Resumos
Para a elaboração adequada de programas de biocontrole de patógenos de solo, é necessário conhecer a distribuição de patógenos e antagonistas nesse ambiente. O objetivo deste trabalho foi estudar a presença dos gêneros Trichoderma e Fusarium em solo rizosférico e não-rizosférico, cultivado com tomateiro e pepineiro, em horta e estufa, e identificar os isolados de Fusarium spp. patogênicos às culturas. Para isso, em horta e estufa, foram realizadas 40 amostragens de solo rizosférico (tomateiro e pepineiro) e 20 de solo não-rizosférico. As suspensões dos solos foram diluídas, incubadas em meio BDA e os fungos identificados. Posteriormente, foi realizado o teste de patogenicidade para o tomateiro e o pepineiro, com os isolados de Fusarium spp. obtidos dos solos coletados. Em estufa, para o tomateiro e para o pepineiro, o número de pontos de amostragem com a presença de Trichoderma spp. em solo rizosférico foi significativamente maior (95 e 45%, respectivamente) do que em solo não-rizosférico (10%). Neste ambiente, ocorreu diferença na presença de Trichoderma spp. e Fusarium spp., sendo encontrados, respectivamente, em 10 e 55% dos pontos de amostragem. No teste de patogenicidade, cinco isolados de Fusarium oxysporum do tomateiro e seis do pepineiro foram patogênicos às respectivas culturas. Em estufa, Trichoderma spp. ocorre com maior freqüência na rizosfera, enquanto Fusarium spp. está distribuído no solo, e a maioria dos isolados de Fusarium spp. não é patogênica ao tomateiro nem ao pepineiro.
olerícolas; rizosfera; Cucumis sativus; Lycopersicon esculentum
For the adequate elaboration of biocontrol programs of soil borne pathogens, it is necessary to understand the distribution of pathogens and antagonists in this environment. This research was aimed at studing the presence of the fungi Trichoderma and Fusarium in rhizosphere and bulk soil, cultivated with tomato and cucumber, in vegetable garden and greenhouse, and to quantify the isolates of Fusarium spp. pathogenic to the crops. For that, in a vegetable garden and a greenhouse, 40 samples of rhizosphere (cucumber and tomato) and 20 of bulk soil were taken,. The soil suspensions were diluted and plated in PDA media and the fungi identified. After that, the test of pathogenicity of the isolates for tomato and cucumber found in the samples, was performed. For tomato and cucumber, in greenhouse, the number of sampling points with the presence of Trichoderma spp. in the rhizosphere (95 e 45%, respectively) was significantly greater than in the bulk soil (10%). In the bulk soil, the presence of Trichoderma spp. and Fusarium spp. differed significantly, being found, respectively, in 10 and 55% of the sampling points. In the pathogenicity test, 5 isolates of Fusarium oxysporum of tomato and 6 of cucumber were pathogenic to the respective crops. In greenhouse, Trichoderma spp. occurs more frequently in the rhizosphere, whereas Fusarium spp. is distributed in the soil and the majority of the Fusarium spp. isolates are neither pathogenic to tomato nor to cucumber.
