Resumos
Foram analisados e comparados os processos de germinação dos isolados ARS-1590, ARS-1261 e ARS-1229 de Zoophthora radicans (Brefeld) Batko sobre ninfas do 5º ínstar de Empoasca kraemeri (Ross & Moore, 1957) a 20ºC e 100% de umidade relativa (RH). A germinação dos conídios primários dos isolados sobre o inseto iniciou-se dentro do período de duas horas após a inoculação. A produção total de tubos germinativos tendeu a se igualar a de capiloconídios sobre o corpo do hospedeiro, doze horas após a inoculação. A proporção de conídios secundários, dentro das formas geradas pelos conídios primários germinados sobre o inseto, não ultrapassou 25% em qualquer das regiões do corpo.
Insecta; cigarrinha verde; entomophthorales; controle biológico
The germination processes of ARS-1590, ARS-1261 and ARS-1229 isolates of Zoophthora radicans (Brefeld) Batko on fifth-instar Empoasca kraemeri (Ross & Moore, 1957) nymphs, at 20ºC and 100% relative humidity were investigated and compared. The primary conidia on the insect germinated within two hours after inoculation for all isolates. The total production of germ tubes tended to be equal to that of capilliconidium on the host body, 12 hours after inoculation. The secondary conidia proportion, among the forms produced by the primary conidia on the insect, did not exceed 25% in all body regions.
Insecta; leafhopper; entomophthorales; biological control
PROCESSOS DE GERMINAÇÃO DE ISOLADOS DO FUNGO Zoophthora radicans SOBRE Empoasca kraemeri
L.G. LEITE1; S.B. ALVES2; S.P. WRAIGHT3; S. GALAINI-WRAIGHT4; D.W. ROBERTS4
1Seção de Controle Biológico das Pragas-IB, C.P. 70, CEP: 13001-970 - Campinas, SP.
2Depto. de Entomologia-ESALQ/USP, C.P. 9, CEP: 13418-900 - Piracicaba, SP.
3USDA, ARS/US Plant, Soil & Nutritions Lab., Ithaca, New York, 14853, USA.
4Boyce Thompson Institute, Insect Pathology Resource Center, Tower Rd. Ithaca, New York, 14853, USA.
RESUMO: Foram analisados e comparados os processos de germinação dos isolados ARS-1590, ARS-1261 e ARS-1229 de Zoophthora radicans (Brefeld) Batko sobre ninfas do 5º ínstar de Empoasca kraemeri (Ross & Moore, 1957) a 20ºC e 100% de umidade relativa (RH). A germinação dos conídios primários dos isolados sobre o inseto iniciou-se dentro do período de duas horas após a inoculação. A produção total de tubos germinativos tendeu a se igualar a de capiloconídios sobre o corpo do hospedeiro, doze horas após a inoculação. A proporção de conídios secundários, dentro das formas geradas pelos conídios primários germinados sobre o inseto, não ultrapassou 25% em qualquer das regiões do corpo.
Descritores: Insecta, cigarrinha verde, entomophthorales, controle biológico
GERMINATION PROCESSES OF ISOLATES OF THE ENTOMOPHTHORALES FUNGUS Zoophthora radicans ON Empoasca kraemeri
ABSTRACT: The germination processes of ARS-1590, ARS-1261 and ARS-1229 isolates of Zoophthora radicans (Brefeld) Batko on fifth-instar Empoasca kraemeri (Ross & Moore, 1957) nymphs, at 20ºC and 100% relative humidity were investigated and compared. The primary conidia on the insect germinated within two hours after inoculation for all isolates. The total production of germ tubes tended to be equal to that of capilliconidium on the host body, 12 hours after inoculation. The secondary conidia proportion, among the forms produced by the primary conidia on the insect, did not exceed 25% in all body regions.
