULLOCK, D.G. et al. A growth analysis comparison of corn grown in conventional and equidistant plant spacing. Crop Science, Madison, v.28, n.2, p.254-258, 1988.
- BUSSAN, A.J. et al. Field evaluation of soybean (Glycine max) genotypes for weed competitiveness. Weed Science, Lawrence, v.45, n.1, p.31-37, 1997.
- DIARRA, A. et al. Growth and morphological characteristics of red rice (Oryza sativa) biotypes. Weed Science, Champaign, v.33, n.3, p.310-314, 1985.
- ESBENSHADE, W.R. et al. Effect of row spacing and herbicides on burcucumber (Sicyos angulatus) control in herbicide-resistant corn (Zea mays). Weed Technology, Lawrence, v.15, n.2, p.348-354, 2001.
- FERNANDEZ, O.N. et al. Competition between corn (Zea mays) and bermudagrass (Cynodon dactylon) in relation to the crop plant arrangement. Agronomie, Les Ulis Cedexa, v.22, n.3, p.293-305, 2002.
- FISCHER, R.A.; MILES, R.E. The role of spatial pattern in the competition between crop plants and weeds. A theoretical analysis. Mathematical Biosciences, New York, v.18, p.335-350, 1973.
- FISCHER, A. et al. Suppression of junglerice [Echinochloa colona (L.) Link] by irrigated rice cultivars in Latin America. Agronomy Journal, Madison, v.89, n.3, p.516-521, 1997.
- FLECK, N.G. et al. Características de plantas de cultivares de arroz irrigado relacionadas à habilidade competitiva com plantas concorrentes. Planta Daninha, Viçosa, v.21, n.1, p.97-104, 2003.
- FLÉNET, F. et al. Row spacing effect on light extinction coefficients of corn, sorghum, soybean, and sunflower. Agronomy Journal, Madison, v.88, n.2, p.185-190, 1996.
- FORD, G.T.; PLEASANT, J.M. Competitive abilities of six corn (Zea mays) hybrids with four weed control practices. Weed Technology, Champaign, v.8, n.1, p.124-128, 1994.
- GOLDBERG, D.E. Components of resource competition in plant communities. In: GRACE, J.B.; TILMAN, D., (eds). Perspectives on plant competition London : Academic, 1990. p.27-49.
- HORAK, M.J.; LOUGHIN, T.M. Growth analysis of four Amaranthus species. Weed Science, Lawrence, v.48, n.3, p.347-355, 2000.
- INDICAÇÕES técnicas para a cultura do milho no Rio Grande do Sul Porto Alegre : FEPAGRO; EMBRAPA TRIGO; EMATER/RS; FECOAGRO/RS, 2001. n.7, 135p.
- JANNINK, J. et al. Feasibility of selection for high weed suppressive ability in soybean: absence of tradeoffs between rapid initial growth and sustained later growth. Euphytica, Dordrecht, v.120, n.2, p.291-300, 2001.
- JORDAN, N. Prospects for weed control through crop interference. Ecological Applications, Washington, v.3, n.1, p.84-91, 1993.
- LINDQUIST, J.L.; MORTENSEN, D.A. Tolerance and velvetleaf (Abutilon theophrasti) suppressive ability of two old and two modern corn (Zea mays) hybrids. Weed Science, Lawrence, v.46, n.5, p.569-574, 1998.
- LINDQUIST, J.L. et al. Mechanisms of corn tolerance and velvetleaf suppressive ability. Agronomy Journal, Madison, v.90, n.6, p.787-792, 1998.
- LOOMIS, R.S. et al. Quantitative descriptions of foliage display and light absorption in field communities of corn plants. Crop Science, Madison, v.8, n.1, p.352-356, 1968.
- MOOMAW, R.S.; MARTIN, A.R. Cultural practices affecting season-long weed control in irrigated corn (Zea mays). Weed Science, Champaign, v.32, n.4, p.460-467, 1984.
- NICE, G.R.W. et al. Sicklepod (Senna obtusifolia) response to shading, soybean (Glycine max) row spacing, and population in three management systems. Weed Technology, Lawrence, v.15, n.1, p.155-162, 2001.