vegetable garden; rhizosphere; Cucumis sativus; Lycopersicon esculentum
ARTIGOS CIENTÍFICOS
DEFESA FITOSSANITÁRIA
Presença dos gêneros Trichoderma e Fusarium em solo rizosférico e não rizosférico cultivado com tomateiro e pepineiro, em horta e estufa
Presence of the genus Trichoderma and Fusarium in rhizosphere and bulk soil cultivated with tomato and cucumber, in vegetable garden and greenhouse
Luciana Zago EthurI; Elena BlumeI,1 1 Autor para correspondência. ; Marlove Fátima Brião MunizI; Zaida Inês AntoniolliII; Cícero NicoliniI; Paola MilanesiI; Fabiano de Oliveira FortesIII
IDepartamento de Defesa Fitossanitária, Universidade Federal de Santa Maria (UFSM), 97105-900, Santa Maria, RS, Brasil. E-mail: eblume@smail.ufsm.br
IIDepartamento de Solos, UFSM, Santa Maria, RS, Brasil
IIIDepartamento de Fitotecnia, UFSM, Santa Maria, RS, Brasil
RESUMO
Para a elaboração adequada de programas de biocontrole de patógenos de solo, é necessário conhecer a distribuição de patógenos e antagonistas nesse ambiente. O objetivo deste trabalho foi estudar a presença dos gêneros Trichoderma e Fusarium em solo rizosférico e não-rizosférico, cultivado com tomateiro e pepineiro, em horta e estufa, e identificar os isolados de Fusarium spp. patogênicos às culturas. Para isso, em horta e estufa, foram realizadas 40 amostragens de solo rizosférico (tomateiro e pepineiro) e 20 de solo não-rizosférico. As suspensões dos solos foram diluídas, incubadas em meio BDA e os fungos identificados. Posteriormente, foi realizado o teste de patogenicidade para o tomateiro e o pepineiro, com os isolados de Fusarium spp. obtidos dos solos coletados. Em estufa, para o tomateiro e para o pepineiro, o número de pontos de amostragem com a presença de Trichoderma spp. em solo rizosférico foi significativamente maior (95 e 45%, respectivamente) do que em solo não-rizosférico (10%). Neste ambiente, ocorreu diferença na presença de Trichoderma spp. e Fusarium spp., sendo encontrados, respectivamente, em 10 e 55% dos pontos de amostragem. No teste de patogenicidade, cinco isolados de Fusarium oxysporum do tomateiro e seis do pepineiro foram patogênicos às respectivas culturas. Em estufa, Trichoderma spp. ocorre com maior freqüência na rizosfera, enquanto Fusarium spp. está distribuído no solo, e a maioria dos isolados de Fusarium spp. não é patogênica ao tomateiro nem ao pepineiro.
Palavras-chave: olerícolas, rizosfera, Cucumis sativus, Lycopersicon esculentum.
ABSTRACT
For the adequate elaboration of biocontrol programs of soil borne pathogens, it is necessary to understand the distribution of pathogens and antagonists in this environment. This research was aimed at studing the presence of the fungi Trichoderma and Fusarium in rhizosphere and bulk soil, cultivated with tomato and cucumber, in vegetable garden and greenhouse, and to quantify the isolates of Fusarium spp. pathogenic to the crops. For that, in a vegetable garden and a greenhouse, 40 samples of rhizosphere (cucumber and tomato) and 20 of bulk soil were taken,. The soil suspensions were diluted and plated in PDA media and the fungi identified. After that, the test of pathogenicity of the isolates for tomato and cucumber found in the samples, was performed. For tomato and cucumber, in greenhouse, the number of sampling points with the presence of Trichoderma spp. in the rhizosphere (95 e 45%, respectively) was significantly greater than in the bulk soil (10%). In the bulk soil, the presence of Trichoderma spp. and Fusarium spp. differed significantly, being found, respectively, in 10 and 55% of the sampling points. In the pathogenicity test, 5 isolates of Fusarium oxysporum of tomato and 6 of cucumber were pathogenic to the respective crops. In greenhouse, Trichoderma spp. occurs more frequently in the rhizosphere, whereas Fusarium spp. is distributed in the soil and the majority of the Fusarium spp. isolates are neither pathogenic to tomato nor to cucumber.
Key words: vegetable garden, rhizosphere, Cucumis sativus, Lycopersicon esculentum.
INTRODUÇÃO
O tomateiro (Lycopersicon esculentum Mill.) e o pepineiro (Cucumis sativus L.) fazem parte das principais olerícolas cultivadas em estufa e horta no Brasil. Um dos agravantes no cultivo dessas olerícolas são as doenças causadas por Fusarium spp., como as murchas e as podridões de raiz.
Fusarium é um dos gêneros mais estudados devido aos sérios problemas econômicos ocasionados pela sua ação como fitopatógeno e ampla distribuição geográfica. Os padrões de distribuição e densidade populacional podem estar relacionados ao clima, à temperatura, ao tipo de vegetação, à microbiota, aos nutrientes e ao tipo de solo (SAREMI et al., 1999). Portanto, a distribuição de patógenos de solo é de extrema importância para a recomendação de medidas de controle adequadas ao tipo de área de cultivo.
