Abstracts
The cashew crop (Anacardium occiedentale L.) is of great economic and social importance for Northeast Brazil, a region usually affected by water and soil salinity. The present study was conducted in a greenhouse to evaluate the effects of four salinity levels established through electrical conductivity of irrigation water (ECw: 0.7, 1.4, 2.1 and 2.8 dS m-1, at 25ºC), on growth and physiological indexes of five rootstocks of dwarf-precocious cashew varieties CCP06, CCP09, CCP1001, EMBRAPA50, and EMBRAPA51. Plant height, leaf area, dry weight of root, shoot and total; water content of leaves, root/shoot ratio, leaf area ratio, absolute and relative growth rates and rate of net assimilation were evaluated. The majority of the evaluated variables were found to be affected by ECw and the effects varied among clones; however, no significant interactive effects were observed for factors. The value of ECw = 1.39 dS m-1 was considered as a threshold tolerance for the precocious cashew rootstocks used in this study. The dwarf-precocious cashew is moderately sensible to soil salinity during the formation phase of rootstock. Clones EMBRAPA51 and EMBRAPA50 presented, respectively, the least and the best development indexes.
Anacardium occidentale; electrical conductivity; clones; saline stress
A cultura de caju (Anacardium occiedentale L.) é de grande importância, econômica e social para o Nordeste brasileiro, região normalmente sujeita a problemas de salinidade da água e do solo. Este estudo, realizado em casa de vegetação, objetivou avaliar efeitos de quatro níveis de condutividade elétrica da água de irrigação (CEa: 0,7, 1,4, 2,1 e 2,8 dS m-1, a 25 ºC), sobre índices fisiológicos e de crescimento de cinco porta-enxertos de cajueiro anão-precoce: CCP06, CCP09, CCP1001, EMBRAPA50 e EMBRAPA51. Foram avaliados altura de planta, área foliar, fitomassa de raízes, da parte aérea e total, teor de água das folhas, relação raiz/parte aérea, razão de área foliar, taxas de crescimento absoluto, relativo e de assimilação líquida. A maioria das variáveis estudadas foi afetada pela salinidade da água de irrigação e variou entre clones, mas sem, haver efeito interativo desses fatores. O valor de CEa = 1,39 dS m-1 foi considerado como limite de tolerância à salinidade para o crescimento dos porta-enxertos utilizados neste estudo. O cajueiro anão-precoce é moderadamente sensível à salinidade do solo na fase de formação de porta-enxertos. Os clones EMBRAPA51 e EMBRAPA50 apresentaram, respectivamente, os piores e os melhores valores para os índices avaliados.
Anacardium occidentale; condutividade elétrica; clones; estresse salino
CROP SCIENCE
Salt tolerance of precocious-dwarf cashew rootstocks physiological and growth indexes
Tolerância de porta-enxertos de cajueiro anão-precoce à salinidade índices fisiológicos e de crescimento
Paulo Torres CarneiroI; Pedro Dantas Fernandes* * Corresponding author < pdantas@deag.ufcg.edu.br> ; Hans Raj Gheyi; Frederico Antônio Loureiro Soares; Sergio Batista Assis Viana
IUFCG/CCT - Depto. de Engenharia Agrícola, CP 10.087, 58109-970 - Campina Grande, PB - Brasil
ABSTRACT
The cashew crop (Anacardium occiedentale L.) is of great economic and social importance for Northeast Brazil, a region usually affected by water and soil salinity. The present study was conducted in a greenhouse to evaluate the effects of four salinity levels established through electrical conductivity of irrigation water (ECw: 0.7, 1.4, 2.1 and 2.8 dS m-1, at 25oC), on growth and physiological indexes of five rootstocks of dwarf-precocious cashew varieties CCP06, CCP09, CCP1001, EMBRAPA50, and EMBRAPA51. Plant height, leaf area, dry weight of root, shoot and total; water content of leaves, root/shoot ratio, leaf area ratio, absolute and relative growth rates and rate of net assimilation were evaluated. The majority of the evaluated variables were found to be affected by ECw and the effects varied among clones; however, no significant interactive effects were observed for factors. The value of ECw = 1.39 dS m-1 was considered as a threshold tolerance for the precocious cashew rootstocks used in this study. The dwarf-precocious cashew is moderately sensible to soil salinity during the formation phase of rootstock. Clones EMBRAPA51 and EMBRAPA50 presented, respectively, the least and the best development indexes.