- NORRIS, R.F. et al. Spatial arrangement, density, and competition between barnyardgrass and tomato: I. Crop growth and yield. Weed Science, Lawrence, v.49, n.1, p.61-68, 2001a.
- NORRIS, R.F. et al. Spatial arrangement, density, and competition between barnyardgrass and tomato: II. Barnyardgrass growth and seed production. Weed Science, Lawrence, v.49, n.1, p.69-76, 2001b.
- RADOSEVICH, S. et al. Weed ecology: implications for management 2.ed. New York : Wiley, 1997. 588p.
- RAJCAN, I.; SWANTON, C.J. Understanding maize-weed competition: resource competition, light quality and the whole plant. Field Crops Research, Amsterdam, v.71, n.2, p.139-150, 2001.
- REGNIER, E.E.; HARRISON, S.K. Compensatory responses of common cocklebur (Xanthium strumarium) and velvetleaf (Abutilon theophrasti) to partial shading. Weed Science, Champaign, v.41, n.4, p.541-547, 1993.
- ROGGENKAMP, G.J. et al. Velvetleaf (Abutilon theophrasti) and green foxtail (Setaria viridis) response to corn (Zea mays) hybrids. Weed Technology, Lawrence, v.14, n.2, p.304-311, 2000.
- ROMAN, E.S. et al. Modeling germination and seedling elongation of common lambsquarters (Chenopodium album). Weed Science, Lawrence, v.47, n.1, p.149-155, 1999.
- SANGOI, L. Understanding plant density effects on maize growth and development: an important issue to maximize grain yield. Ciência Rural, Santa Maria, v.31, n.1, p.159-168, 2001.
- SEIBERT, A.C.; PEARCE, R.B. Growth analysis of weed and crop species with reference to seed weight. Weed Science, Champaign, v.41, n.1, p.52-56, 1993.
- SIDDIQUE, K.H.M. et al. Growth, development, and light interception of old and modern wheat cultivars in a mediterranean - type environment. Australian Journal of Agricultural Research, Collingwood, v.40, n.3, p.473-487, 1989.
- SLAFER, G.A. Genetic improvement of field crops New York : Marcel Dekker, 1994. 470p.
- SMITH JR., R.J. Competition of barnyardgrass with rice cultivars. Weed Science, Champaign, v.22, n.5, p.423-426, 1974.
- SWANTON, C.J.; WEISE, S.F. Integrated weed management: the rational and approach. Weed Technology, Champaign, v.5, n.3, p.657-663, 1991.
- TEASDALE, J.R. Influence of narrow row/high corn population (Zea mays) on weed control and light transmittance. Weed Technology, Champaign, v.9, n.1, p.113-118, 1995.
- TEASDALE, J.R. Influence of corn (Zea mays) population and row spacing on corn and velvetleaf (Abutilon theophrasti) yield. Weed Science, Lawrence, v.46, n.4, p.447-453, 1998.
- THARP, B.E.; KELLS, J.J. Effect of glufosinate-resistant corn (Zea mays) population and row spacing on light interception, corn yield, and common lambsquarters (Chenopodium album) growth. Weed Technology, Lawrence, v.15, n.3, p.413-418, 2001.
- TOLLENAAR, M.; AGUILERA, A. Radiation use efficiency of an old and a new maize hybrid. Agronomy Journal, Madison, v.84, n.3, p.536-541, 1992.
- TOLLENAAR, M. et al. Effect of crop density on weed interference in maize. Agronomy Journal, Madison, v.86, n.4, p.591-595, 1994a.
- TOLLENAAR, M. et al. Effect of weed interference and soil nitrogen on four maize hybrids. Agronomy Journal, Madison, v.86, n.4, p.596-601, 1994b.
- TOLLENAAR, M. et al. Grain yield is reduced more by weed interference in an old than in a new maize hybrid. Agronomy Journal, Madison, v.89, n.2, p.239-246, 1997.
Datas de Publicação
-
Publicação nesta coleção
13 Jan 2005 -
Data do Fascículo
Fev 2005
Histórico
-
Aceito
14 Jul 2004 -
Recebido
15 Out 2003