ABSTRACT:
Spodoptera frugiperda (J. E. Smith) is the main target pest of Bt corn technologies, such as YieldGard VT PRO(tm) (Cry1A.105/Cry2Ab2) and PowerCore(tm) (Cry1A.105/Cry2Ab2/Cry1F). In this study, it was evaluated the biological activity of Bt proteins expressed in different plant structures of YieldGard VT PRO(tm) and PowerCore(tm) corn against S. frugiperda . Complete mortality ofS. frugiperda neonates was observed on leaf-disc of both Bt corn technologies. However, the mortality in silks and grains was lower than 50 and 6%, respectively. In addition, more than 49% of the surviving larvae in silks and grains completed the biological cycle. However, all life table parameters were negatively affected in insects that developed in silks and grains of both Bt corn events. In summary, the low biological activity of Bt proteins expressed on silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) corn can contribute to the resistance evolution inS. frugiperda populations.
Key words:
fall armyworm;
Bacillus thuringiensis
; Bt corn; insect resistance management
RESUMO:
Spodoptera frugiperda (J. E. Smith) (Lepidoptera:Noctuidae ) é a principal praga-alvo dos eventos de milho YieldGard VT PRO(tm) (Cry1A.105/Cry2Ab2) e PowerCore(tm) (Cry1A.105/Cry2Ab2/Cry1F). Para subsidiar o manejo de resistência, avaliou-se a atividade biológica das proteínas de Bt expressas em diferentes estruturas dessas tecnologias de milho Bt contra S. frugiperda . Em discos de folha de milho YieldGard VT PRO(tm) e PowerCore(tm), houve mortalidade completa de neonatas deS. frugiperda . No entanto, em estilo-estigmas e grãos, a mortalidade foi inferior a 50% e 6%, respectivamente. Em adição, mais de 49% das larvas sobreviventes em estilos-estigmas e grãos completaram o ciclo biológico. No entanto, todos os parâmetros de tabela de vida de fertilidade foram negativamente afetados nos insetos que se desenvolveram em estilos-estigmas e grãos de milho Bt. A baixa atividade biológica das proteínas Bt expressas em estilos-estigmas e grãos de milho YieldGard VT PRO(tm) e PowerCore(tm) pode contribuir para a evolução de resistência em populações deS. frugiperda .
Palavras-chave:
lagarta-do-cartucho;
Bacillus thuringiensis
; milho Bt; manejo da resistência de insetos
INTRODUCTION
The fall armyworm, Spodoptera frugiperda (J. E. Smith 1797) (Lepidoptera: Noctuidae ) is the primary pest of corn in South American countries (POGUE, 2002POGUE M.G. A world revision of the genus Spodoptera Gueneé (Lepidoptera: Noctuidae). Memoirs of the American Entomological Society, v.43, p.1-201, 2002. Available from: <Available from: http://ia700806.us.archive.org/7/items/memoirsofameric432002amer/memoirsofameric432002amer.pdf
>. Accessed: Nov. 18, 2014.
http://ia700806.us.archive.org/7/items/m...
). This species is the most destructive pest of corn in Brazil (BARROS et al., 2010BARROS, E.M. et al. Oviposition, development and reproduction of Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) fed on different hosts of economic importance. Neotropical Entomology, v.39, n.6, p.996-1001, 2010. Available from: <Available from: http://dx.doi.org/10.1590/S1519-566X2010000600023
>. Accessed: Nov. 10, 2014. doi: 10.1590/S1519-566X2010000600023.
http://dx.doi.org/10.1590/S1519-566X2010...
). Besides defoliation, S. frugiperda also attacks corn ears, where neonates initially feed on corn silks and, subsequently penetrate the corn ears and attack the grains (CRUZ, 1995CRUZ, I. A lagarta-do-cartucho na cultura do milho. Sete Lagoas: Embrapa - Embrapa Milho e Sorgo, 1995. 45p. (Circular Técnica, 21). ). This attack allows the entry of other insects and pathogens that causes indirect damages to the grains (CRUZ & TURPIN et al., 1983CRUZ, I.; TURPIN, F.T. Yield impact of larval infestations of fall armyworm (Lepidoptera: Noctuidae) to midwhorl growth stage of corn. Journal of Economic Entomology, v.76, n.5, p. 1052-1054, 1983. Available from: <Available from: http://dx.doi.org/10.1093/jee/76.5.1052
>. Accessed: Nov. 11, 2014. doi: 10.1093/jee/76.5.1052.
http://dx.doi.org/10.1093/jee/76.5.1052...
).
In Brazil, the main control tactic of S. frugiperda involves the use of Bt corn, which are cultivated in more than 80% of the current area of this crop (CÉLERES, 2015CÉLERES, 2015. 2nd follow-up on agricultural biotechnology adoption for the 2014/15 crop. Available from: <Available from: http://www.celeres.com.br/
>. Accessed: May. 20, 2015.
http://www.celeres.com.br/...