Key words:Anacardium occidentale, electrical conductivity, clones, saline stress
RESUMO
A cultura de caju (Anacardium occiedentale L.) é de grande importância, econômica e social para o Nordeste brasileiro, região normalmente sujeita a problemas de salinidade da água e do solo. Este estudo, realizado em casa de vegetação, objetivou avaliar efeitos de quatro níveis de condutividade elétrica da água de irrigação (CEa: 0,7, 1,4, 2,1 e 2,8 dS m-1, a 25 oC), sobre índices fisiológicos e de crescimento de cinco porta-enxertos de cajueiro anão-precoce: CCP06, CCP09, CCP1001, EMBRAPA50 e EMBRAPA51. Foram avaliados altura de planta, área foliar, fitomassa de raízes, da parte aérea e total, teor de água das folhas, relação raiz/parte aérea, razão de área foliar, taxas de crescimento absoluto, relativo e de assimilação líquida. A maioria das variáveis estudadas foi afetada pela salinidade da água de irrigação e variou entre clones, mas sem, haver efeito interativo desses fatores. O valor de CEa = 1,39 dS m-1 foi considerado como limite de tolerância à salinidade para o crescimento dos porta-enxertos utilizados neste estudo. O cajueiro anão-precoce é moderadamente sensível à salinidade do solo na fase de formação de porta-enxertos. Os clones EMBRAPA51 e EMBRAPA50 apresentaram, respectivamente, os piores e os melhores valores para os índices avaliados.
Palavras-chave:Anacardium occidentale, condutividade elétrica, clones, estresse salino
INTRODUCTION
In Brazil, cashew (Anacardium occidentale L.) cultivation is concentrated in the Northeast region, mainly in the States of Ceará, Rio Grande do Norte and Piauí, which produce 126 thousand tons of cashew nut, of which approximately 28 thousand tons are exported, resulting in an annual income of about US$135 million in foreign exchange (FNP Consultoria e Comércio, 1998). The cultivation is characterized as activity of great socio-economical importance for the region, absorbing labour and generating foreign exchange for the country. Besides cashew nut, the pseudo-carp obtained from cashew has also great potential use in the most different forms and it may even become the main product of cashew exploration (Ramos et al., 1996).
Based on the false premise that the plant could be cultivated under conditions of extreme water adversity, the majority of the cashew orchards are non-irrigated, with non-grafted seedlings of common cashew trees, which results in low productivity. The cultivation of dwarf-precocious cashew permits the adoption of dense planting systems with improved clones, assuring productivity increments up to 942% of nuts and the full use of the pseudo-carp (Ramos et al., 1996). In this context, the perspectives of irrigated cashew cropping are increasing, aiming at higher productivity, reduced exploration risk, extension of the harvest period, and improvement in nut and pseudo-carp qualities (Oliveira et al., 1998). However, improper use of water in irrigation projects in the Northeast region of Brazil has promoted soil salinization (Audry & Suassuna, 1995).
The accumulation of soluble salts in the root zone reduces growth and development of the plants due to the decrease of the osmotic potential of the soil solution, resulting in water stress, and also because of toxicity problems and nutritional disorders (Fageria, 1989; Shannon, 1997). Saline stress, represents one of the most serious factors that limit the growth and yield of the crops, inducing morphological, structural and metabolical modifications in higher plants (Izzo et al., 1991).
In spite of the recognized socio-economic importance of this crop for the Northeast region, and the magnitude of the salinity problems, a few research projects have been conducted to investigate the effects of the irrigation with water of different qualities on cashew cultivation. In earlier studies, the water used in irrigation was either prepared with NaCl plus CaCl2 (Meireles, 1999) or just with NaCl (Ferreira et al., 2000; Viégas et al., 2001), without any information on the combined presence of the three main cations (Na, Ca and Mg), which are predominant in the irrigation waters used in the Northeast (Medeiros, 1992). In this study, the salinity tolerance of five clones of dwarf-precocious cashews was evaluated during the rootstock formation stage through physiological and growth indexes.
