Abstracts
Tanner grass (Brachiaria radicans Napper) is a forage plant that is adapted to well-drained soils or wetlands, and responds well to nitrogen (N) fertilization. The assimilation of N involves the nitrate reductase (NR) enzyme, and its activity seems to be dependent on N supply. Molybdenum (Mo) is also important because it is a cofactor of NR. In this study, the variables of an in vivo assay were optimized for measuring nitrate reductase activity (NRA) in the leaves and stem tissues. This method was used to evaluate NO3- metabolism in plants fertilized with NaNO3, NH4Cl or urea, in association with or without application of H2MoO4, aiming to provide guidelines for N management of this species. The best conditions to determine NRA involved the incubation of 300 mg of tissues in a medium composed of 200 mmol dm3 phosphate buffer (pH 7.4), 60 mmol dm3 KNO3, 10 cm³ dm3 n-butanol, 0.1 cm³ dm3 detergent (triton-X-100®), under vacuum and in the dark for a period of 60 to 100 minutes. Leaves showed NRA levels two to three times higher than stems. Although there were some interactions between treatments, stem fresh weight and NRA were not affected by N sources. Plants fertilized with NaNO3 showed the best growth and NRA values when compared with NH4Cl and urea, which had, respectively, the lowest and intermediate scores. The application of Mo in the absence of N improved NRA and did not affect leaf and stalk growth. In the presence of N, the Mo levels applied limited leaf NRA and plant development.
Brachiaria arrecta; nitrogen; molybdenum; enzyme
A 'tanner grass' (Brachiaria radicans Napper) é uma planta forrageira que se adapta bem tanto em solos bem drenados como em encharcados, e responde bem à adubação nitrogenada. A assimilação do nitrogênio (N) envolve a enzima nitratro redutase (NR) cuja atividade é dependente do suprimento de N. O Molibdênio (Mo) também é importante por ser um co-fator da RN. Neste trabalho foi otimizado um procedimento para a determinação da atividade da redutase do nitrato (NRA) in vivo para tecidos de folha e caule. Este método foi aplicado para avaliar plantas fertilizadas com NaNO3, NH4Cl ou uréia, com e sem a aplicação de H2MoO4, visando contribuir no balizamento da adubação nitrogenada para esta espécie. As melhores condições para a determinação da NRA foram: incubação de 300 mg de tecidos em um meio composto por tampão fosfato 200 mmol dm3 (pH 7,4); KNO3 60 mmol dm3; n-butanol 10 cm³ dm3; triton-x-100® 0,1 cm³ dm3; infiltração a vácuo no escuro; por período de 60 a 100 minutos. As folhas apresentaram NRA duas a três vezes maior que os caules. A matéria fresca do caule e a NRA não foi afetada pelas fontes de N. As plantas adubadas com NaNO3, NH4Cl e uréia apresentaram, respectivamente, os maiores, os menores e intermediários crescimento e NRA. A aplicação de Mo na ausência de N aumentou a NRA e não afetou o crescimento, mas, na presença de N, limitou a NRA nas folhas e o crescimento das plantas.
Brachiaria arrecta; nitrogênio; molibdênio; enzima
SOILS AND PLANT NUTRITION
Nitrate reductase activity in leaves and stems of tanner grass (Brachiaria radicans Napper)
Atividade da redutase do nitrato em folhas e caules de 'tanner grass'(Brachiaria radicans Napper)
Jairo Osvaldo Cazetta* * Corresponding author < cazetta@fcav.unesp.br> ; Luciana Cristine Vasques Villela
UNESP/FCAV - Depto. de Tecnologia - Via de Acesso Prof. Paulo Donato Castellane s/n, 14884-900 - Jaboticabal, SP - Brasil
ABSTRACT
Tanner grass (Brachiaria radicans Napper) is a forage plant that is adapted to well-drained soils or wetlands, and responds well to nitrogen (N) fertilization. The assimilation of N involves the nitrate reductase (NR) enzyme, and its activity seems to be dependent on N supply. Molybdenum (Mo) is also important because it is a cofactor of NR. In this study, the variables of an in vivo assay were optimized for measuring nitrate reductase activity (NRA) in the leaves and stem tissues. This method was used to evaluate NO3- metabolism in plants fertilized with NaNO3, NH4Cl or urea, in association with or without application of H2MoO4, aiming to provide guidelines for N management of this species. The best conditions to determine NRA involved the incubation of 300 mg of tissues in a medium composed of 200 mmol dm3 phosphate buffer (pH 7.4), 60 mmol dm3 KNO3, 10 cm3 dm3 n-butanol, 0.1 cm3 dm3 detergent (triton-X-100®), under vacuum and in the dark for a period of 60 to 100 minutes. Leaves showed NRA levels two to three times higher than stems. Although there were some interactions between treatments, stem fresh weight and NRA were not affected by N sources. Plants fertilized with NaNO3 showed the best growth and NRA values when compared with NH4Cl and urea, which had, respectively, the lowest and intermediate scores. The application of Mo in the absence of N improved NRA and did not affect leaf and stalk growth. In the presence of N, the Mo levels applied limited leaf NRA and plant development.