Key Words: Insecta, leafhopper, entomophthorales, biological control
INTRODUÇÃO
O fungo Zoophthora radicans Brefeld (Humber) (=Erynia radicans = Entomophthora sphaerosperma) é bem conhecido no mundo por ser entomopatógeno polífago (Ben Ze'ev et al., 1984), sendo uma espécie de Entomophthorales amplamente estudada (Galaini-Wraight et al., 1991). O fungo já foi citado sobre diversas espécies de insetos, agrupadas em um mínimo de quatro ordens (Glare et al., 1987), tendo também sido relatado como patógeno capaz de provocar altos níveis de infecção em populações de hospedeiros (Sawyer, 1929; Jaques & Patterson, 1962; Vandenberg & Soper, 1978). No Brasil, Z. radicans foi primeiramente citado em populações de Anticarsia gemmatalis Hubner em campo de soja (Hoffmann et al., 1979) e, posteriormente, em Empoasca kraemeri Hoss e More na cultura do caupi (Daoust et al., 1983).
A possibilidade de produção massal de micélio seco armazenável desse fungo permite considerá-lo como agente de controle biológico com potencial de uso para o combate de diversas pragas, dentre as quais a cigarrinha verde do feijão, Empoasca sp.. O fungo, na forma de partículas de micélio seco ou em estado natural no cadáver do inseto, quando em contato com o orvalho, entra em conidiogênese e ejeta os conídios nas proximidades. Os conídios, em função do substrato que atingem, podem germinar de três formas diferentes, dando origem a tubo germinativo, que penetra no hospedeiro, bem como capiloconídio ou conídio secundário, os quais podem resultar no processo de infecção ou formar conídios terciários. O capiloconídio liga-se ao conídio primário por um conidióforo longo e possui disseminação passiva, enquanto que o conídio secundário liga-se por um conidióforo curto, possuindo disseminação ativa.
Neste trabalho foram analisados e comparados os processos de germinação de três isolados de Z. radicans sobre E. kraemeri.
MATERIAL E MÉTODOS
Os estudos foram realizados no Laboratório de Patologia de Insetos do Centro Nacional de Pesquisa de Arroz e Feijão (CNPAF), da EMBRAPA, com três isolados de Z. radicans, no tocante à viabilidade dos conídios e seus processos de germinação sobre ninfas do 5º ínstar de E. kraemeri, obtidas de criação em laboratório. As observações foram feitas no dorso do inseto, separadamente, nas regiões da cabeça, tórax e abdome. Os conídios não germinados foram considerados inviáveis, enquanto que os germinados foram considerados viáveis e submetidos a avaliações referentes a formação de conídios secundários, capiloconídios e tubos germinativos.
Os isolados do fungo utilizados foram o ARS-1590 e o ARS-1229, originados de Empoasca sp., respectivamente dos Estados de Goiás e Santa Catarina, no Brasil, bem como o ARS-1261, originado de E. fabae do Estado de Wisconsin, nos Estados Unidos, os quais foram fornecidos pelo Laboratório de Patologia de Insetos do Instituto Boyce Thompson em Ithaca, EUA. Esses foram produzidos na superfície de meio de cultura líquido à base de dextrose (1%) e extrato de levedura (1%) e utilizados na forma de micélio fresco. Para a instalação do experimento, o fungo foi previamente filtrado, pulverizado com maltose a 10% e transferido para uma câmara úmida formada dentro de uma placa de Petri, contendo papel de filtro umedecido com a mesma solução. Em seguida, a placa de Petri contendo o micélio fresco do patógeno foi acondicionada dentro de geladeira e mantida por aproximadamente quinze horas, visando estimular a conidiogênese.
As inoculações do fungo foram efetuadas mediante a utilização de pequenas gaiolas. Cada gaiola era constituída de uma placa de plástico de 4,0cm de diâmetro e de fundo aberto, contendo apenas um segmento de 0,5cm de largura, ligando as duas extremidades laterais opostas. A placa foi colocada sobre a face inferior de uma folha de caupi, que por sua vez foi estendida com a face superior sobre uma chapa de acrílico (15,5 x 10cm). O conjunto foi fixado com um parafuso ligando o segmento do fundo da placa com a chapa de acrílico. Na parte de dentro da tampa da gaiola foi colocada uma camada de ágar-água com antibiótico. Para a inoculação dos insetos, pequenos círculos de micélio fresco foram distribuídos sobre a camada de ágar-água da tampa, e acondicionados dentro de uma câmara úmida (saco plástico contendo papel toalha umedecido), no interior de uma incubadora regulada a 20 ± 0,5ºC, onde foram mantidos por mais quatro horas, visando aumentar a conidiogenese do fungo.