MATERIAL AND METHODS
The experiment was carried out in Campina Grande, PB, Brazil, (7º15'18" S, 35º52'28" W; altitude 550 m), from November 1999 to March 2000. The soil is an Alfisol (sandy-loam, pH 4.54, base saturation (BS) 33.7%, cation exchange capacity 41.0 mmolc kg-1, non-saline and non-sodic).
Treatments consisted of four salinity levels (T) of the irrigation water (T10.7; T21.4; T32.1 and T42.8 dS m-1, 25 ºC), tested on five clones (C) of dwarf-precocious cashew: CCP06 (C1), CCP09 (C2), EMBRAPA50 (C3), EMBRAPA51 (C4) and CCP1001 (C5), in a randomized block experimental design, with four replications, in a 4 x 5 factorial scheme. Experimental units were represented by five pots with one plant per pot. A randomized rearrangement of the pots inside each block was made biweekly.
Irrigation waters were prepared by the addition of NaCl, CaCl2.2H2O and MgCl2.6H2O, to obtain the desired electrical conductivity (ECw) for each treatment, maintaining an equivalent ratio of 7:2:1 for Na, Ca and Mg, respectively. Trials were set up in plastic containers with 25 cm height and diameter of 10 cm, with holes at the bottom. Containers were filled with substrate composed of the soil material and earthworm humus, in the proportion of 29:1 (weight basis); the substrate was previously treated with 'methyl bromide' (150 mL m-3).
Liming was made by the BS method one month before sowing, to increase the BS percentage to 70% (Osaki, 1991); calcium hydroxide was used as corrective. After liming, phosphate fertilizer (1.38 g of P2O5 per pot) was applied according to the recommendation of EMBRAPA (1993). Two applications of 50 mL of a solution containing 1.0 g L-1 of the fertilizer (composition: Total nitrogen (N) - 19%; Nitrate - 5.5%; Amonium - 3.5%; Amide - 10%; P2O5 - 19%; K2O - 19%; Fe - 1000 mg kg-1; Mn - 500 mg kg-1; B - 200 mg kg-1; Zn - 150 mg kg-1; Mo - 70 mg kg-1) were made in each container, at 64 and 74 days after sowing (DAS). The substrate was leached three times, the first two with 1.0 L of water from a local water-supply system (ECw = 1.16 dS m-1), and the last with 0.5 L of distilled water in order to reduce the salinity. Seeds were submitted to thermal shock for uniform germination, being submerged in water, at the temperature of 40ºC, returning gradually to the environmental conditions over a period of 24 h; before planting seeds were treated with 'Benomyl' (200 g of the product for 100 g of seeds); one seed per container was inserted in the vertical position (3 cm depth), with the base turned upwards (EMBRAPA, 1993).
Irrigation was performed after sowing, late in the afternoon, 300 mL per container, each other day until 64 DAS. Thereafter, water was applied every three days. Irrigation volumes were adopted starting from 67 DAS and were different for each treatment as a function of the evapotranspiration demand, estimated by the difference between the applied (VA) and drained (VD) volume, evaluating the volume of drained water and the consumption of water by the plants. The irrigation volume (mL) was calculated based on the evapotranspirated volume [IV = (VAVD) / (1LF)] to replenish the soil moisture to container capacity and to obtain a leaching fraction (LF) of approximately 0.2.
At 100 DAS, records were taken on: plant height (PH); leaf area (LA) obtained by multiplying the length of the leaf (L) by its maximum width (W) and the adjustment factor f (0.6544), determined from evaluating the plants of two blocks through LA = (L x W)f (cm2), knowing the real area of the leaves and the relationship between leaf disks of known area and their respective dry matter weight (Fernandes, 2000); leaf water content (LWC) by the difference between the fresh (FW) and dry weight (DW) of the leaf, respectively; dry matter of the shoot (DMS), root (DMR) and total (DMT), and root/shoot ratio (R/S = DMR/DMS). From the leaf area and dry matter data the following indexes were calculated: leaf area ratio (LAR = LA/DMT; cm2 g-1); absolute growth ratio [AGR = (DMTfDMTi)/t] (g day-1), where DMTi and DMTf are the initial and final total dry matter and t the time interval between samplings (100 - 50 days); relative growth ratio (RGR = AGR / DMT; mg mg-1 day-1) and net assimilation ratio (NAR = RGR / LAR) (mg cm-2 day-1) were determined by the procedures described by Benincasa (1988) and Medeiros et al. (1990). Real evapotranspiration (ETr) was also evaluated during 67-85 and 86-97 DAS, estimated through the water balance using the equation proposed by Van Hoorn & Van Alphen (1994): ETr = [(VAVD) x 10-3 / [(p x D2 / 4) x IF)], where IF is the irrigation frequency or interval (days) and D is the exposed diameter of the container (0.10 m).