Key words:Brachiaria arrecta, nitrogen, molybdenum, enzyme
RESUMO
A 'tanner grass' (Brachiaria radicans Napper) é uma planta forrageira que se adapta bem tanto em solos bem drenados como em encharcados, e responde bem à adubação nitrogenada. A assimilação do nitrogênio (N) envolve a enzima nitratro redutase (NR) cuja atividade é dependente do suprimento de N. O Molibdênio (Mo) também é importante por ser um co-fator da RN. Neste trabalho foi otimizado um procedimento para a determinação da atividade da redutase do nitrato (NRA) in vivo para tecidos de folha e caule. Este método foi aplicado para avaliar plantas fertilizadas com NaNO3, NH4Cl ou uréia, com e sem a aplicação de H2MoO4, visando contribuir no balizamento da adubação nitrogenada para esta espécie. As melhores condições para a determinação da NRA foram: incubação de 300 mg de tecidos em um meio composto por tampão fosfato 200 mmol dm3 (pH 7,4); KNO3 60 mmol dm3; n-butanol 10 cm3 dm3; triton-x-100® 0,1 cm3 dm 3; infiltração a vácuo no escuro; por período de 60 a 100 minutos. As folhas apresentaram NRA duas a três vezes maior que os caules. A matéria fresca do caule e a NRA não foi afetada pelas fontes de N. As plantas adubadas com NaNO3, NH4Cl e uréia apresentaram, respectivamente, os maiores, os menores e intermediários crescimento e NRA. A aplicação de Mo na ausência de N aumentou a NRA e não afetou o crescimento, mas, na presença de N, limitou a NRA nas folhas e o crescimento das plantas.
Palavras-chave:Brachiaria arrecta, nitrogênio, molibdênio, enzima
INTRODUCTION
Tanner grass is the common name for Brachiaria radicans Napper (Skerman & Riveros, 1990), lately described as Brachiaria arrecta (Souza, 1999). This forage grass is very competitive when compared with other forage species, and grows well under tropical conditions (Mitidieri, 1983; Skerman & Riveros, 1990). As in the majority of crops, nitrogen (N) is one of the most important nutrients in grasses. To improve the efficiency of N fertilization, more information on the processes of N utilization is required for each plant species (Baer & Collet, 1981). There is evidence suggesting that nitrate reductase activity (NRA) is the main limiting step in N assimilation in most plants (Srivastava, 1990; Lea, 1997). Leaf NRA seems to be dependent on the source of N (Roberts & Menary, 1994), and on a continuous supply of nitrate (NO3-) through the xylem (Barthes et al., 1996; Kawachi et al., 2002). Molybdenum (Mo) is also important because it is a cofactor of NR (Lea, 1997). Thus, some interactions between the effects of N sources and Mo fertilization on NRA have been reported (Dordas et al., 2001). The NRA levels in a plant are of help in the study of plant metabolism and its complex relationship with the environment (Muller & Janieshch, 1993; Faleiros & Cazetta, 1996; Viégas et al., 1999; Stolz & Bazu, 2002), and one of the challenges of research in N metabolism is to develop approaches to analyze this complex interaction (Stitt et al., 2002).