). Among Bt corn technologies, YieldGardVT PRO(tm) (Cry1A.105/Cry2Ab2) and PowerCore(tm) (Cry1A.105/Cry2Ab2/Cry1F) are used to S. frugiperda control (CTNBIO, 2010CTNBIO (COMISSÃO TÉCNICA NACIONAL DE BIOSSEGURANÇA). Commercial release of insect resistant and herbicide tolerant maize containing events MON 89034 × TC1507 × NK603. Technical Opinion n. 2753/2010. Available from: <Available from: http://www.ctnbio.gov.br/index.php/content/view/17032.html
>. Accessed: Nov. 22, 2014.
http://www.ctnbio.gov.br/index.php/conte...
). Corn hybrids expressing two or more Bt proteins are usually more effective in the control of S. frugiperda , than plants expressing a single Bt protein, especially when proteins had low potential for cross-resistance (ZHAO et al., 2003ZHAO, J.Z. et al. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nature Biotechnology, v.21, n.12, p.1493-1497, 2003. Available from: <Available from: http://www.nature.com/nbt/journal/v21/n12/pdf/nbt907.pdf
>. Accessed: Dec. 02, 2014. doi:10.1038/nbt907.
http://www.nature.com/nbt/journal/v21/n1...
; HERNÁNDEZ-RODRÍGUEZ et al., 2013HERNÁNDEZ-RODRÍGUEZ, C.S. et al. Shared midgut binding sites for Cry1A.105, Cry1Aa, Cry1Ab, Cry1Ac and Cry1Fa proteins from Bacillus thuringiensis in two important corn pests, Ostrinia nubilalis and Spodoptera frugiperda. PLoS ONE, v.8, n.7, p.e68164, 2013. Available from: <Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0068164
>. Accessed: Nov. 15, 2014. doi: 10.1371/journal.pone.0068164.
http://journals.plos.org/plosone/article...
).
In Brazil, the intensive crop production system and the high adoption of Bt corn technology exposes S. frugiperda populations to high selection pressure, which favor the resistance evolution. In this scenario, S. frugiperda field-evolved resistance to the Cry1F protein expressed in Herculex(tm) (TC1507) corn event (FARIAS et al., 2014FARIAS, J.R. et al. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Protection, v.64, n.2, p.150-158, 2014a. Available from: <Available from: http://dx.doi.org/10.1016/j.cropro.2014.06.019
>. Accessed: Dec. 08, 2014. doi:10.1016/j.cropro.2014.06.019.
http://dx.doi.org/10.1016/j.cropro.2014....
). S. frugiperda also evolved resistance to Cry1F corn in Puerto Rico (STORER et al., 2010STORER, N.P. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. Journal of Economic Entomology, v.103, n.4, p.1031-1038, 2010. Available from: <Available from: http://dx.doi.org/10.1603/EC10040
>. Accessed: Dec. 02, 2014. doi: 10.1603/EC10040.
http://dx.doi.org/10.1603/EC10040...
), and recently Cry1F resistant fall armyworm has been found in the Southern US (HUANG et al., 2014HUANG F. et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: single gene versus pyramided Bt maize. PLoS ONE, v.11, e112958, 2014. Available from: <Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4234506/pdf/pone.0112958.pdf
>. Accessed: Dec. 15, 2014. doi: 10.1371/journal.pone.0112958.
http://www.ncbi.nlm.nih.gov/pmc/articles...
).
Another aspect that may affect the performance of Bt plants in the control of target pests is the variation in the Bt protein expression throughout plant development and plant structures (HUANG et al., 2011HUANG, F. et al. F2 screening for resistance to pyramided Bacillus thuringiensis maize in Louisiana and Mississippi populations of Diatraea saccharalis (Lepidoptera: Crambidae). Pest Management Science, v.67, n.10, p.1269-1276, 2011. Available from: <Available from: http://onlinelibrary.wiley.com/doi/10.1002/ps.2182/epdf
>. Accessed: Dec. 15, 2014. doi: 10.1002/ps.2182.
http://onlinelibrary.wiley.com/doi/10.10...
). This potentially exposes target pests to sub-lethal doses and negatively affects the mortality response (HUANG et al., 2011HUANG, F. et al. F2 screening for resistance to pyramided Bacillus thuringiensis maize in Louisiana and Mississippi populations of Diatraea saccharalis (Lepidoptera: Crambidae). Pest Management Science, v.67, n.10, p.1269-1276, 2011. Available from: <Available from: http://onlinelibrary.wiley.com/doi/10.1002/ps.2182/epdf
>. Accessed: Dec. 15, 2014. doi: 10.1002/ps.2182.
http://onlinelibrary.wiley.com/doi/10.10...