Em seguida, as gaiolas e as superfícies expostas das folhas foram umedecidas com uma fina névoa de água destilada, para simular o orvalho natural. Os insetos foram anestesiados com CO2 e colocados na superfície úmida da folha em cada gaiola. As tampas das gaiolas, contendo o fungo em conidiogênese, foram colocadas sobre os insetos e mantidas por aproximadamente cinco minutos (período de duração da anestesia), expondo-os à "chuva" de conídios. Durante cada período de exposição, as tampas contendo o fungo eram constantemente giradas para assegurar uma distribuição uniforme dos conídios.
Após cada inoculação, a tampa com o fungo era removida e substituída por outra limpa. As gaiolas contendo os insetos inoculados foram, então, colocadas dentro de sacos plásticos contendo umidade saturada (produzida através de papel toalha umedecido), e mantidas no escuro a 20 ± 0,5ºC. Os estudos foram feitos sobre os insetos, após duas, seis e doze horas da inoculação. No primeiro horário, foram utilizados dez, doze e oito insetos para os isolados ARS-1590, ARS-1261 e ARS-1229, respectivamente, enquanto que, no segundo, foram utilizados 13, 16 e 10 insetos, respectivamente, e, no terceiro, 20, 22 e 20 insetos. Os insetos foram coletados de dentro das gaiolas de inoculação, sendo um deles utilizado para as observações mais imediatas e os demais acondicionados em geladeira, para que o fungo inoculado fosse inativado e pudesse em seguida ser observado.
As observações foram feitas com uma objetiva de 400 x em um microscópio óptico com contraste de fase e fluorescência, tendo sido utilizado um excitador Schott BG-12 (350-490nm), um refletor Dichroic(500mm) e uma barreira Schott 06 515 (525nm). Os insetos foram colocados sobre uma gota de Calcofluor White (Sigma) a 0,5% (W/V) depositada sobre uma lâmina e cobertos, sem serem pressionados, por uma lamínula de vidro.
RESULTADOS E DISCUSSÃO
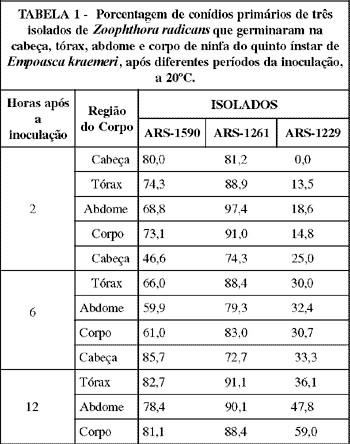
O isolado ARS-1261 apresentou uma viabilidade pouco maior que a do ARS-1590, e bastante superior em relação a do ARS-1229, tendo havido, respectivamente, 88,4, 81,1 e 59% de conídios primários germinados no corpo do inseto, doze horas após a inoculação (TABELA 1). A baixa viabilidade do ARS-1229 sobre os insetos pode limitar a eficiência deste isolado para o controle da praga. Smith & Grula (1981) constataram que uma fonte de carbono é necessária para a germinação de conídios de Beauveria bassiana (Bals.) Vuill.. Segundo Ferron (1983), conídios de fungos entomopatogênicos têm reserva de nitrogênio endógeno suficiente para a síntese de proteína e o processo de germinação. No entanto,necessitam de uma fonte de carbono para que este fenômeno ocorra. As principais fontes de carbono citadas pelo autor são ácidos nucléicos e alguns ácidos gordurosos. Considerando-se que a germinação de Z. radicans tem sido constatada em substrato inerte (Wraight (USDA), informação pessoal),é possível que fontes de carbono tenham pouca influência na germinação do fungo. A redução na porcentagem de germinação dos conídios inoculados,do abdome para a cabeça,em todos os tempos, para o isolado ARS-1229, pode ser devida a existência de algum composto químico inibidor da germinação deste isolado, presente na cutícula do hospedeiro em maior concentração nas camadas de maior espessura e esclerotizadas.