After separation of roots, the soil was submitted to the following analyses: pH; moisture at saturation (%); electrical conductivity of the saturation extract (ECse), and sodium adsorption ratio (SAR), according to methodologies of EMBRAPA (1997). The osmotic pressure of the soil solution (OP) at the container capacity (CC) was estimated by the relation between electrical conductivity of the saturation extract and osmotic pressure (OP = 0.36 x ECse) (Daker, 1988), and considering the soil moisture content at CC as half of the saturation percentage.
Data were submitted to ANOVA and the F test. Polynomial regression analysis was performed for the factors 'salinity levels of the irrigation water' and 'clones'; the Tukey test was applied (P = 0.05) for the comparison of the mean values. For analysis purposes, LA and AGR data were transformed to  ; DMS, DMR, DMT, R/S ratio, RGR, NAR data to
; DMS, DMR, DMT, R/S ratio, RGR, NAR data to  ; and the data of LWC to arc sin
; and the data of LWC to arc sin  , according to the recommendations of Ferreira (2000). The discussion of data, however, was based on the original values aiming a better understanding of the effects of the studied factors.
, according to the recommendations of Ferreira (2000). The discussion of data, however, was based on the original values aiming a better understanding of the effects of the studied factors.
RESULTS AND DISCUSSION
Plant height was affected (P < 0.01) by the salinity of the irrigation water, but, no differences were observed for the clones (Table 1). The effect of the saline treatments on plant height was linear and negative (Figure 1A), with approximately 10% decrease for unit increment of ECw in comparison to T1. Meireles (1999), studying two of the dwarf-precocious cashew clones used in this work (CCP06 and CCP1001), irrigated with water containing NaCl and CaCl2 (1:1), also observed the reduction of plant height with the increase of ECw above 0.7 dS m-1, at 60 DAS.
Leaf area was affected (P < 0.01) by the electrical conductivity of the irrigation water and varied among clones. Even though the effect of salinity did not depend on genotype, the leaf area decreased linearly with the increase of the salinity (Figure 1B), the reduction being approximately 8% (in relation to T1) for unit increase of ECw. Similar behaviour was observed by Bezerra (2001), working with the clones CCP1001 and CCP06 of dwarf-precocious cashew, for the electrical conductivity of the irrigation water from 0.5 to 5.5 dS m-1; at 40 DAS the relative reduction of the leaf area was of 8.74% for each increased unit in ECw. The reduction of the leaf area and, consequently, of the photosynthesis, contributed in a certain way for the adaptation of crop to salinity (Läuchli & Epstein, 1990; Plant, 1994). The clone C3 (EMBRAPA50) presented larger leaf area, over 25% superior to the other (Table 1), and also presented a larger (P < 0.01) number of leaves (Carneiro, 2001).
Leaf water content (LWC) increased linearly with the levels of ECw (Figure 1C), with increment of 1.30, 2.61 and 3.91% for T2, T3 and T4, in comparison to T1, respectively. This may be attributed to osmotic adjustments, with plants accumulating ions in cell vacuoles or synthesizing organic compounds (Shannon, 1997), thereby lowering the internal water potential and guaranteeing the absorption of water to maintain cell turgescence. However, the water inside the cells, although in larger amount, starts to have a lower state of energy as a result of the decrease of the osmotic potential, resulting in lower plant growth (Van Hoorn et al., 1993).
LWC varied among clones (P < 0.01) independent of water salinity, once the interaction between these factors was not significant. The water content of the leaves of the clones EMBRAPA51 (C4), CCP06 (C1) and EMBRAPA50 (C3) was higher perhaps because of their genetic constitution, although there was no difference between the last one in relation to the others.
