Resumos
O presente trabalho teve por objetivo estudar as interferências da disponibilidade hídrica no processo de germinação das sementes do feijoeiro. Para tanto, substratos com vários potenciais hídricos foram obtidos através do uso de soluções com polietileno glicol (PEG 6000) para avaliar a marcha de absorção de água pelas sementes e o desenvolvimento inicial das plântulas. Foi concluído que o teor mínimo de água nas sementes, exigido para a emergência da raiz primária indicadora do início da germinação visível situa-se no intervalo de 48 a 50%; o decréscimo do potencial hídrico externo causa maiores prejuízos ao crescimento das plântulas do que ao processo de emissão das raízes primárias.
Phaseolus vulgaris L.; germinação; polietileno glicol; absorção; plântulas; raiz; feijão
The present work has the objective of studying the interferences of water availability on the germination process of dry bean seeds. For this purpose substrates with several hydric potentials were obtained using aqueous solutions of polyethylene glycol (PEG 6000) to evaluate the water uptake by seeds and the initial development of the seedlings. It was concluded that the minimum seed moisture content required by seeds for the radicle protrusion, which is the visible indicator of the onset of germination, is between 48 and 50%. The decrease of the external hydric potential causes greater reduction in the initial development of the seedlings than on the normal process of radicle protrusion.
Phaseolus vulgaris L.; germination; polyethylene glycol; moisture; seedlings; root; bean
CONTROLE DA HIDRATAÇÃO DAS SEMENTES E DESENVOLVIMENTO INICIAL DE PLÂNTULAS DO FEIJOEIRO
P.S. SHIOGA1; W.R. da SILVA2
2Instituto Agronômico do Paraná, C.P. 1331, CEP: 86001-970 - Londrina, PR.
3Depto. de Agricultura-ESALQ/USP, C.P. 9, CEP: 13418-900 - Piracicaba, SP.
RESUMO: O presente trabalho teve por objetivo estudar as interferências da disponibilidade hídrica no processo de germinação das sementes do feijoeiro. Para tanto, substratos com vários potenciais hídricos foram obtidos através do uso de soluções com polietileno glicol (PEG 6000) para avaliar a marcha de absorção de água pelas sementes e o desenvolvimento inicial das plântulas. Foi concluído que o teor mínimo de água nas sementes, exigido para a emergência da raiz primária indicadora do início da germinação visível situa-se no intervalo de 48 a 50%; o decréscimo do potencial hídrico externo causa maiores prejuízos ao crescimento das plântulas do que ao processo de emissão das raízes primárias.
Descritores:Phaseolus vulgaris L., germinação, polietileno glicol, absorção, plântulas, raiz, feijão
SEED HYDRATION CONTROL AND INITIAL DEVELOPMENT OF COMMON BEAN (Phaseolus vulgaris L.) SEEDLINGS
ABSTRACT: The present work has the objective of studying the interferences of water availability on the germination process of dry bean seeds. For this purpose substrates with several hydric potentials were obtained using aqueous solutions of polyethylene glycol (PEG 6000) to evaluate the water uptake by seeds and the initial development of the seedlings. It was concluded that the minimum seed moisture content required by seeds for the radicle protrusion, which is the visible indicator of the onset of germination, is between 48 and 50%. The decrease of the external hydric potential causes greater reduction in the initial development of the seedlings than on the normal process of radicle protrusion.
Key Words:Phaseolus vulgaris L., germination, polyethylene glycol, moisture, seedlings, root, bean
INTRODUÇÃO
Durante o processo de hidratação em condições de campo, a absorção de água pelas sementes sofre a interferência de fluxos de água, tais como precipitação, evapotranspiração (incluindo a própria semente como superfície evaporante), escoamento superficial, percolação e ascensão de água do lençol freático (Labouriau,1983). Além disso, estão diretamente envolvidos, na disponibilidade e velocidade do fluxo de água para as sementes, o potencial mátrico do solo ou substrato, a textura do solo, o gradiente de potencial de água entre o solo e a semente, a condutividade hidráulica do solo, o tamanho e a estrutura do tegumento das sementes e a temperatura (Collis-George & Hector, 1966; Carvalho & Nakagawa, 1983; Bewley & Black, 1985 e Peske & Delouche 1985). Enquanto as sementes absorvem água, o seu potencial de água se eleva, diminuindo o gradiente existente com o do solo até a igualação entre ambos (Shaykewich & Willians, 1971; Young et al., 1983).