NRA can be measured by either an in vivo or in vitro assay (Viégas et al., 1999). The in vitro assay is accomplished by extracting the enzymatic fraction from the tissue (Lillo, 1983; Viégas et al., 1999), and by measuring the reaction in an appropriate buffered medium (Aslam et al., 1984; Lillo, 1983). On the other hand, the in vivo assay can be carried out easily, without enzyme extraction, by incubating tissue samples in a buffered medium containing NO3-, solvent and/or detergent, and by measuring NO2- produced during an adequate period of time, in the dark (Jaworski, 1971; Jones & Sheard, 1977; Harper, 1981). Nitrite production in the in vivo assay depends on the diffusion of NO3- into the tissue, to reach the cytoplasm of cells where the enzyme is located. Therein, nitrate reductase catalyses the conversion of NO3- to NO2-, by using the reducing power of NADH (Nicotinamide Adenine Dinucleotide, reduced form) coming from the respiratory oxidative pathways. Under normal conditions, NO2- produced in green tissues is transported into chloroplasts and reduced to ammonia, using the reducing power of reduced ferredoxin, derived from photosynthetic electron transport (Lea, 1997). However, during the in vivo assay procedure, the tissue is kept in the dark, which inhibits NO2- reduction. This leads to NO2- accumulation, which drives its diffusion from the tissue toward the incubation medium, where it is measured as the product of NR activity. Thus, NO2- present in the medium not only depends on the NRA, but also on the diffusion processes.
Although NR always catalyzes the same reaction, its location within tissues, structure, and regulatory mechanisms are organism-dependent (Stolz & Bazu, 2002). Due to differences between species, or even between various plant parts, it is best to optimize the conditions for the in vivo assay, before NRA determinations are made (Lin & Kao, 1980; Faleiros & Cazetta, 1996). Therefore, the first objective of this research was to establish the optimum conditions for tanner grass leaf and stem NRA determinations, using the in vivo assay method. The second objective was to evaluate NRA in leaves and stems of tanner grass plants fertilized with NaNO3, NH4Cl, or urea, with or without Mo, for the purpose of providing guidelines for N management of this species in the future.
MATERIAL AND METHODS
Plants were collected in Jaboticabal-SP, Brazil (21°15'17''S and 48°19'20''W), in December of 2002, and separated into leaf blades and stems (stalk plus leaf sheath adhering to the stalk). The central portion of leaves was cut into discs with a diameter of 1.0 cm. The stems were cut transversally into 2 mm long pieces. Both tissues were assayed for their in vivo NRA. The basal assay procedure used was a modification of the Jaworski (1971) method, described by Faleiros & Cazetta (1996), based on the incubation of fresh tissue (300 mg) in 10.0 cm3 of a medium containing 25 mmol dm3 phosphate buffer (pH 7.2), 25 mmol dm3 KNO3, 30 cm3 dm3 n-propanol, and 30 cm3 dm3 of triton X-100® (t-octyl-phenoxy-polyethoxy-ethanol, Rohm & Haas Co: U.S.A.). In order to induce anaerobic conditions in the incubation medium, the system was subjected to vacuum infiltration three times, as recommended by Klepper et al. (1971) to remove air from the tissue. The medium was maintained in the dark at 30ºC for 60 minutes.
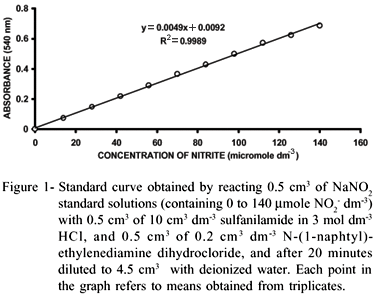
The NO2- produced by action of the NR enzyme was determined by drawing an 0.5 cm3 aliquot of the incubation medium, and treating this sample with 0.5 cm3 of 10 g dm3 sulfanilamide in 3 mol dm3 HCl and 0.5 cm3 of 0.2 g dm3 N-(1-naphtyl)-ethylenediamine dihydrochloride. After 20 minutes, the solution was diluted to 4.5 cm3 with deionized water, and the absorbance (540 nm) was measured using an Ultrospec 2000® spectrophotometer (Pharmacia Biotech: England). In order to calculate the amount of NO2- contained in the sample, a standard curve was prepared in the same way as the sample, but using aliquots of 0.5 cm3 of NaNO2 standard solutions (containing from 0 to 140 mmol dm3 NO2-).
The optimization of the NRA assay was developed by evaluating the best incubation time (ranging from 20 to 120 minutes), pH level (6.0 to 8.0), surfactant compounds (solvents: ethanol, n-propanol, n-butanol, ketone, ethyl acetate; and detergents: triton X100® or tween 80® (poly-oxy-ethylene sorbitan monooleate, Atlas Chemie: Germany), as well as the best concentration for phosphate (ranging from 0 to 400 mmol dm 3), NO3- (0 to 100 mmol dm3), solvent (0 to 100 cm3 dm3), and surfactant (0 to 100 g dm3), for the incubation medium. The effect of vacuum infiltration on the performance of the assay was also studied. The NO2- diffusion results were expressed as micromoles of NO2- per gram of fresh tissue, after a given incubation time, and NRA results were expressed as micromoles of NO2- produced per g of fresh tissue per hour (mmol g1 h1 NO2-).