DMS and DMR production decreased (P < 0.01) linearly with the salinity of the irrigation water (Figure 2A and B), with reductions of nearly 13.98% for DMS and 15.39% for DMR, per unit increase in salinity over that of the water with lowest salt concentration (0.7 dS m-1). Differences were also observed among clones for fresh weight accumulation of shoot and root (Table 1), independent of the water salinity. Plants of clone C3 tended to form more DMS and those of C2, C3 and C5 more DMR, while the lowest values for these variables were observed for C4. DMT also was affected by the salinity of the irrigation water and varied among clones (P < 0.01). However, there was no interactive effect of the salinity on the studied genotypes.
The effect of the saline treatments on DMT was linear and negative (Figure 2C), with a decrease of 14.42% per unit increase in water salinity, in relation to T1. Based on the criterion of Ayers & Westcot (1991), it is observed that for the formation of the studied rootstocks, water up to EC of 1.39 dS m-1 may be used, and would correspond to a maximum loss of 10% in the total dry matter production of the plants. When the isolated effect of clones was analyzed (Table 1), the genotype EMBRAPA51 (C4) was found to be least vigorous, with a tendency to produce less DMT as compared to the other clones; in relation to C3 (EMBRAPA50), EMBRAPA51 produced 31.75% less DMT.
The salinity of the irrigation water did not influence the root/shoot ratio (R/S), probably because of the small variation in terms of relative decrease for each unit increase in ECw that occurred for DMS and DMR of the plants (13.98 and 15.39%, respectively). Evaluating cashew rootstocks (CCP06 and CCP1001) at 40 DAS, Bezerra (2001) observed increments of the R/S ratio with increasing water salinity, as a result of higher reduction in shoot fresh weight. The effects on shoots were also more pronounced at the beginning, probably because of the fact that, in relation to roots, salinity prolonged the germination of seeds, but the differences disappeared in time, as plants recovered growth. The increase of the R/S ratio is an attempt of the plant to explore a larger soil volume (Ludlow & Muchow, 1990). Since this experiment used containers of 2.0 L, the possibility of the plant to explore larger soil volume became limited and similar effects of salinity occurred after 100 days. In relation to the R/S ratio, effects were observed (P < 0.01) among genotypes. Even though no statistical differences were observed, clones CCP1001 (C5) and CCP09 (C2) reached higher R/S ratios in relation to the other genotypes; that may be explained by higher proportion of DMS in relation to DMT produced by C1, C3 and C4 (Table 1).
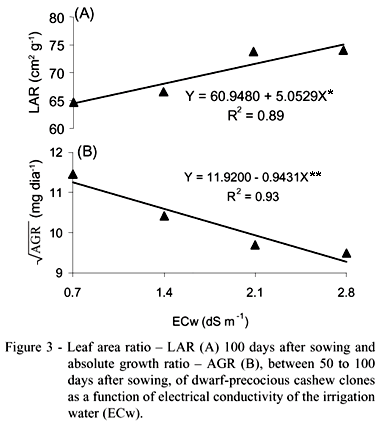
The effect of the saline treatments on LAR was linear (P < 0.05) and increasing, because the effect had been more pronounced on the production of DMT than on LA. LAR is an indicator of the relative dimension of the assimilation apparatus it relates the leaf area to the dry matter resulting from the photosynthesis. According to the concept of LAR, a larger proportion of the photo assimilate products was used in the formation of the photosynthetic apparatus, with increasing salinity. LAR increased by 7.84% per unit increment of salinity above that of the water with lowest salt concentration (Figure 3A). LAR varied among clones (P < 0.01); although effects of the saline treatments on LAR were observed, it was not interactive with clones (Table 2), similar to those observed by Bezerra (2001), also working with rootstocks of dwarf-precocious cashews.
Higher leaf area ratio was observed for EMBRAPA51 (C4), similar to the water content in the leaves (Table 2), not differing only from C3 (EMBRAPA50). The increment of LAR for EMBRAPA51 was 24.74%, in comparison to the genotype CCP09, which had the smallest relative dimension of the assimilation apparatus. Usually, as the leaf grows, lower is the photo assimilate proportion in the leaf blade, in other words, the photo assimilate export to other parts of the plant is higher. For the EMBRAPA51 clone, photosynthesis products were more used for leaf formation.