À medida em que decresce o potencial hídrico externamente, a velocidade de absorção pelas sementes diminui (Parmar & Moore, 1968 ; Bradford, 1986), aumentando o período necessário para que seja atingindo o teor mínimo de água exigido para a emergência da raiz primária ou do epicótilo (Koller & Hadas, 1982; Prokof'Ev et al., 1983; Bradford, 1986).
O processo de divisão celular parece não estar envolvido diretamente na expansão inicial da raiz primária (Bewley & Black, 1985). Observações, efetuadas em ambientes de estresse hídrico, permitem concluir que eventos, como alongamento celular e subseqüente crescimento de plantas, são governados de maneira independente (Heydecker, 1977, Ross & Hegarty, 1979; Bewley & Black, 1985; Van de Venter, 1988). Espécies que germinam adequadamente sob baixos potenciais hídricos não são, necessariamente, as que sobreviverão sob severas condições de estresse hídrico (Mc Ginnies, 1960).
A disponibilidade de água para as sementes é diretamente relacionada com a germinação e emergência (Collis-George & Hector, 1966; Peske & Delouche, 1985), uma vez que a hidratação é fator limitante dos processos metabólicos (Kramer et al., 1966). A redução progressiva, do potencial hídrico, gera tendências de redução na velocidade e na porcentagem de germinação (Machado et al., 1976; Akalehiywot & Bewley, 1977; Young et al., 1983; Bewley & Black, 1985; Peske & Delouche, 1985; Bradford, 1986), assim como no desenvolvimento inicial de plântulas (Prisco & O'Leary, 1970; Magalhães & Carelli, 1972; Machado et al., 1976; Sá, 1987; Silva & Marcos Filho, 1990).
Dessa forma, considerando a hidratação como uma etapa limitante de vários outros processos fisiológicos que ocorrem na semente germinante, o presente estudo objetivou averiguar as interferências da disponibilidade hídrica no desenvolvimento inicial das plântulas do feijoeiro.
MATERIAL E MÉTODOS
O trabalho foi conduzido no Laboratório de Análise de Sementes do Departamento de Agricultura da Escola Superior de Agricultura "Luiz de Queiroz" (USP), Piracicaba/SP. Para compor o experimento, empregaram-se dois lotes (A e B) de sementes de feijão preto do cultivar IAPAR 20.
As sementes foram classificadas por espessura pela passagem em peneiras de crivos oblongos superpostas, em ordem decrescente (13/64 x 3/4, 12/64 x 3/4, 11/64 x 3/4 de polegada), para aproveitamento exclusivo daquelas retidas na peneira 12/64 x 3/4. Nessa condição, com equivalência no tamanho, as sementes sofreram homogeneização e permaneceram em condições de ambiente não controlado para uniformização do teor de água entre os lotes.
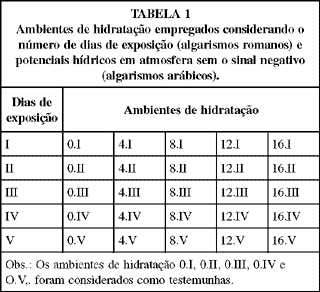
Para a obtenção dos tratamentos, além do controle (substrato saturado com água destilada) denominado 0 atm, foram empregadas soluções de polietileno glicol (PEG 6000) de modo a serem desenvolvidos potenciais hídricos de -4, -8, -12 e -16 atm (1 MPa = 10 bar = 9,87 atm). Todas as soluções, bem como o controle utilizado, foram preparados com água destilada. As quantidades de soluto para a obtenção dos potenciais hídricos tiveram como base de cálculo a fórmula desenvolvida por Michel & Kaufmann (1973). A dissolução do soluto foi efetuada em agitador mecânico com um dia de antecedência à fase experimental. A água destilada (0 atm) e as soluções foram aplicadas em papel toalha, na razão de 2,5:1,0 em peso, constituindo o substrato para as sementes. A TABELA 1 indica os tratamentos considerados.