). The variation in Bt protein expression can favor the survival of target pests and contribute to the resistance evolution, because among surviving insects there is the heterozygous individuals, the most common carriers of resistance alleles (GOULD, 1998GOULD, F. Sustainability of transgenic insecticidal cultivars: integrating pest genetics and ecology. Annual Review of Entomology, v.43, p.701-726, 1998. Available from: <Available from: http://www.annualreviews.org/doi/abs/10.1146/annurev.ento.43.1.701
>. Accessed: Dec. 08, 2014. doi: 10.1146/annurev.ento.43.1.701.
http://www.annualreviews.org/doi/abs/10....
; ANDOW, 2008ANDOW, D.A. The risk of resistance evolution in insects to transgenic insecticidal crops. Collection of Biosafety Reviews, v.4, p.142-199, 2008. Available from: <Available from: http://biosafety.icgeb.org/sites/default/files/Andow.pdf
>. Accessed: Nov. 13, 2014.
http://biosafety.icgeb.org/sites/default...
).
To support resistance management strategies, the biological activity of Btproteins expressed in leaves, silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) corn technologies against S. frugiperda were evaluated.
MATERIALS AND METHODS
Insects and corn plants
In the bioassays were used a susceptible reference population of S. frugiperda, which were kept in the laboratory for >10 yr, free of selection pressure by Bt proteins. In all studies, YieldGard VT PRO(tm) (LH198 + LH172) and PowerCore(tm) (2B633) corn technologies expressing Cry1A.105/Cry2Ab2 and Cry1A.105/Cry2Ab2/Cry1F, respectively, were used. As a control treatment, corn plants (DKB(tm) 390) without Bt proteins expression, were used. To obtain leaves, silks and grains, YieldGard VT PRO(tm), PowerCore(tm) and non-Bt corn was cultivated in separately small plots (4m width × 4m length) with 6 rows/plot (5 plants per m). When the emergence of silks started, all corn ears were covered with a brown paper bag to prevent cross-pollination. Later, pollination was artificially performed using pollen from tassels of the same corn hybrid, simulating natural pollination in the field.
Leaf-disc, silk and grain bioassays
To perform leaf-disc bioassays, fully expanded leaves were removed from the corn whorl at V3 and V8 stages (MAGALHÃES & DURÃES, 2006MAGALHÃES, P.C.; DURÃES, F.O.M. Fisiologia da Produção de Milho. Sete Lagoas: Embrapa Milho e Sorgo, 2006. 10p. (Circular Técnica, 76).). In laboratory, leaf discs (2.4cm in diameter) were cut using a metallic cutter, and placed on a gelled mixture of water-agar 2.5% (1mL per well) in 12-well acrylic plates (Corning Incorporated, New York, NY). Leaf discs were separated from the water-agar layer by a filter paper disc. Later, one neonate ofS. frugiperda (<24h old) was infested in each well using a fine paintbrush. Bioassays were also performed with silks at phenological stage R1 (silking) and grains at stage R3 (milk grains). Silks were removed (cut) from the corn ears, and placed on a gelled mixture of water-agar and filter paper in 6-well acrylic plates (Corning Incorporated, New York, NY) (3g/well). In bioassays with grains, these structures were carefully removed from the corn ears and placed on acrylic plates using the same methodology described above. Later, three neonates ofS. frugiperda (<24 h old) were infested in each well. All plates were sealed with transparent plastic film (Magipack(r)), covered by the plate lid and kept in a climatic chamber at 27±1°C, 60±10% RH and 14h photoperiod. The experimental design was completely randomized with 10, 5 and 5 replicates for leaf-disc, silk and grains bioassays. In each replicate, 12, 18 and 18 larvae were tested, respectively. Mortality was measured at five days. Mortality data of S. frugiperda in YieldGard VT PRO(tm) and PowerCore(tm) were corrected based on non-Bt corn using Abbott's formula (ABBOTT, 1925ABBOTT, W.S. A method of computing the effectiveness of an insecticide. Journal of Economic Entomology, v.18, n.1, p.265-267, 1925. Available from: <Available from: http://dx.doi.org/10.1093/jee/18.2.265a265-267
>. Accessed: Nov. 15, 2014. doi: 10.1093/jee/18.2.265a265-267.
http://dx.doi.org/10.1093/jee/18.2.265a2...
). Percent mortality (x) was transformed to , and subjected to an analysis of variance (ANOVA), and the means were compared using Tukey's test (P≤0.05; PROC ANOVA, SAS INSTITUTE, 2000SAS INSTITUTE INC. Statistical analysis system: getting started with the SAS learning. Cary, NC. 2000.).