Os estudos referentes à distribuição populacional de Fusarium spp. limitam-se a áreas de grandes culturas, como soja e trigo, possivelmente por serem o principal alvo do mercado de agroquímicos. Fusarium solani (Mart.) Sacc. f. sp. glycines apresentou aumento populacional, em solo não rizosférico, pela sucessão soja aveia soja (FREITAS, 2003) e, segundo RUPE et al. (1999), a maior concentração de propágulos de Fusarium spp. foi encontrada em até 15cm de profundidade em solo infestado naturalmente. Para o trigo, foi observada variação sazonal na população de Fusarium sp. em solo não rizosférico (BATEMAN & MURRAY, 2001), o que sugere uma profundidade e época preferenciais para seu controle. Para cultivos de olerícolas, em hortas ou estufas, não são encontrados relatos sobre a distribuição desse patógeno.
Trichoderma spp. tem sido utilizado como agente de biocontrole contra uma gama de patógenos causadores de doenças em várias culturas (SIVAN & CHET, 1989). Trichoderma spp. nativo foi encontrado na rizosfera de algumas culturas, como nas de citros (GESHEVA, 2002), batata, algodoeiro (SIVAN & CHET, 1989) e tomateiro (LARKIN & FRAVEL, 1998), uma vez que o isolamento de possíveis antagonistas geralmente ocorre em solo rizosférico devido à habilidade dos isolados em habitar esse ambiente (COOK, 1993). Embora existam relatos da existência de Trichoderma spp. em solo rizosférico e não-rizosférico, não se encontram dados sobre sua distribuição em áreas de cultivo de olerícolas, principalmente em ambiente protegido, o qual apresenta modificações ambientais peculiares.
O entendimento da presença e distribuição de Trichoderma spp. e de Fusarium spp. no solo de diferentes culturas, em diferentes ambientes de cultivo, é importante para a elaboração de programas eficazes no biocontrole de patógenos de solo. Nesse contexto, o objetivo deste trabalho foi estudar a presença dos gêneros Trichoderma e Fusarium em solo rizosférico e não-rizosférico, cultivado com tomateiro e pepineiro, em horta e em estufa, e quantificar os isolados de Fusarium spp. patogênicos a cada cultura.
MATERIAL E MÉTODOS
Presença dos gêneros fúngicos Trichoderma e Fusarium
A coleta de solo foi realizada em estufa e horta, localizadas na área experimental do Departamento de Defesa Fitossanitária da Universidade Federal de Santa Maria. Os locais escolhidos haviam sido utilizados em experimentos com cultivos de olerícolas, incluindo tomateiro e pepineiro, nos quais foram observados sintomas de murcha e podridão de fusário.
O tomateiro e o pepineiro utilizados foram do grupo cereja e de conserva, respectivamente, sendo que as sementes (Bionatur®) não receberam qualquer tipo de tratamento, confirmando-se a ausência de patógenos através de teste de sanidade. As mudas de tomateiro e pepineiro foram cultivadas em substrato comercial (Plantimax®), em bandejas de isopor, em estufa. Aos 30 dias após a semeadura, as mudas foram transplantadas para oito camalhões, em estufa e horta.
Os camalhões mediram 19m de comprimento por 45cm de largura, com espaçamento de 55cm entre os mesmos. Em cada camalhão foram plantadas, intercaladas, oito mudas de tomateiro e oito de pepineiro, em intervalos de 1m, totalizando 16 plantas e um comprimento útil de camalhão de 15m. Ao longo dos oito camalhões, foram transplantadas 64 mudas de tomateiro e 64 de pepineiro, em horta e em estufa.
As plantas da estufa receberam manejo comumente empregado no cultivo local, ou seja, os camalhões foram cobertos por plástico preto (para evitar o desenvolvimento de plantas daninhas) com irrigação diária de 40 min pelo sistema de gotejamento. Os camalhões da horta não foram cobertos por plástico e não receberam irrigação, exceto nos sete dias iniciais, quando as mudas foram regadas diariamente.
Foram coletadas amostras de solo da estufa e da horta e encaminhadas ao Laboratório Central de Análises de Solos (UFSM), para análise química.
A presença dos gêneros Trichoderma e Fusarium foi determinada pela coleta de amostras de solo cultivado com tomateiro e pepineiro, em horta e estufa, ao longo dos oito camalhões. A coleta constou da retirada de 20 amostras de solo rizosférico, para cada cultura, e 20 amostras de solo não-rizosférico (Figura 1).