A distribuição de 10 sementes por repetição sobre duas folhas de papel, cobertas por uma terceira, obedeceu a um espaçamento equidistante entre as sementes. Estas, por sua vez, foram colocadas sobre um traço picotado alinhado no sentido longitudinal, localizado no terço superior do substrato, com o hilo posicionado de tal forma que ficasse para baixo quando as sementes fossem levadas ao germinador. Posteriormente, os papéis eram enrolados, embalados em sacos plásticos e levados para germinador regulado à temperatura constante de 25ºC, em ausência de luz.
As determinações laboratoriais, realizadas em quatro repetições por tratamento, foram as seguintes:
a) Determinação do grau de umidade: baseou-se nos procedimentos estabelecidos nas Regras para Análise de Sementes (Brasil, 1980) para a determinação do teor de umidade pelo método da estufa (105 3ºC/24 horas). Os resultados foram expressos em porcentagem considerando as sementes e as plântulas presentes.
b) Acréscimo diário no teor de água: foi obtido através da subtração do valor do teor de água (%) encontrado no dia anterior (N-1) daquele presente no dia (N) da avaliação. O resultado do primeiro dia foi calculado através da subtração do valor obtido, no dia da avaliação, do encontrado anteriormente à semeadura.
c) Número de raízes emersas: representado pela contagem diária dos indivíduos que, em qualquer estádio de desenvolvimento, apresentavam o tegumento rompido pela emissão da raiz primária.
d) Peso da matéria fresca das plântulas: obtido através de avaliações diárias realizadas após a eliminação dos tegumentos e cotilédones. Os resultados foram expressos em miligramas.
e) Comprimento das plântulas: determinado por medições diárias das plântulas. Os resultados foram expressos em milímetros.
Uma vez obtidos, os dados passaram por análise de variância realizada, de modo independente, para cada dia e lote. As médias foram comparadas pelo teste de Tukey (5% de probabilidade).
RESULTADOS E DISCUSSÃO
Os dados obtidos, através das determinações diárias do teor de água e do cálculo de seu acréscimo nas sementes e plântulas, estão apresentados nas TABELAS 2 e 3.
O controle da hidratação, exercido pelos potenciais hídricos, foi eficiente na sua ação. Os dados experimentais de teor de água, contidos na TABELA 2, revelam que houve reduções à medida em que decrescia o potencial hídrico, dentro de cada dia, para ambos os lotes. Paralelamente, foi possível observar tendência dos teores de água, nos potenciais de -12 e -16 atm, se igualarem ao longo dos cinco dias de exposição, nos dois lotes avaliados. Em termos gerais, houve elevação nos valores do teor de água, com o passar do tempo, em todos os ambientes de hidratação; contudo, esta tendência se atenuou com a redução dos potenciais hídricos de tal forma que, a partir de -8 atm, os dados apresentaram variações diárias reduzidas e direcionadas à estabilização. Estes fatos foram observados por Prisco & O'Leary (1970) quando obtiveram valores crescentes de absorção de água pelas sementes, no decorrer do tempo de exposição, até o potencial de -4 atm e estabilização nos potenciais iguais ou menores que -8 atm.