Biological parameters of S. frugiperda fed in silks and grains of Bt and non-Bt corn
The same Bt corn hybrids used above was also used in this study. At phenological stage R1, silks were removed from corn ears, and they were placed on a gelled mixture of agar-water at 2.5% in 16-well bioassay trays (Advento do Brasil, São Paulo, Brazil), and were separated from the agar-water layer by filter paper. Later, five neonates ofS. frugiperda (<24h old) were infested in each well. Larvae were fed with silks until the 2nd larval instar, when the silks were replaced by grains, which were removed from corn ears at phenological stage R3. Larvae were fed with grains until the pupa stage. Silks and grains were replaced every 48h. The experimental design was completely randomized with 20 replicates per treatment (10 larvae per replicate), and bioassay trays was placed in a climatic chamber at 27±1°C, 60±10% relative humidity and 14h photoperiod. The following biological parameters were assessed: duration and survival rates egg, larval and pupal periods; total cycle duration (egg to adult); larval weight at 12 days; pupal weight at 24h after pupation; sex ratio; duration of pre-oviposition, oviposition and post-oviposition periods; male and female longevity; fecundity; and fertility. Egg viability and duration of egg, larval and pupal periods and total cycle were determined in daily observations. Female longevity and fecundity were evaluated from the formation of 15 pairs per treatment that were kept in PVC cages (23cm height × 10cm diameter) internally coated by paper towel (oviposition substrate) and closed at the top with a voile-type fabric. Adults were fed with a 10% honey/aqueous solution provided on cotton. Number of eggs and mortality of adults were assessed. To determine the embryonic period and viability, 50 eggs were obtained from the second oviposition of each pair. Eggs were placed into glass tubes with flat bottoms (8.5×2.5cm). A piece of paper moistened with distilled water daily was placed inside the tube, which was closed with plastic film. Eggs and the number of larvae hatched were counted daily. Duration of larva and pupal stages, larval and pupal weight, pre-oviposition, oviposition and post-oviposition periods were transformed using arcsen , because of non-normal distributions of residuals. Later, all data were subjected to an analysis of variance (ANOVA), and the means were compared using Tukey test (P≤0.05; PROC ANOVA, SAS INSTITUTE, 2000SAS INSTITUTE INC. Statistical analysis system: getting started with the SAS learning. Cary, NC. 2000.). Putative deviation in the sex ratio was compared using the Chi-squared test (χ2) (P≤0.05; PROC FREQ, SAS INSTITUTE, 2000SAS INSTITUTE INC. Statistical analysis system: getting started with the SAS learning. Cary, NC. 2000.). In addition, a fertility life table was calculated by estimating the mean generation time (T), the net reproductive rate (Ro), the intrinsic rate of increase (rm) and the finite rate of increase (λ). Life table parameters were estimated by the "Jackknife" method using "lifetable.sas" procedure (MAIA et al., 2000MAIA, A.H.N. et al. Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. Journal of Economic Entomology, v.93, n.2, p.511-518, 2000. Available from: <Available from: http://dx.doi.org/10.1603/0022-0493-93.2.511
>. Accessed: Nov. 22, 2014. doi: 10.1603/0022-0493-93.2.511.
http://dx.doi.org/10.1603/0022-0493-93.2...
) in SAS(r) software (SAS INSTITUTE, 2000SAS INSTITUTE INC. Statistical analysis system: getting started with the SAS learning. Cary, NC. 2000.).
RESULTS
Leaf-disc, silk and grain bioassays
There was complete mortality of S. frugiperda neonates in leaf discs of YieldGard VT PRO(tm) and PowerCore(tm) corn at V3 (F=138.47; df= 6, 28; P<0.0001) and V8 (F=70.53; df= 6, 28; P<0.0001). In contrast, in non-Bt corn the mortality was <15%. In silks of YieldGard VT PRO(tm) and PowerCore(tm) corn, mortality of S. frugiperda neonates was near 25 and 50% (F=42.53; df=2, 12; P<0.0001). In grains of both Bt corn, the mortality was near 5% (F=2.53; df =2, 12; P=0.2680). In silks and grains of non-Bt corn the mortality was near 5%.