AGR was influenced (P < 0.01) by the salinity of the irrigation water (T), without, however, varying among clones (C), and as a function of the interaction T x C. The saline treatments affected the absolute growth rate of plants, with a decrease in relation to T1 of 15.53% per unit increase in ECw (Figure 3B). Evaluating the effect of salinity on two cashew rootstocks (CCP06 and CCP1001) at 40 DAS, Bezerra (2001) also observed a reduction of plant growth, however not so sharp (between 4.6 and 4.7% per unit increase of ECw) in comparison to the present study. The higher reduction found in this study can be attributed to the larger period (100 days) to which the plants were exposed to the adverse salinity conditions.
In opposition to the observation for AGR, salinity did not affect RGR, nor NAR, that is, the use of saline water did not affect the crop efficiency in producing new biomass per unit of pre-existent biomass, nor the photosynthetic efficiency, but both indexes varied among clones. Regarding salinity, similar results were observed by Bezerra (2001) for two cashew clones grown up to 40 DAS. The clone CCP1001 (C5) had the tendency to produce smaller NAR (Table 2), equal to 59.32% of EMBRAPA51 (C4) production; it presented lower efficiency in relation to genotype CCP1001, in converting photo assimilates in new photosynthetic products, the net photosynthesis rate per unit of leaf area being also low.
The leaching fraction decreased in the second period (86-97 DAS) of evaluation (Figure 4A), which could be explained by the smaller volume of applied water (VA), once the water consumption of the plants submitted to the highest levels of salinity decreased with time (Table 3). The real evapotranspiration (ETr) decreased with the increase of ECw along the evaluation (Figure 4B). A relative decrease of 18.56% occurred between T1 and T4 in the interval 67-85 DAS, and of 32.89% in the period 86-97 DAS. The osmotic effect of salinity is evidenced, indicating that the plants suffered water stress induced by the saline stress; the concentration of soluble salts in the root zone results in the reduction of the osmotic potential of soil solution and reduces the water flow in the direction soil®plant®atmosphere, with consequent reduction of the plant transpiration, affecting its growth (Rhoades & Loveday, 1990). A direct relationship exists between evapotranspiration (ET) and the growth of the plants, ET and growth being at their maximum when the supply of water is plentiful (Doorenbos & Kassam, 1994). On the other hand, under water restriction, reduction in growth takes place as observed in the present study with the increase of ECw.
Regarding the clones, C3 (EMBRAPA50) had the highest average ETr in both periods of evaluation (10.60 and 10.21 mm day-1, during 67-85 and 86-97 DAS, respectively), a fact that proves the close relationship between the transpiration and the leaf surface (Plant, 1994; Shannon, 1997). As previously discussed, only the genotype EMBRAPA50 stood out in leaf area, with a value statistically higher than the others (Table 1), resulting in higher production of total biomass. Conversely, ETr of EMBRAPA51 plants (8.20 mm day-1, during the period of 67-85 DAS and 7.20 mm day-1 in the period of 86-97 DAS) was lower, as a consequence of their lower leaf area, accumulating less total dry matter. These considerations are in agreement with Plant (1994) who mentioned that the decrease in the growth of the leaf area, and consequently in the yield of the plants, could be considered as the first visible reaction to the water deficit.
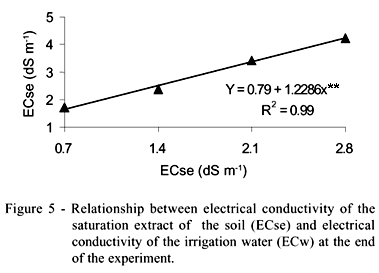
The increase of the irrigation water salinity resulted in increases of the electrical conductivity of the soil saturation extract (ECse), varying from 1.71 to 4.23 dS m-1 for ECw of 0.7 (T1) and 2.8 dS m-1 (T4), respectively (Table 4). According to the coefficient 'b' of the equation presented in the Figure 5, ECse was on average 1.23 times the value of ECw.
The estimated values of the osmotic pressure of the soil solution increased with the raising of ECw and at the end of the experiment, varying from 124 kPa for T1 to 308 kPa for T4, which explains the observed reduction of the ETr for the higher salinity levels, and confirms observations of Rhoades & Loveday (1990). The osmotic effect in the clone C4 (EMBRAPA51) was higher; this genotype presented the highest water content in the leaves (Table 1), probably because of the mechanism of osmotic adjustment, in which the plants accumulate the absorbed ions in the cellular vacuole or synthesize organic compounds to reduce the internal water potential, guaranteeing water uptake to maintain the turgescence of the cells (Shannon, 1997).