Para a coleta de solo rizosférico, os pepineiros e tomateiros foram retirados do solo, aos 40 dias após o plantio, com o auxílio de uma pá de corte, retirando-se, com uma leve agitação manual, o solo não aderido às raízes. A parte aérea das plantas foi separada das raízes e desprezada. Os 10cm superiores de raízes foram separados do restante do sistema radicular, colocados em sacos plásticos e agitados até a retirada total do solo rizosférico.
As amostras de solo não-rizosférico foram retiradas com um trado, que foi imerso em hipoclorito de sódio 75% entre amostragens. As amostras foram retiradas na lateral do camalhão, a 50cm de um pepineiro e um tomateiro, na profundidade de 10cm.
Diluições das amostras de solo rizosférico e não-rizosférico foram realizadas para a avaliação da presença dos gêneros Trichoderma e Fusarium. Para as suspensões, foram pesados 10g de solo rizosférico e não-rizosférico de cada amostra e adicionados em frascos de vidro contendo 90mL de água destilada e esterilizada, os quais foram colocados em agitador mecânico durante 30min. Dessa suspensão, foram retirados 5mL e acrescentados em frasco com 95mL de água destilada e esterilizada. Após agitação, uma alíquota de 1mL placa-1 foi espalhada em três placas de Petri contendo meio BDA. Antes de o meio de cultura ser vertido nas placas, adicionou-se, ao mesmo, uma solução de cloranfenicol a 10% (5mL L-1). As placas foram incubadas em câmara climatizada a 24°C, com fotoperíodo de 12h, durante três dias.
Após o período de incubação, os fungos encontrados nas placas foram identificados em nível de gênero, empregando-se bibliografia geral (BARNETT & HUNTER, 1999) e especializada (BISSETT, 1991; VENTURA, 2000).
Utilizou-se o teste dos sinais (CAMPOS, 1983) para comparar o número de amostras que apresentaram os fungos Trichoderma e Fusarium entre solos rizosférico e não-rizosférico e entre os dois ambientes de cultivo (estufa e horta), para o tomateiro e o pepineiro. A presença dos fungos Trichoderma e Fusarium foi considerada positiva (+) e a ausência negativa (-), sendo que a comparação ocorreu entre os positivos.
Patogenicidade dos isolados de Fusarium spp. ao tomateiro e pepineiro
Foram utilizados 46 isolados de Fusarium spp. obtidos de solo não-rizosférico para as duas culturas, e 72 e 73 isolados obtidos de solo rizosférico para o tomateiro e pepineiro, respectivamente.
Cento e dezoito (72 + 46) isolados de Fusarium spp. para o tomateiro e 119 (73 + 46) para o pepineiro foram repicados para placas de Petri contendo o meio NS (NASH & SNYDER, 1962). As placas de Petri foram armazenadas em câmara climatizada a 24°C, com fotoperíodo de 12h, durante sete dias. Foram adicionados 3mL de água destilada e esterilizada em cada placa, e a superfície do meio foi raspada levemente com o auxílio de uma alça de Drigalski, liberando micélio e conídios. Sementes de tomateiro (cereja) e pepineiro (conserva) foram semeadas em substrato comercial Plantimax® em bandejas plásticas medindo 29cm x 43cm. As bandejas ficaram em câmara de crescimento, com temperaturas de 26±2°C, com fotoperíodo de 12h.
Quatorze dias após a semeadura, 354 plântulas de tomateiro e 357 de pepineiro foram transplantadas para bandejas de isopor, de 128 células, preenchidas com substrato comercial. Após o transplante, foi inoculado 1mL da suspensão contendo 106 conídios mL-1 e fragmentos de micélio dos isolados de Fusarium spp., com o auxílio de uma pipeta volumétrica, no colo das plântulas, em direção às raízes (ROSE et al., 2003). Para o tratamento testemunha do tomateiro e pepineiro, foi aplicado 1mL de água destilada e esterilizada.