Biological parameters of S. frugiperda fed in silks and grains of Bt and non-Bt corn
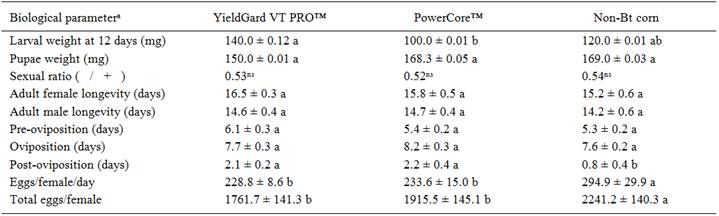
There was no significant difference in the duration of egg (F=0.14; df=2, 29; P=0.8702), larval (F=0.74; df=2, 29; P=0.4874), pupal (F=0.29; df=2, 29; P=0.7526) and egg-adult period (F=1.58; df=2, 29; P=0.2236) of S. frugiperda fed in silks and grains of YieldGard VT PRO(tm), PowerCore(tm) and non-Bt corn (Figure 1). No significant differences were also detected in the survival of egg (F=0.18; df=2, 29; P=0.6502) and pupal (F=0.77; df=2, 29; P=0.4732) stages in both Bt corn technologies (Figure 1). However, larval survival was significantly lower in silks and grains of YieldGard VT PRO(tm) (62%) and PowerCore(tm) (50%) corn, when compared with non-Bt corn (69%) (F=12.12; df=2, 29; P=0.1238). Lower larval survival negatively affected the survival of egg-adult period, with 54 and 44% of insects completing the biological cycle in YieldGard VT PRO(tm) and PowerCore(tm) corn, respectively. In contrast, the survival in non-Bt corn was 62% (F=12.28; df=2, 29; P=0.0253). There was also a significant reduction in the larval weight of S. frugiperda fed in silks and grains of PowerCore(tm) corn compared with the non-Bt corn (F=2.98; df=2, 29; P=0.0511) (Table 1). However, the larval weight in silks and grains of YieldGard VT PRO(tm) corn was similar to that in non-Bt corn (Table 1). The differences in larval weight did not affect pupae weight (F=0.51; df=2, 29; P=0.6057) (Table 1). Sex ratio was also not affected in Bt corn (χ2=18.78; df=2, 29; P=0.2802) (Table 1). Adults of S. frugiperda originating from larvae fed in silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) showed similar female (F=2.02; df=2, 40; P=0.1116) and male (F=2.42; df=2, 40; P=0.0921) longevity; pre-oviposition (F=1.01; df=2, 40; P=0.3723); oviposition (F=0.04; df=2, 40; P=0.9573) and post-oviposition (F=2.84; df=2, 40; P=0.0534) periods, those from non-Bt corn (Table 1). However, females ofS. frugiperda obtained from larvae fed in silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) showed 20% reduction in the eggs/day (F=5.00; df=2, 40; P=0.0118) and total number of eggs (F=4.68; df=2, 40; P=0.0153) (Table 1).

Duration and survival of the life stages of Spodoptera frugiperda fed in silks and grains of Bt and non-Bt corn. Bars with the same letter are not significantly different by Tukey's test (P<0.05).
The life table parameters of S. frugiperda were negatively affected when fed in silks and grains of Bt corn technologies (Table 2). The mean generation time (T) was significantly longer (2 days) for S. frugiperda fed in silks and grains of Bt corn compared to non-Bt corn. Furthermore, feeding S. frugiperda in silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) reduced the net reproductive rate (Ro ) by 27 and 42%, respectively (Table 2). Based on these results, it was estimated that after 42 days (T) of S. frugiperda development in silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) approximately 726 and 568 females are expected of eachfemale, respectively, where on non-Bt corn 1008 females are expected after 40 days (T). Furthermore, S. frugiperda fedin reproductive structures of YieldGard VT PRO(tm) and PowerCore(tm) corn reduced the intrinsic rate of increase (rm ) by 10 and 15%, respectively. The finite rate of increase (λ) also was lower in both Bt corn (Table 2).
DISCUSSION
The Bt proteins expressed on leaves of YieldGard VT PRO(tm) (Cry1A.105/Cry2Ab2) and PowerCore(tm) (Cry1A.105/Cry2Ab2/Cry1F) corn showed high toxicity against S. frugiperda . However, S. frugiperda neonates presented high larval survival in silks and grains of both Bt corn technologies. This larval survival can be explained by the less expression of Bt proteins in reproductive structures (HUANG et al., 2011). For resistance management, the larval survival of S. frugiperda on corn ears can favor the resistance evolution, because it allows the survival of heterozygous individuals in the reproductive structures, favoring an increase in the frequency of these insects. Survival of heterozygous insects on silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) may also compromise the future efficacy of these technologies. This was demonstrated in Helicoverpa zea (Boddie) that increased larval survival on Bt cotton (Cry1Ac/Cry2Ab2) (BRÉVAULT et al., 2013BRÉVAULT, T. et al. Potential shortfall of pyramided transgenic cotton for insect resistance management. Proceedings of the National Academy of Sciences of the United States of America, v.110, n.15, p.5806-5811, 2013. Available from: <Available from: http://www.pnas.org/content/110/15/5806
>. Accessed: Nov. 11, 2014. doi: 10.1073/pnas.1216719110.
http://www.pnas.org/content/110/15/5806...
).