The optimized NRA method was applied to study NO3- assimilation in stems and leaves of tanner grass plants fertilized with different sources of N, and varying levels of Mo. This experiment was carried out using a medium-textured Typic Hapludox, collected from a field previously cultivated with corn (Zea mays L.). The initial soil properties were: pH in 0.01 mol L3 CaCl2 solution = 4.7; organic matter = 12 g dm3; P = 11.0 mg dm3; K = 1.5 mmolc dm3; Ca = 16.0 mmolc dm3; Mg = 5.0 mmolc dm3; H++Al3+ = 37.0 mmolc dm3, cation exchange capacity (CEC) = 59.5, and base saturation = 37.8%. To reduce acidity, CaO was added and blended into the soil at a rate necessary to obtain a base saturation of 60%. The mixture was kept moist, and let stand for 15 days. The soil was then fertilized with Ca(H2PO4)2, K2SO4, and Mg(SO4)2, at levels corresponding to 80 kg ha1 P2O5, 60 kg ha1 K2O, and 30 kg ha1 Mg, respectively, and then transferred to 5 dm3 pots. Rooted sprouts of tanner grass of similar size were collected from the field, and individually planted in 32 pots.
The eight treatments consisted of three sources of N: NaNO3, NH4Cl, and urea. All N sources were applied at a rate corresponding to 300 kg N ha1, combined with two levels of Mo fertilization (0 and 20 kg Mo ha1, as H2MoO4), plus two control treatments: no N or Mo fertilization, and fertilization with Mo, applied without N. The Mo fertilizer was added to appropriate treatments before planting. The different sources of N were applied with the irrigation water, divided into four weekly applications, which were started 10 days after planting.
The experiment was organized as a completely randomized design, with eight treatments and four replications, where each 5 dm3 pot containing one plant comprised an individual experimental unit. Plants were maintained in a growth chamber with a photoperiod of 16 h (176 mmol PPFD m2 s1) provided by incandescent and fluorescent bulbs. The temperature was maintained between 20 and 35°C (night and day, respectively) and the experiment was conducted for 40 days after planting. Plants were then harvested and separated into stems (stalk plus sheath adhering to stalk) and leaf blades and weighed, and NRA determinations were made according to the method previously described.
A factorial design (three N sources × two Mo levels, plus two controls), with four replications, was used for statistical analysis in this study. The F test was applied to detect main and interaction effects, and means were compared using Tukey test (P < 0.05). Analyses of variance and factorial analyses were performed using SAS® (SAS version 6.11 software package, Statistical Analysis System Institute: U.S.A).
RESULTS AND DISCUSSION
Results obtained for the standard curve (Figure 1) showed high correlation (r2 = 0.999) between absorbance at 540 nm and [NO2-]. This indicates that the assay model was reliable to determine NO2- with good precision.
Stem NRA showed some variations within the first 60 minutes of incubation, but tended to be stable from 60 to 120 minutes (Figure 2), and exhibited an activity of about 20% of that observed for leaves.
NRA results for leaves revealed an increase (P < 0.05) during incubation times from 20 to 120 minutes (Figure 2). The behavior of NRA during incubation was reported to increase (Jaworski, 1971; Streeter & Bosler, 1972; Blondel & Blanc, 1975; Lin & Kao, 1980; Faleiros & Cazetta, 1996), to remain constant up to 120 minutes (Brunnetti & Hageman, 1976, Heuer & Plaut, 1978), to decrease during the first 60 minutes (Streeter & Bosler, 1972), or to decrease after 60 minutes (Klepper et al., 1971), indicating that it is dependent upon the species or tissue under study. In order to be reduced to NO2- in the in vivo assay, NO3- has to diffuse into the tissue and the NO2- produced has to diffuse into the incubation medium in order to be measured. Thus, since induced NRA is not expected to increase during incubation, increases suggest that variations may rely on diffusion processes. This possibility reinforced the need for solvents and surfactants as medium components.