Foram realizadas três repetições (três plântulas) por tratamento e as bandejas de isopor que continham as plântulas foram acondicionadas em câmara de crescimento com temperaturas de 26±2°C e fotoperíodo de 12h. A avaliação da patogenicidade foi realizada 15 e 30 dias após a inoculação do patógeno e constou da observação de algum dos possíveis sintomas: mudas tombadas, folhas amarelecidas e/ou murchas, colo necrosado e podridão de raiz.
As plântulas inteiras e as raízes (plantas com 30 dias) com sintomas foram coletadas, lavadas com álcool 70%, hipoclorito de sódio 0,5%, três imersões em água destilada e esterilizada e deixadas para secar sobre papel filtro. Posteriormente, essas plântulas/raízes foram colocadas em placas de Petri contendo o meio NS para o isolamento do agente causal da doença. Os isolados de Fusarium spp. patogênicos foram encaminhados ao Laboratório de Pesquisas Micológicas (LAPEMI - UFSM), para a identificação das espécies.
RESULTADOS E DISCUSSÃO
Presença dos gêneros Trichoderma e Fusarium
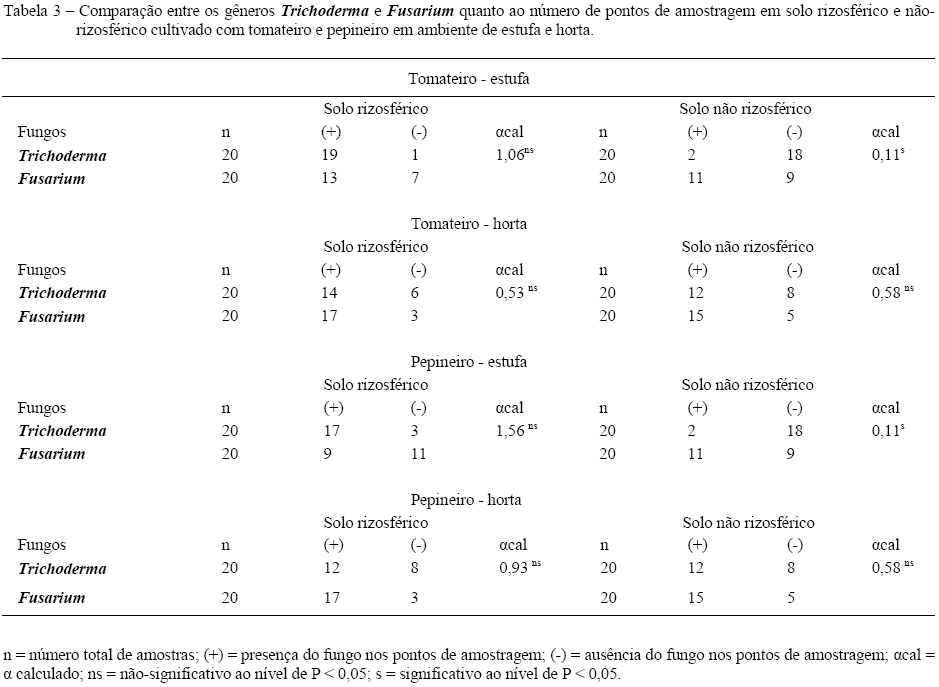
Os gêneros Trichoderma e Fusarium foram encontrados no solo rizosférico e não-rizosférico, cultivado com tomateiro e pepineiro (Figura 1), em ambos os ambientes estudados. Para o tomateiro, em estufa, encontraram-se diferenças significativas, comparando-se o número de pontos de amostragem com a presença de Trichoderma spp., entre solo rizosférico (19 ) e solo não-rizosférico (2) (Tabela 1).
Para o pepineiro, em estufa, também foram encontradas diferenças significativas, quanto à presença de Trichoderma spp., entre o solo rizosférico (17 pontos) e o solo não-rizosférico (2 pontos) (Tabela 1). Na comparação entre solo rizosférico e não-rizosférico, em culturas a campo, SIVAN & CHET (1989) encontraram 5,1 x 106 e 2,0 x 106 UFC (unidade formadora de colônia) g-1 de solo de Trichoderma spp., respectivamente, confirmando maior preferência do fungo pela rizosfera, independentemente do ambiente de cultivo.