Field-evolved resistance of S. frugiperda to Cry1F expressed in Herculex(r) (TC1507) corn in Brazil (FARIAS et al., 2014FARIAS, J.R. et al. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Protection, v.64, n.2, p.150-158, 2014a. Available from: <Available from: http://dx.doi.org/10.1016/j.cropro.2014.06.019
>. Accessed: Dec. 08, 2014. doi:10.1016/j.cropro.2014.06.019.
http://dx.doi.org/10.1016/j.cropro.2014....
) is an aspect that can also reduce the efficacy of YieldGard VT PRO(tm) and PowerCore(tm) corn against this species. This aspect, in association with the low expression of Cry1A.105, Cry2Ab2 and Cry1F proteins in reproductive structures, can favor an increase in the frequency of heterozygous and resistant insects in field populations of S. frugiperda . Furthermore, the high probability of cross-resistance between Cry1F and Cry1A.105, which share the same binding site, also contributes to the resistance evolution (HERNÁNDEZ-RODRÍGUEZ et al., 2013HERNÁNDEZ-RODRÍGUEZ, C.S. et al. Shared midgut binding sites for Cry1A.105, Cry1Aa, Cry1Ab, Cry1Ac and Cry1Fa proteins from Bacillus thuringiensis in two important corn pests, Ostrinia nubilalis and Spodoptera frugiperda. PLoS ONE, v.8, n.7, p.e68164, 2013. Available from: <Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0068164
>. Accessed: Nov. 15, 2014. doi: 10.1371/journal.pone.0068164.
http://journals.plos.org/plosone/article...
). In this context, the long-term efficacy of YieldGard VT PRO(tm) and PowerCore(tm) corn technologies is partly dependent of the Cry2Ab2 protein, which does not exhibit cross-resistance with Cry1 proteins (FERRÉ & VAN RIE, 2002FERRÉ J.; VAN RIE, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annual Review Entomology, v.47, p.501-533, 2002. Available from: <Available from: http://www.annualreviews.org/doi/full/10.1146/annurev.ento.47.091201.145234
>. Accessed: Nov. 12, 2014. doi: 10.1146/annurev.ento.47.091201.145234.
http://www.annualreviews.org/doi/full/10...
; CACCIA et al., 2010CACCIA, S. et al. Target site alteration is responsible for field-derived resistance to Bacillus thuringiensis Cry2A insecticidal proteins in Helicoverpa spp. PLoS ONE, v.5, p.e9975, 2010. Available from: <Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2848615/
>. Accessed: Nov. 11, 2014. doi: 10.1371/journal.pone.0009975.
http://www.ncbi.nlm.nih.gov/pmc/articles...
). However, Cry2Ab2 protein showed low toxicity against S. frugiperda (SIVASUPRAMANIAM et al., 2008SIVASUPRAMANIAM, S.W. et al. Toxicity and characterization of cotton expressing Bacillus thuringiensis Cry1Ac and Cry2Ab2 proteins for control of lepidopteran pests. Journal of Economic Entomology, v.101, n.2, p.546-554, 2008. Available from: <Available from: http://www.bioone.org/doi/full/10.1603/0022-0493%282008%29101%5B546%3ATACOCE%5D2.0.CO %3B2
>. Accessed: Nov. 15, 2014. doi: 10.1603/0022-0493(2008)101[546:TACOCE]2.0.CO;2.
http://www.bioone.org/doi/full/10.1603/0...
).
Results reported in this study showed that the survival of S. frugiperda in corn ears of YieldGard VT PRO(tm) and PowerCore(tm) can represent a potential factor that favor the resistance evolution. However, to implement more effective resistance management strategies is essential understanding the selection pressure that populationsof S. frugiperda will be exposed in a region and crop production system. Furthermore, our results were obtained in bioassays conducted in the laboratory, without occurrence of other natural mortality factors. Other factors to be considered are age and closure of corn ears, which have a direct effect on the access of larvae to grains and vary drastically among different available corn hybrids.
CONCLUSIONS
The biological activity of Bt proteins expressed in YieldGard VT PRO(tm) and PowerCore(tm) corn against S. frugiperda is high in leaves and low in silks and grains. However, the expression of Bt proteins in silks and grains of YieldGard VT PRO(tm) and PowerCore(tm) negatively affect the population growth parameters of S. frugiperda .
ACKNOWLEDGEMENTS
The authors are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for granting scholarship to the senior author and financial support of this study (Process number 141618/2011-6).
REFERENCES:
- ABBOTT, W.S. A method of computing the effectiveness of an insecticide. Journal of Economic Entomology, v.18, n.1, p.265-267, 1925. Available from: <Available from: http://dx.doi.org/10.1093/jee/18.2.265a265-267 >. Accessed: Nov. 15, 2014. doi: 10.1093/jee/18.2.265a265-267.