The inclusion of n-butanol or n-propanol in the leaf incubation medium promoted higher NRA, when compared to the addition of ethanol. Acetone and ethyl acetate suggested intermediate promotion (Figure 3). Among the surfactants, triton-X-100® improved NRA, in comparison to tween-80® (Figure 3). When these same chemicals were included in the incubation medium for stem tissue, no differences (P > 0.05) were observed (Figure 3). The inclusion of solvents in the incubation medium was intended to increase membrane permeability. This would facilitate the influx of NO3- into tissue cells, and NO2- efflux into the medium. For triticale (Triticosecale Wittmack) propanol was more effective than butanol (Lin & Kao, 1980), but for tanner grass similar results were obtained for both alcohols (Figure 3). Butanol was chosen because of its supposedly lower toxicity and higher water solubility, compared to propanol (Jaworski, 1971).
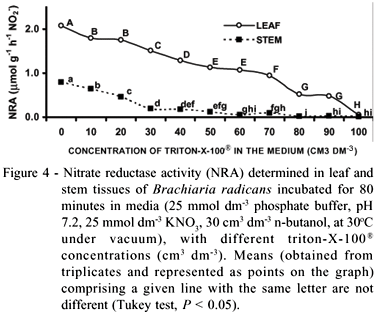
The presence of a detergent in the medium is important in order to minimize liquid surface tensions, and improve contact of the tissue with the medium. However, our data indicated that NRA decreased when triton-X-100® was added to the medium (Figure 4). By increasing the concentrations of the triton-X-100® detergent from 0 to 100 cm3 dm3 in the incubation medium, leaf NRA decreased by an average of 5.8 % for each 10 cm3 dm3 increment of triton-X-100® added. Stem tissue NRA decreased by an average of 47% when the triton-X-100® concentration in the medium was increased from 0 cm3 dm3 to 30 cm3 dm3. Higher concentrations tended to induce further reductions, but at much smaller rates (Figure 4). These results suggest that triton-X-100® could be excluded from the incubation medium for both tissues. Nevertheless, considering that the concentration required to reduce liquid tension is very low (about 0.1 cm3 dm3), and also that a mixture of surfactants tend to induce a synergistic effect (Streeter & Bosler, 1972; Tingey et al., 1974; Lawrence & Herrick, 1982; Faleiros & Cazetta, 1996), it would be advisable to include 0.1 cm3 dm3 triton-X-100®.
After choosing n-butanol as the solvent to be used, the effect of different concentrations in the medium was observed. Increasing the concentration from 0 cm3 dm3 to 10 cm3 dm3 increased NRA by an average of 5 % for leaf, and 18% for stem tissue, while concentrations higher than 10 cm3 dm3 decreased NRA. The inhibitory effect was proportional to the concentration of butanol, chiefly in the range from 10 cm3 dm3 to 70 cm3 dm3 for leaf, and from 10 cm3 dm3 to 30 cm3 dm3 for stem tissues. Comparing the results of butanol at 10 cm3 dm3 with butanol at 100 cm3 dm3, NRA was reduced by 93% and 89% for leaf and stem tissues, respectively (Figure 5).
In spite of the fact that data from the different tests were obtained on different dates, and with the use of different plants, it becomes clear that when the average values for NRA are compared, the lower activities observed in the first tests (Figure 2, 3) were mainly due to the use of excessive concentrations of surfactants (30 cm3 dm3). The best butanol concentration (30 cm3 dm3) for Cajanus cajan (Faleiros & Cazetta, 1996) promoted a significant inhibitory effect when applied to tanner grass.
Nitrate reductase activity was enhanced by increases in NO3- supply from 0 mmol dm3 to 33 mmol dm3, while no further effect was observed for [NO3-] higher than 33 mmol dm3, for either leaf or stem tissues, indicating that the optimum concentration should be between 17 mmol dm3 and 33 mmol dm3 (Figure 6). Media containing [NO3-] higher than 83 mmol dm3 promoted a decreased NRA in stem tissue. These findings agree with data from previous studies (Nicholas et al., 1976; Watanabe et al., 1985; Faleiros & Cazetta, 1996), which showed that high [NO3-] could decrease enzyme activity. However, in this research, leaves did not exhibit a decrease in NRA, even when [NO3-] was increased to 100 mmol dm3. This suggests that tanner grass leaves tolerated a wider range of [NO3-] than did other plants (Nicholas et al., 1976; Faleiros & Cazetta, 1996). These results confirm that the addition of exogenous NO3- (60 mmol dm3) is necessary in order to ensure maximum NRA during an in vivo assay of tanner grass tissues. This also indicates that concentrations of about 60 mmol dm3 are sufficient to saturate the enzyme in in vivo assays for both tissues analyzed.