Todavia, em horta, tanto para o tomateiro quanto para o pepineiro, não houve diferença significativa quanto à presença de Trichoderma spp. entre solo rizosférico e não-rizosférico (Tabela 1), possivelmente devido à incorporação de restos culturais de cultivos anteriores à implantação das culturas em estudo. Segundo HOWELL (2003), espécies de Trichoderma são facilmente isoladas de solos com restos de vegetais e outras formas de matéria orgânica.
Para o tomateiro e para o pepineiro, foi encontrada diferença significativa quanto à presença de Trichoderma spp., entre os ambientes estufa (2 pontos) e horta (12 pontos), somente em solo não-rizosférico (Tabela 2). Na estufa, a baixa umidade nas laterais dos camalhões, devido à irrigação ser localizada (por gotejamento), aliada à temperatura mais elevada (média de 30°C na estufa e 25°C na horta), pode ter influenciado nesse resultado. O ressecamento do solo não-rizosférico, em estufa, foi observado pela dificuldade no manuseio do trado, o que não aconteceu em solo de horta. SIVAN & CHET (1989) observaram diferenças na colonização das raízes do meloeiro, algodoeiro e tomateiro, por T. harzianum em solo irrigado e não-irrigado, enquanto na escassez de água, mesmo em solo rizosférico, o agente de biocontrole apresentou redução populacional em torno de 40%, demonstrando dificuldade em se manter no ambiente.
Os nutrientes encontrados no solo de horta também podem ter interferido para o melhor desenvolvimento de Trichoderma spp. nesse ambiente. Os solos de horta e estufa apresentaram, características químicas semelhantes, mas ocorreram diferenças quanto ao teor de matéria orgânica (%m V-1): 3,4 e 2,5 e concentração de P (mg L-1): 37,7 e 69,5; K (mg L-1): 152 e 38; e Fe (mg L-1): 210,7 e 115,3, respectivamente. Segundo HUBRARD et al. (1983) a concentração de ferro interferiu no desenvolvimento e estabelecimento de T. hamatum na rizosfera de plantas de ervilha.
Isolados de Fusarium spp. foram encontrados nos dois ambientes, tipos de solo e culturas estudadas, não sendo encontradas diferenças significativas quanto ao número de amostragens que apresentaram o fitopatógeno (Tabela 2).
Entre Trichoderma spp. e Fusarium spp., encontraram-se diferenças significativas apenas em solo não-rizosférico de estufa, para as duas culturas (Tabela 3), pois os dois gêneros foram encontrados em 2 e 11 pontos de amostragem, respectivamente (Figura 1). Pode-se inferir que o fitopatógeno apresentou maior capacidade de resistir nesse ambiente do que Trichoderma spp., já que o solo estava mais seco, o que está relacionado com a formação de clamidósporos. Os clamidósporos representam a principal forma de sobrevivência de espécies de Fusarium no solo, pois resistem à ação de inúmeros fatores adversos, além de poderem permanecer dormentes por longos períodos de tempo na ausência do hospedeiro (AMORIM, 1995).
Patogenicidade dos isolados de Fusarium spp. ao tomateiro e pepineiro
A maior parte dos isolados de Fusarium spp. não foram patogênicos ao tomateiro e pepineiro, sendo que os mesmos podem ser de espécies não-patogênicas ou forma specialis diferentes. Dos 119 isolados de Fusarium spp. utilizados no teste de patogenicidade para o pepineiro, foram encontrados seis patogênicos (5%), sendo três de solo rizosférico de estufa e três de solo rizosférico de horta. Para o tomateiro, dos 118 isolados de Fusarium spp., apenas três de solo rizosférico de estufa e dois de solo rizosférico de horta foram patogênicos, totalizando cinco isolados (4%). Os isolados patogênicos foram identificados como Fusarium oxysporum, confirmando os sintomas de folhas amarelecidas e/ou murchas observados nas plantas.
Embora os isolados de Fusarium patogênicos tenham sido obtidos de solo rizosférico, as plantas das quais foram obtidos não manifestavam sintomas da doença. Pôde-se constatar que, apesar da presença do hospedeiro e do patógeno, o ambiente determinou que a doença não se manifestasse. Os fatores ambientais podem determinar o grau de predisposição do hospedeiro, influenciando desde o estabelecimento da doença numa cultura até o desencadeamento da epidemia (BEDENDO, 1995).