» https://doi.org/10.1093/jee/18.2.265a265-267.» http://dx.doi.org/10.1093/jee/18.2.265a265-267 - ANDOW, D.A. The risk of resistance evolution in insects to transgenic insecticidal crops. Collection of Biosafety Reviews, v.4, p.142-199, 2008. Available from: <Available from: http://biosafety.icgeb.org/sites/default/files/Andow.pdf >. Accessed: Nov. 13, 2014.
» http://biosafety.icgeb.org/sites/default/files/Andow.pdf - BARROS, E.M. et al. Oviposition, development and reproduction of Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) fed on different hosts of economic importance. Neotropical Entomology, v.39, n.6, p.996-1001, 2010. Available from: <Available from: http://dx.doi.org/10.1590/S1519-566X2010000600023 >. Accessed: Nov. 10, 2014. doi: 10.1590/S1519-566X2010000600023.
» https://doi.org/10.1590/S1519-566X2010000600023.» http://dx.doi.org/10.1590/S1519-566X2010000600023 - BRÉVAULT, T. et al. Potential shortfall of pyramided transgenic cotton for insect resistance management. Proceedings of the National Academy of Sciences of the United States of America, v.110, n.15, p.5806-5811, 2013. Available from: <Available from: http://www.pnas.org/content/110/15/5806 >. Accessed: Nov. 11, 2014. doi: 10.1073/pnas.1216719110.
» https://doi.org/10.1073/pnas.1216719110.» http://www.pnas.org/content/110/15/5806 - CACCIA, S. et al. Target site alteration is responsible for field-derived resistance to Bacillus thuringiensis Cry2A insecticidal proteins in Helicoverpa spp. PLoS ONE, v.5, p.e9975, 2010. Available from: <Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2848615/ >. Accessed: Nov. 11, 2014. doi: 10.1371/journal.pone.0009975.
» https://doi.org/10.1371/journal.pone.0009975.» http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2848615/ - CÉLERES, 2015. 2nd follow-up on agricultural biotechnology adoption for the 2014/15 crop. Available from: <Available from: http://www.celeres.com.br/ >. Accessed: May. 20, 2015.
» http://www.celeres.com.br/ - CRUZ, I. A lagarta-do-cartucho na cultura do milho. Sete Lagoas: Embrapa - Embrapa Milho e Sorgo, 1995. 45p. (Circular Técnica, 21).
- CRUZ, I.; TURPIN, F.T. Yield impact of larval infestations of fall armyworm (Lepidoptera: Noctuidae) to midwhorl growth stage of corn. Journal of Economic Entomology, v.76, n.5, p. 1052-1054, 1983. Available from: <Available from: http://dx.doi.org/10.1093/jee/76.5.1052 >. Accessed: Nov. 11, 2014. doi: 10.1093/jee/76.5.1052.
» https://doi.org/10.1093/jee/76.5.1052.» http://dx.doi.org/10.1093/jee/76.5.1052 - CTNBIO (COMISSÃO TÉCNICA NACIONAL DE BIOSSEGURANÇA). Commercial release of insect resistant and herbicide tolerant maize containing events MON 89034 × TC1507 × NK603. Technical Opinion n. 2753/2010. Available from: <Available from: http://www.ctnbio.gov.br/index.php/content/view/17032.html >. Accessed: Nov. 22, 2014.
» http://www.ctnbio.gov.br/index.php/content/view/17032.html - FARIAS, J.R. et al. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Protection, v.64, n.2, p.150-158, 2014a. Available from: <Available from: http://dx.doi.org/10.1016/j.cropro.2014.06.019 >. Accessed: Dec. 08, 2014. doi:10.1016/j.cropro.2014.06.019.
» https://doi.org/10.1016/j.cropro.2014.06.019» http://dx.doi.org/10.1016/j.cropro.2014.06.019 - FERRÉ J.; VAN RIE, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annual Review Entomology, v.47, p.501-533, 2002. Available from: <Available from: http://www.annualreviews.org/doi/full/10.1146/annurev.ento.47.091201.145234 >. Accessed: Nov. 12, 2014. doi: 10.1146/annurev.ento.47.091201.145234.
» https://doi.org/10.1146/annurev.ento.47.091201.145234.» http://www.annualreviews.org/doi/full/10.1146/annurev.ento.47.091201.145234 - GOULD, F. Sustainability of transgenic insecticidal cultivars: integrating pest genetics and ecology. Annual Review of Entomology, v.43, p.701-726, 1998. Available from: <Available from: http://www.annualreviews.org/doi/abs/10.1146/annurev.ento.43.1.701 >. Accessed: Dec. 08, 2014. doi: 10.1146/annurev.ento.43.1.701.