The tolerance of the crop to salinity is usually expressed based on the response to soil salinity in terms of electrical conductivity of the saturation extract (ECse). As previously discussed (Table 1), the production of DMT decreased for the cashew rootstocks, with a relative decrease of 10.59% (Figure 6) per unit increment of ECse starting from 0.7 dS m-1, which corresponds to ECse of 1.71 dS m-1. According to Maas (1984), crops which produce 90% of their relative yield with a salinity threshold (expressed in terms of ECse) ranging on 1.3 to 3.0 dS m-1 are classified as moderately sensitive to salinity: therefore, the clones of dwarf cashew studied can be considered moderately sensitive to salinity, once the relative production of DMT was 90% in conditions of ECse of 2.65 dS m-1.
CONCLUSIONS
The dwarf-precocious cashew is 'moderately sensitive' to salinity in the phase of rootstock formation, with threshold salinity of 1.39 dS m-1 in terms of electrical conductivity of the water and of 1.71 dS m-1 in terms of soil saturation extract.
The water content of the leaves and the leaf area ratio increase linearly with the salinity of irrigation water, but it does not affect the relative growth rate and the net assimilation rate of the rootstocks.
The worst and the best growth indexes were observed for the clones EMBRAPA51 and EMBRAPA50, respectively, independent of the salinity effect.
Received August 26, 2002
Accepted October 29, 2003
- AUDRY, P.; SUASSUNA, J. A salinidade das águas disponíveis para a pequena irrigação no sertão do Nordeste: caracterização, variação sazonal, limitação de uso. Recife: CNPq, 1995. 128p.
- AYERS, R.S.; WESTCOT, D.W. A qualidade da água na agricultura Campina Grande: UFPB, 1991. 218p. (Estudos FAO: Irrigação e Drenagem, 29).
- BENINCASA, M.M.P. Análise de crescimento de plantas Jaboticabal: FUNEP, 1988. 42p.
- BEZERRA, I.L. Produção de mudas enxertadas de cajueiro anão precoce, usando águas de diferentes salinidades. Campina Grande: UFPB, 2001. 85p. (Dissertação - Mestrado).
- CARNEIRO, P.T. Germinação e desenvolvimento inicial de clones de cajueiro anão-precoce sob condições de salinidade. Campina Grande: UFPB, 2001. 84p. (Dissertação - Mestrado).
- DAKER, A. A água na agricultura 7.ed. Rio de Janeiro: Freitas Bastos, 1988. v.3, 543p.
- DOORENBOS, J.; KASSAM, A.H. Efeito da água no rendimento das culturas Campina Grande: UFPB, 1994. 306p. (Estudos FAO: Irrigação e Drenagem, 33).
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Aspectos agroeconômicos sobre a cultura do cajueiro Fortaleza: EMBRAPA, 1993. 124p.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Manual de métodos de análise de solo 2.ed. Rio de Janeiro: EMBRAPA, 1997. 212p.
- FAGERIA, N.K. Solos tropicais e aspectos fisiológicos das culturas Brasília: EMBRAPA, CNPAF, 1989. 425p. (Documento, 18).
- FERNANDES, P.D. Análise de crescimento e desenvolvimento vegetal Campina Grande: UFPB, Departamento de Engenharia Agrícola, 2000. 22p.
- FERREIRA, O.S.; MATOS, N.N.; MENESES JR., J.; BARROS, L. de M.; LIMA JR., A.; SILVEIRA, J.A.G. da. Avaliação inicial da tolerância ao estresse salino em materiais de cajueiro (Anacardium occidentale L.) através de índices de crescimento. (Compact disc) In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 16., Fortaleza, 2000. Anais. Fortaleza: SBF, 2000.
- FERREIRA, P.V. Estatística experimental aplicada à agronomia Maceió: UFAL; EDUFAL; FUNDEPES, 2000. 422p.
- FNP CONSULTORIA E COMÉRCIO. Agrianual 98: Anuário estatístico de agricultura brasileira. São Paulo, 1998. 481p.