CONCLUSÕES
Em solo de estufa, Trichoderma spp. ocorre com maior freqüência na região rizosférica do tomateiro e do pepineiro, enquanto que, em horta, sua ocorrência é distribuída entre a região rizosférica e não-rizosférica.
Solos rizosféricos e não-rizosféricos de estufa e horta apresentam a mesma freqüência do gênero Fusarium, tanto para o tomateiro quanto para o pepineiro.
A maioria dos isolados de Fusarium spp. obtidos de horta e estufa não são patogênicos aos grupos de tomateiro e pepineiro estudados, sendo que os patogênicos são oriundos da rizosfera.
Recebido para publicação 01.08.06
Aprovado em 25.04.07
- AMORIM, L. Sobrevivência do inóculo. In: BERGAMIN FILHO, A. (Ed.). Manual de fitopatologia São Paulo: Agronômica Ceres, 1995. p.246-267.
- BARNETT, H.L.; HUNTER, B.B. Illustrated genera of imperfect fungi Minnesota: Burgess, 1999. 218p.
- BATEMAN, G.L.; MURRAY, G. Seasonal variations in populations of Fusarium species in wheat-field soil. Applied Soil Ecology, v.18, p.117-128, 2001.
- BEDENDO, I.P. Ambiente e doença. In: BERGAMIN FILHO, A. (Ed.). Manual de fitopatologia São Paulo: Agronômica Ceres, 1995. p.331-341.
- BISSETT, J. A revision of the genus Trichoderma III. Section Pachybasium. Canadian Journal of Botany, v.69, p.2373-2417, 1991.
- CAMPOS, H. Estatística experimental não-paramétrica 4.ed. Piracicaba: USP/ESALQ, 1983. 349p.
- COOK, R.J. Making greater use of introduced microorganisms for biological control of plant pathogen. Annual Review Phytopathology, v.31, p.53-80, 1993.
- FREITAS, T.M.Q. Dano, dinâmica populacional e fontes de resistência genética a Fusarium solani f. sp. glycines 2003. 145f. Dissertação (Mestrado em Agronomia) - Programa de Pós-graduação em Agronomia, Universidade Federal de Santa Maria.
- GESHEVA, V. Rhizosphere microflora of some citrus as a source of antagonistic actinomycetes. European Journal of Soil Biology, v.38, p.85-88, 2002.
- HOWELL, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant Disease, v.87, p.4-10, 2003.
- HUBRARD, J.P. et al. Effect of soilborne Pseudomonas spp. On the biological control agent, Trichoderma hamatum, on pea seeds. Phytopathology, v.73, p.655-659, 1983.
- LARKIN,R.P.; FRAVEL, D.R. Efficacy of various fungal and bacterial biocontrol organisms for control of fusarium wilt of tomato. Plant Disease, v.82, p.1022-1028, 1998.
- NASH, S.M.; SNYDER, W.C. Quantitative estimations by plate counts of propagules of the bean root rot Fusarium in field soils. Phytopathology, v.52, p.567-572, 1962.
- ROSE, S. et al. Efficacy of biological and chemical treatments for control of fusarium root and stem rot in greenhouse cucumber. Plant Disease, v.87, p.1462-1470, 2003.
- RUPE, J.C. et al. Vertical and temporal distribution of Fusarium solani and Heterodera glycines in fields with sudden death syndrome of soybean. Soil Biology and Biochemistry, v.31, p.245-251, 1999.
- SAREMI, H. et al. Temperature effects on the relative abundance of Fusarium species in a model plant-soil ecosystem. Soil Biology and Biochemistry, v.31, p.941-947, 1999.
- SIVAN, A.; CHET I. The possible role of competition between Trichoderma harzianum and Fusarium oxysporum on rhizosphere colonization. Phytopathology, v.79, p.198-203, 1989.
- VENTURA, J.A. Taxonomia de Fusarium e seus segredos. Parte II Chaves para identificação. Revisão Anual de Patologia de Plantas, v.8, p.303-338, 2000.
Datas de Publicação
-
Publicação nesta coleção
14 Dez 2007 -
Data do Fascículo
Fev 2008
Histórico
-
Aceito
25 Abr 2007 -
Recebido
01 Ago 2006