As porcentagens dos conídios germinados dos isolados, que produziram tubos germinativos, tubos capilares e conídios secundários, variaram conforme a região do corpo do hospedeiro (TABELA 2). O isolado ARS-1261 aumentou a produção de tubos germinativos da cabeça para o tórax e abdome em todos os períodos após a inoculação, visto que, na média dos tempos, esta forma foi gerada por 42,0; 54,4 e 66,5% dos conídios primários germinados nas respectivas regiões. Este comportamento, embora não tenha sido verificado para o isolado ARS-1590 e observado somente doze horas após a inoculação para o ARS-1229, pode ser característico do fungo, pois foi constatado também por Wraight et al. (1990). Segundo estes autores, a produção de tubos germinativos tende a aumentar da cabeça para o abdome devido ao fato dessa forma, produzindo apressório, ser favorecida nas regiões do hospedeiro com cutícula mais fina, como o abdome. Como conclusão, os autores relataram que um estimulante químico, presente em maior concentração no abdome, induz a produção de tubos germinativos. Van Roermund et al. (1984) mencionaram que a produção de tubos germinativos é governada também por estimulantes nutricionais, pois verificaram maior produção de tubos germinativos em substrato rico em nutriente do que em substrato composto de ágar-água.
O isolado ARS-1261 apresentou uma redução na produção de tubos capilares da cabeça do inseto para o tórax e abdome, visto que, na média dos tempos, esta forma foi gerada por 58,0; 45,6 e 33,5% dos conídios primários germinados nas respectivas regiões. Essa redução deveu-se ao aumento da produção de tubos germinativos, provocado pelo aumento de estímulos para a geração desta forma. Já o isolado ARS-1590, apresentou um sensível aumento na produção de tubos capilares da cabeça para o tórax. Sampedro et al. (1984) mencionaram que o modo de germinação do fungo Conidiobolus obscurus Brefeld (1884) é dependente do isolado, bem como das condições físicas e nutricionais do substrato. Para qualquer um dos isolados, os inóculos germinados sobre o inseto apresentaram, doze horas após a inoculação, uma produção total de tubos germinativos equilibrada com a de tubos capilares.
No que se refere aos conídios secundários, estes representaram baixas porcentagens dentro das formas geradas pelo fungo, tendo sido produzido, 12 horas após a inoculação, por 1,9, 2,2 e 1,9% dos conídios primários germinados do isolado ARS-1590, nas regiões da cabeça, tórax e abdome, respectivamente, e por 24,9 e 4,6% dos conídios primários germinados do isolado ARS-1229 na cabeça e abdome, respectivamente.
A germinação dos inóculos dos isolados ARS-1590 e ARS-1261 iniciou-se e praticamente estabilizou-se duas horas após a inoculação do fungo. Assim, pode-se afirmar que o isolado ARS-1229 foi o mais lento quanto a germinação dos conídios, visto que, as porcentagens de conídios germinados nas diferentes regiões do corpo do inseto foram baixas duas horas após a inoculação, aumentando nos períodos posteriores. Além disso, a produção de tubos capilares não ocorreu antes das duas horas. De modo geral, pode-se afirmar que a germinação dos conídios de Z. radicans ocorre num período extremamente curto após a inoculação, tendo por comparação os Hifomicetos B. bassiana e Nomuraea rileyi Farlow (Samson), cuja germinação de conídios ocorre em cerca de 12 e 18 horas, respectivamente (Alves, 1986), bem como Metarhizium anisopliae (Metsch.) Sorokin, que leva, comumente, cerca de 18 a 24 horas (Samuels et al., 1989). Segundo Steinhaus (1949), a germinação dos conídios de fungos Entomophthorales ocorre logo após a descarga dos mesmos.
CONCLUSÕES
1) A germinação dos conídios primários dos isolados sobre o inseto iniciou-se dentro do período de duas horas após a inoculação.
2) A produção total de tubos germinativos tendeu a se igualar a de capiloconídios sobre o corpo do hospedeiro, doze horas após a inoculação.
3) A proporção de conídios secundários, dentro das formas geradas pelos conídios primários germinados sobre o inseto, não ultrapassou 25% em qualquer das regiões do corpo.
AGRADECIMENTOS
Ao convênio EMBRAPA/CNPAF-BOYCE THOMPSON INSTITUTE pelo apoio para realização dessa pesquisa.
Recebido para publicação em 13.11.95
Aceito para publicação em 09.02.96
- ALVES, S.B. Controle microbiano de insetos Săo Paulo: Manole,1986. p.29-60: Epizootiologia.