» https://doi.org/10.1146/annurev.ento.43.1.701.» http://www.annualreviews.org/doi/abs/10.1146/annurev.ento.43.1.701 - HERNÁNDEZ-RODRÍGUEZ, C.S. et al. Shared midgut binding sites for Cry1A.105, Cry1Aa, Cry1Ab, Cry1Ac and Cry1Fa proteins from Bacillus thuringiensis in two important corn pests, Ostrinia nubilalis and Spodoptera frugiperda. PLoS ONE, v.8, n.7, p.e68164, 2013. Available from: <Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0068164 >. Accessed: Nov. 15, 2014. doi: 10.1371/journal.pone.0068164.
» https://doi.org/10.1371/journal.pone.0068164.» http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0068164 - HUANG, F. et al. F2 screening for resistance to pyramided Bacillus thuringiensis maize in Louisiana and Mississippi populations of Diatraea saccharalis (Lepidoptera: Crambidae). Pest Management Science, v.67, n.10, p.1269-1276, 2011. Available from: <Available from: http://onlinelibrary.wiley.com/doi/10.1002/ps.2182/epdf >. Accessed: Dec. 15, 2014. doi: 10.1002/ps.2182.
» https://doi.org/10.1002/ps.2182.» http://onlinelibrary.wiley.com/doi/10.1002/ps.2182/epdf - HUANG F. et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: single gene versus pyramided Bt maize. PLoS ONE, v.11, e112958, 2014. Available from: <Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4234506/pdf/pone.0112958.pdf >. Accessed: Dec. 15, 2014. doi: 10.1371/journal.pone.0112958.
» https://doi.org/10.1371/journal.pone.0112958» http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4234506/pdf/pone.0112958.pdf - MAGALHÃES, P.C.; DURÃES, F.O.M. Fisiologia da Produção de Milho. Sete Lagoas: Embrapa Milho e Sorgo, 2006. 10p. (Circular Técnica, 76).
- MAIA, A.H.N. et al. Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. Journal of Economic Entomology, v.93, n.2, p.511-518, 2000. Available from: <Available from: http://dx.doi.org/10.1603/0022-0493-93.2.511 >. Accessed: Nov. 22, 2014. doi: 10.1603/0022-0493-93.2.511.
» https://doi.org/10.1603/0022-0493-93.2.511.» http://dx.doi.org/10.1603/0022-0493-93.2.511 - POGUE M.G. A world revision of the genus Spodoptera Gueneé (Lepidoptera: Noctuidae). Memoirs of the American Entomological Society, v.43, p.1-201, 2002. Available from: <Available from: http://ia700806.us.archive.org/7/items/memoirsofameric432002amer/memoirsofameric432002amer.pdf >. Accessed: Nov. 18, 2014.
» http://ia700806.us.archive.org/7/items/memoirsofameric432002amer/memoirsofameric432002amer.pdf - SAS INSTITUTE INC. Statistical analysis system: getting started with the SAS learning. Cary, NC. 2000.
- SIVASUPRAMANIAM, S.W. et al. Toxicity and characterization of cotton expressing Bacillus thuringiensis Cry1Ac and Cry2Ab2 proteins for control of lepidopteran pests. Journal of Economic Entomology, v.101, n.2, p.546-554, 2008. Available from: <Available from: http://www.bioone.org/doi/full/10.1603/0022-0493%282008%29101%5B546%3ATACOCE%5D2.0.CO %3B2 >. Accessed: Nov. 15, 2014. doi: 10.1603/0022-0493(2008)101[546:TACOCE]2.0.CO;2.
» https://doi.org/10.1603/0022-0493(2008)101[546:TACOCE]2.0.CO;2.» http://www.bioone.org/doi/full/10.1603/0022-0493%282008%29101%5B546%3ATACOCE%5D2.0.CO %3B2 - STORER, N.P. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. Journal of Economic Entomology, v.103, n.4, p.1031-1038, 2010. Available from: <Available from: http://dx.doi.org/10.1603/EC10040 >. Accessed: Dec. 02, 2014. doi: 10.1603/EC10040.
» https://doi.org/10.1603/EC10040.» http://dx.doi.org/10.1603/EC10040 - ZHAO, J.Z. et al. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nature Biotechnology, v.21, n.12, p.1493-1497, 2003. Available from: <Available from: http://www.nature.com/nbt/journal/v21/n12/pdf/nbt907.pdf >. Accessed: Dec. 02, 2014. doi:10.1038/nbt907.
» https://doi.org/10.1038/nbt907» http://www.nature.com/nbt/journal/v21/n12/pdf/nbt907.pdf
-
1
CR-2015-0047.R2
Publication Dates
-
Publication in this collection
June 2016
History
-
Received
18 Jan 2015 -
Accepted
07 July 2015 -
Reviewed
24 Mar 2016