Considerando os valores da TABELA 3, constata-se um destacado acréscimo no teor de água no primeiro dia em ambos os lotes, em função do gradiente de potencial hídrico existente entre as sementes e os substratos; a absorção de água foi realizada com grande velocidade e alta dependência das forças mátricas responsáveis pela primeira fase de absorção de água pelas sementes (Mayer & Poljakoff-Mayber, 1963, Prisco & O'Leary, 1970; Paulilo, 1980; Bewley & Black, 1985; Salisbury & Ross, 1985). A observação dos valores de acréscimo, no primeiro dia, permite verificar superioridade nos ambientes com maiores potenciais hídricos, que se atenuou com o andamento experimental. Avaliando os resultados, segundo a evolução do período de exposição aos ambientes da hidratação, verifica-se que, apesar da diferenciação presente no 1º dia, os acréscimos diários tenderam a se igualar; esta ocorrência foi mais nítida no lote B que, no 5 dia, não indicou variações estatisticamente identificáveis entre os tratamentos.
Dessa maneira, uma análise conjunta das TABELAS 2 e 3 permite constatar que, apesar da testemunha haver mostrado, com o passar do tempo, superioridade crescente quanto ao teor de água, a sua taxa de absorção, medida através do acréscimo diário, apresentou tendência ao nivelamento com os demais tratamentos. Examinando as duas TABELAS, os valores dos dois lotes nos potenciais mais negativos(-8 -12 e -16 atm) a partir do 3º dia, observa-se uma acomodação dos resultados, possivelmente em razão dos potenciais hídricos, das sementes ou plântulas, se aproximarem do equilíbrio com os potenciais hídricos das soluções. Ainda, comparando essas TABELAS, em cada ambiente de hidratação individualmente, observa-se semelhança nos dados obtidos nos dois lotes, sugerindo não haver diferença marcante na hidratação de ambos.
Os dados, relativos ao número de raízes emersas (TABELA 4), indicaram comportamentos similares nos dois lotes. No 1º dia, a testemunha apresentou, praticamente, todas as sementes com as raízes emersas, enquanto outros tratamentos não mostravam nenhuma emissão. No segundo dia, aproximadamente 50% do total de sementes estavam com raízes emitidas no potencial de -4 atm, não havendo a partir do terceiro dia diferenças estatísticas entre a testemunha e este potencial. Excetuando-se as indicações de emissão da raiz primária nos ambientes 8.IV e 8.V no lote B, observa-se que, nos potenciais iguais ou inferiores a -8 atm, não houve desenvolvimento das estruturas embrionárias capaz de provocar a ruptura do tegumento. À medida em que decresceu o potencial hídrico do substrato, o teor de água se reduziu (TABELA 2), retardando o início da emissão da raiz primária. Estes fatos foram observados entre a testemunha, cuja emissão ocorreu no 1º dia, e o potencial de -4 atm com emissão iniciada no 2º dia; o mesmo fato aconteceu entre os potenciais de -4 e de -8 atm, cuja emissão teve início no 4º dia no lote B. Esta observação concorda com dados de Bradford (1986) ao concluir que, quando o potencial externo é reduzido, o teor de água das sementes é inferior ao presente em potenciais maiores e o início da germinação é retardado. Considerando que a emergência da raiz primária depende da obtenção de um teor mínimo de água pelas sementes (Koller & Hadas, 1982; Prokof'Ev et al., 1983; Bradford, 1986), a análise conjunta das TABELAS 2 e 4 dos ambientes de hidratação 4.II, nos dois lotes, e 8.IV e 8.V. no lote B, oferece indicações de que valores, entre 48 e 50% no teor de água das sementes, são suficientes para que tenha início o crescimento visível do eixo embrionário. No ambiente de hidratação 8.V, apesar do teor de água ter alcançado 48,5% sem o surgimento de sinal visível de crescimento, a ausência de emissão radicular pode estar relacionada com a qualidade fisiológica inferior do lote A e, em função disso, ser justificado um desenvolvimento mais lento.
A análise dos pesos da matéria fresca e dos comprimentos das plântulas (TABELAS 5 e 6) indicou comportamentos semelhantes dos tratamentos nas duas determinações; a única exceção, situada no 4º dia, ocorreu quando o teste de comprimento das plântulas detectou superioridade de -4 atm sobre os potenciais inferiores, em ambos os lotes, contrastando com a ausência de diferenças verificada no peso da matéria fresca. A testemunha apresentou superioridade em relação aos demais tratamentos em todos os dias avaliados. Prisco & O'Leary (1970) e Machado et al. (1976) afirmaram que baixos potenciais de água reduziram o comprimento do hipocótilo e da raiz primária das plântulas de feijão submetidas a potenciais hídricos inferiores a -3,5 atm.