- IZZO, R.; NAVARI-IZZO, F.; QUARTACCI, F. Growth and mineral absorption in maize seedlings as affected by increasing NaCl concentrations. Journal of Plant Nutrition, v.14, p.687-699, 1991.
- LÄUCHLI, A.; EPSTEIN, E. Plant responses to saline and sodic conditions. In: TANJI, K.K. (Ed.) Agricultural salinity assessment and management. New York: ASCE, 1990. cap.6, p.113-1137.
- LUDLOW, M.M.; MUCHOW, R.C. A critical evolution of traits for improving crop yields in water-limited environments. Advances in Agronomy, v.43, p.107-153, 1990.
- MAAS, E.V. Crop tolerance. California Agriculture, v.38, p.20-21, 1984.
- MEDEIROS, J.F. Qualidade de água de irrigação e evolução da salinidade nas propriedades assistidas pelo 'GAT' nos estados de RN, PB e CE. Campina Grande: UFPB, 1992. 173p. (Dissertação - Mestrado).
- MEDEIROS, J.G.; PEREIRA, W.; MIRANDA, J.E.C. Análise de crescimento em duas cultivares de batata-doce (Ipomoea batatas L. Lan). Revista Brasileira de Fisiologia Vegetal, v.2, p.23-29, 1990.
- MEIRELES, A.C.M. Salinidade da água de irrigação e desenvolvimento de mudas de cajueiro anão-precoce (Anacardium occidentale L.) Fortaleza: UFC, 1999. 60p. (Dissertação - Mestrado).
- OLIVEIRA, V.H. de; CRISÓSTOMO, L.A.; MIRANDA, F.R. de. Produtividade de clones de cajueiro anão-precoce (Anacardium occidentale L) irrigados no município de Mossoró-RN Fortaleza: EMBRAPA, CNPAT, 1998. 6p. (Comunicado Técnico, 14).
- OSAKI, F. Calagem e adubação 2.ed. Campinas: Instituto Brasileiro de Ensino Agrícola, 1991. 503p.
- PLANT, Z. Photosyinthesis in plant/crops under water and salt stress. In: PESSARAKALI, M. (Ed) Hanbook of plant and crop stress New York: Marcel Dekker, 1994. p.587-603.
- RAMOS, A.D.; BLEICHER, E.; FREIRE, F. das C. de O.; CARDOSO, J.E.; PARENTE, J.I.G.; BARROS, L. de M.; CRISÓSTOMO, L.A.; FROTA, P.C.E.; CORREA, M.P.F.; PAULA PESSOA, P.F.A. de; MELO, Q.M.S.; OLIVEIRA, V.H. de. A cultura do caju Brasília: SPI, 1996. 96p. (Coleção Plantar, 34).
- RHOADES, J.D.; LOVEDAY, J. Salinity in irrigated agriculture. In: STEWART, D.R.; NIELSEN, D.R. (Ed.) Irrigation of agricultural crops. Madison: ASA; CSSA; SSSA, 1990. p.1089-1142. (Agronomy, 30).
- SHANNON, M.C. Adaptation of plants to salinity. Advances in Agronomy, v.60, p.75-120, 1997.
- VAN HOORN, J.W.; VAN ALPHEN, J.G. Salinity control. In: RITZEMA, H.P. (Ed.) Drainage principles and applications. Wageningen: ILRI, 1994. p.533-600. (Publication, 16).
- VAN HOORN, J.W.; KATERJI, N.; HAMDY, A.; MASTRORILLI, M. Effect of saline water on soil salinity and on water stress, growth, and yield of wheat and potatoes. Agricultural Water Management, v.23, p.247-265, 1993.
- VIÉGAS, R.A.; SILVEIRA, J.A.G. da; LIMA JR., A.R. de; QUEIROZ, J.E.; FAUSTO, M.J.M. Effects of NaCl-salinity on growth and inorganic solute accumulation in young cashew plants. Revista Brasileira de Engenharia Agrícola e Ambiental, v.5, p.216-222, 2001.
Publication Dates
-
Publication in this collection
16 Feb 2004 -
Date of issue
Feb 2004
History
-
Accepted
29 Oct 2003 -
Received
26 Aug 2002