- BEN-ZE'EV, I.; KENNETH, R.G.; SOPER, R.S. The Entomophthorales of Israel and their arthropod hosts: seasonal occurrence. Phytoparasitica, v.12, n.3/4, p.167-176, 1984.
- DAOUST, R.A.; ROBERTS, D.W.; SOPER, R. Fungal diseases of cowpea pests in North, Northeast and Central West Brazil. In: ANNUAL MEETING OF THE SOCIETY FOR INVERTEBRATE PATHOLOGY, 16., New York, 1983. p.42.
- FERRON, P. Fungal control. In: KERKHUT, G.A.; GILBERT, L.I., eds. Comprehensive insect physiology, biochemistry and pharmacology, Oxford: Pergamon, 1983. p.314-346.
- GALAINI-WRAIGHT, S.; WRAIGHT, S.P.; CARRUTHERS, R.I.; MAGALHĂES, B.P.; ROBERTS, D.W. Description of a Zoophthora radicans (Zigomycetes: Entomophthoraceae) epizootic in a population of Empoasca kraemeri (Homoptera: Cicadellidae) on beans in Central Brasil. Journal of Invertebrate Pathology, v.58, p.311-326, 1991.
- GLARE, T.R.; MILNER, R.J.; CHILVERS, G.A.; MAHON, R.J.; BROWN, W.V. Taxonomic implications of interspecific variation amongst isolates of the aphid-pathogenic fungi Zoophthora radicans Brefeld and Z. phalloides Batko (Zygomycetes: Entomophthoraceae). Australian Journal of Botany, v.35, p.49-67, 1987.
- HOFFMANN, C.B.; NEWMAN, G.G.; FOERSTER, L.A. Incidęncia estacional de doenças e parasitas em populaçőes naturais de Anticarsia gemmatalis Hubner, 1818 e Plusia spp. em soja. Anais da Sociedade Entomologica do Brasil, v.8, n.1, p.115-124, 1979.
- JAQUES, R.P.; PATTERSON, N.A. Control of the apple sucker, Psylla mali Schmidb, by the fungus Entomophthora sphaerosperma (Fresenius). Canadian Entomologist, v.94, p.818-825, 1962.
- SAMPEDRO, M.L.; LATGÉ, J.P.; UZIEL, A. Agressiveté de Conidiobolus obscurus vis-ŕ-vis du puceron du pois Acyrthosiphon pisum. Mycophathologia, v.86, p.3-19, 1984.
- SAMUELS, K.D.Z.; HEALE, J.B.; LLEWELLYN, M. Characteristics relating to the pathogenicity of Metarhizium anisopliae toward Nilaparvata lugens Journal of Invertebrate Pathology, v.53, p.25-31, 1989.
- SAWYER, W.H. Observations on some entomogenous members of the Entomophthoraceae in artificial culture. American Journal of Botany, v.16, p.27-121, 1929.
- SMITH, R.J.; GRULA, E.A. Nutritional requirements for conidial germination and hyphal growth of Beauveria bassiana. Journal of Invertebrate Pathology, v.37, p.222-230, 1981.
- STEINHAUS, E.A. Principles of insect pathology, New York: McGraw-Hill, 1949. 757p.
- VANDERBERG, J.D.; SOPER, R.S. Prevalence of Entomophthorales mycoses in populations of spruce budworm, Choristoneura fumiferana Environmental Entomology, v.7, p.847-853, 1978.
- VAN ROERMUND, H.J.W.; PERRY, D.F.; TYRRELL, D. Influence of temperature, light, nutrients and pH in determination of the mode of conidial germination in Zoophthora radicans. Transactions of the British mycological Society, v.82, n.1, p.31-38, 1984.
- WRAIGHT, S.P.; BUTT, T.M.; GALAINI-WRAIGHT, S.; ALEE, L.L.; SOPER, R.S.; ROBERTS, D.W. Germination and infection processes of the Entomophthoralean fungus Erynia radicans on the potato leafhopper, Empoasca fabae. Journal of Invertebrate Pathology, v.56, p.157-174, 1990.
Datas de Publicação
-
Publicação nesta coleção
26 Fev 1999 -
Data do Fascículo
Jan 1996
Histórico
-
Aceito
09 Fev 1996 -
Recebido
13 Nov 1995