Confrontando a TABELA 4 com as TABELAS 5 e 6, para ambos os lotes, particularmente no potencial de -4 atm, observa-se que a partir do 2º dia a quase totalidade das sementes havia emitido as raízes primárias igualando-se estatisticamente à testemunha; o mesmo não aconteceu com o desenvolvimento inicial das plântulas, em que ocorreu inferioridade significativa do potencial -4 atm, em relação à testemunha, em todos os dias. Esses fatos permitem concluir que o desenvolvimento da plântula sofreu mais as restrições hídricas estabelecidas do que o antecedente processo de emissão da raiz primária. Van de Venter (1988) afirmou que sementes de milho, em condições hídricas adversas, são mais negativamente afetadas no crescimento da parte aérea do que na porcentagem de germinação. Essa observação foi efetuada por Prisco & O'Leary (1970) e confirmada por Eira (1988) ao verificarem que as sementes, apesar de emitirem raiz primária, originaram plântulas anormais em situação de déficit hídrico.
CONCLUSÕES
- O teor mínimo de água das sementes de feijão, exigido para a emergência da raiz primária indicadora do início da germinação visível, situa-se no intervalo entre 48 e 50%.
- O decréscimo do potencial hídrico externo causa maiores prejuízos ao crescimento das plântulas do que ao processo de emissão das raízes primárias.
Recebido para publicação em 20.02.97
Aceito para publicação em 22.10.97
- AKALEHIYWOT, T.; BEWLEY, J.D. Promotion and synchronization of cereal grain germination by osmotic pretreatment with polyetylene glycol. Journal of Agricultural Science, v.89, p.503-506, 1977.
- BEWLEY, J.D.; BLACK, M. Seeds: physiology of development and germination. New York: Plenum Press, 1985. 367p.
- BRADFORD, K.J. Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. HortScience, v.21, n.5, p.1105-1112, 1986.
- BRASIL. Ministério da Agricultura. Regras para análise de sementes. Brasília, 1980. 188p.
- CARVALHO, N.M.; NAKAGAWA, J. Sementes: cięncia, tecnologia e produçăo. 2.ed. Campinas: Fundaçăo Cargill, 1983. 429p.
- COLLIS-GEORGE, N.; HECTOR, J.B. Germination of seeds as influenced by matric potential and by area of contact between seed and soil water. Australian Journal of Soil Research, v.4, p.145-165, 1966.
- EIRA, M.T.S. Condicionamento osmótico de sementes de alface (Lactuca sativa L.): efeitos sobre a germinaçăo e desempenho sob estresses hídrico, salino e térmico. Piracicaba, 1988. 90p. Dissertaçăo (Mestrado) - Escola Superior de Agricultura "Luiz de Queiroz", Universidade de Săo Paulo.
- HEYDECKER, W. Stress and seed germination. In: KHAN., A.A. (Ed.). The physiology and biochemistry of seed dormancy and germination. Amsterdan: Elsevier; North Holland, 1977. p.240-282.
- KOLLER, D.; HADAS, A. Water relations in the germination of seeds. In: LANGE, D.L.: NOBEL, P.S.; OSMOND, C.B.; ZIEGLER, H. (Ed.). Encyclopedia of plant physiology Berlin: Springer-Verlag, 1982. p.401-431. (New Series, 12b).
- KRAMER, P.J.; KNIPLING, E.B.; MILLER, L.N. Terminology of cell-water relations. Science, v.155, p.889-890, 1966.
- LABOURIAU, L.G. A germinaçăo das sementes. Washington: OEA, 1983. 174p. (OEA. Coleçăo de Monografias Científicas Biológicas, 24).
- Mc GINNIES, W.J. Effects of moisture stress and temperature on germination of six range grasses. Agronomy Journal, v.52, p.159-163, 1960.
- MACHADO, R.C.R. ; RENA, A.B.; VIEIRA, C. Efeito da desidrataçăo osmótica na germinaçăo de sementes de vinte cultivares de feijăo (Phaseolus vulgaris). Revista Ceres, v.23, n.128, p.310-320, 1976.
- MAGALHĂES, A.C.; CARELLI, M.L. Germinaçăo de sementes de feijăo (Phaseolus vulgaris L.) sob condiçőes variadas de pressăo osmótica. Bragantia, v.31, p.19-26, 1972.
- MAYER, A.M.; POLJAKOFF-MAYBER, A. The germination of seeds London: Pergamon Press, 1963. 236p.
- MICHEL, B.E.; KAUFMANN, M.R. The osmotic potencial of polytylene glycol 6000. Plant Physiology, v.51, p.914-916, 1973.
- PARMAR, M.T.; MOORE, R.P. Carbowax 6000, Mannitol and Sodium Chloride for simulating drought conditions in germination studies of corn (Zea mays L.) of strong and weak vigour. Agronomy Journal, v.60, p.192-195, 1968.
- PAULILO, M.T.S. Efeito de ácido giberélico e 6-benziladenina na utilizaçăo das reservas da semente de Phaseolus vulgaris L. cv. Carioca. Campinas, 1980. 105p. Dissertaçăo (Mestrado) Universidade Estadual de Campinas.
- PESKE, S.T.; DELOUCHE, J.C. Semeadura de soja em condiçőes de baixa umidade do solo. Pesquisa Agropecuária Brasileira, v.20, n.1, p.69-85, 1985.
- PRISCO, J.T.; O'LEARY, J.W. Osmotic and "toxic" effects of salinity on germination of Phaseolus vulgaris L. seeds. Turrialba, v.20, n.2, p.177-184, 1970.
- PROKOFEV, A.A.; OBRUCHEVA, V.; KAVALDO, L.S.; KULIEVA, L.K.; KOZHEMAKINA, I.S. Level of seed water content critical for the outset of germination. Soviet Plant Physiology,v.30, p.144-148, 1983.
- ROSS, H.A.; HEGARTY, T.W. Sensitivity of seed germination and seedling radicle growth to moisture stress in some vegetable crop species. Annals of Botany, v.43, p.241-243, 1979.
- SÁ, M.E. Relaçőes entre qualidade fisiológica, disponibilidade hidrica e desempenho de sementes de soja (Glycine max (L.)) Merril. Piracicaba, 1987. 147p. Dissertaçăo (Mestrado) - Escola Superior de Agricultura "Luiz de Queiroz", Universidade de Săo Paulo.
- SALISBURY, F.B.; ROSS, C.W. Plant physiology 3.ed. Belmonte: Wadsworth, 1985. 540p.
- SHAYKEWICH, C.F.; WILLIANS, J. Resistence to water absorption in germination rap seed (Brassica napus L.). Journal of Experimental Botany, v.70, p.19-24, 1971.
- SILVA, W.R.; MARCOS FILHO, J., Avaliaçăo da embebiçăo e do desenvolvimento inicial das estruturas embrionárias de sementes de milho submetidas a diferentes potenciais hídricos. Scientia Agricola, v.47, p.335-359, 1990.
- VAN DE VENTER, H.A. Relative response of maize (Zea mays L.) seed lots to different stress conditions. Seed Science and Technology, v.16, p.19-28, 1988.
- YOUNG, J.A.; EVANS, R.A.; ROUNDY, B.; GREG, C. Moisture stress and seed germination. Washington: USDA, 1983. 40p. (Agricultural Research Service, 36).
Datas de Publicação
-
Publicação nesta coleção
12 Nov 1998 -
Data do Fascículo
Jan 1998
Histórico
-
Aceito
22 Out 1997 -
Recebido
20 Fev 1997