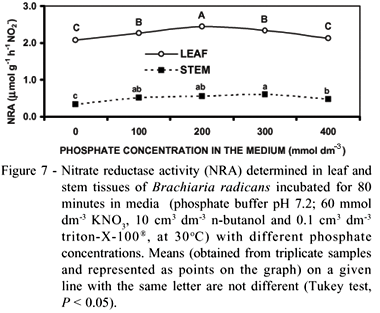
The highest values were obtained when leaf and stem tissues were incubated in media containing phosphate buffer at concentrations of 200 mmol dm3 and 300 mmol dm3, respectively (Figure 7). A significant (P < 0.05) effect of the [PO43] concentration in the medium on NRA was observed for tanner grass, but not for other species (Jones & Sheard, 1977; Nicholas et al., 1976; Streeter & Bosler, 1972).
Leaf NRA was enhanced when pH increased from 6.0 to 7.6, but dropped when pH further increased from 7.6 to 8.0. Nitrate reductase activity for pH 7.2 and 7.6 did not differ (P > 0.05), indicating that the optimum pH for the incubation medium may be between those values. For stems, probably due to the low levels of NRA in this tissue, no difference (P > 0.05) was detected for pH ranging from 6.8 to 8.0 (Figure 8), even though the behavior of NRA as a function of pH was similar to that observed for leaves. These results show similar patterns to those found for soybean (Glycine max L. Merril), beans (Phaseolus vulgaris L.), and pigeon pea (Cajanus cajan L. Millsp) (Jaworski, 1971; Nicholas et al., 1976; Puranik & Srivastava, 1983; Faleiros & Cazetta, 1996), while different values were observed for triticale (Triticosecale Wittmack) (Lin & Kao, 1980).
Both pH and buffer isotonic strength are important variables in NRA determinations (Figures 7 and 8). Despite the stems having much lower levels of NRA compared to those of leaves, the stem tissue showed the highest activity at a lower pH and higher [PO43 ], and exhibited tolerance to a wider range of both pH or buffer strength.
The incubation of tissues under vacuum conditions enhanced (P < 0.05) NRA for leaves and stems by an average of 21% and 15%, respectively, compared to the procedure without vacuum infiltration and incubation under room pressure (Figure 9). The assay showed a better performance after vacuum infiltration, with incubation maintained under vacuum conditions. Faleiros & Cazetta (1996) suggested that this is probably due to the elimination of inter-cellular gases from the tissue, which improves the contact between solution and tissue. This causes the tissue to submerge into the medium. The removal of O2 from the tissue also reduces the possibility of NO2- reduction to NH4+ in the cells, maintaining its diffusion towards the medium. Based on these results, the best in vivo assay to determine Brachiaria radicans NRA is the incubation of tissues in a medium composed of 200 mmol dm3 phosphate buffer pH 7.4, 60 mmol dm3 KNO3, 10 cm3 dm3 n-butanol, and 0.1 cm3 dm3 triton-X-100®. This should be done under vacuum and in the dark for a period of time between 60 and 100 minutes.
When tanner grass was grown in pots, it produced 1.5 to 2.5 times more stem than leaf fresh weight (Table 1). However, the average NRA per unit of fresh weight was higher (from 4.6 to 10.5 fold) in leaves, compared to the stem tissue. When NRA in the whole plant was calculated, leaf tissue had an activity from 2.1 to 6.8 times greater than stem tissue.
The treatments did affect plant development and NRA of tanner grass tissues (Table 2). Except for leaf fresh weight and NRA per unit of stem fresh weight, there were no significant N source x Mo level interactions (Table 2), indicating that N sources and levels of Mo, under this soil and experimental conditions, tended to play independent roles.
Plants fertilized with NaNO3 and urea produced more leaf fresh weight than those fertilized with NH4Cl (Table 3). No differences, however, were observed for N sources on stem fresh weight. Nitrate fertilizer tended to induce higher NRA in the leaf tissue when compared to NH4Cl and urea. The ammonium fertilizer induced slower shoot development, when compared to NaNO3 (Table 3), suggesting that tanner grass cannot detoxify NH4+, as reported for other species (Roberts & Menary, 1994).
Plants fertilized with urea tended to show intermediate growth and NRA values, despite visual similarities between NaNO3 or NH4Cl fertilized plants (Table 3). Despite the fact that urea is a source of NH4+, not all of it is immediately available, but is slowly released during its hydrolysis in the soil. The rate of urea hydrolysis in soils containing low levels of organic matter (like the one used in this research) tends to be low (Reynolds et al., 1985). Thus, NH4+ is released at such a rate that the plant can absorb NH4+ with little risk of toxicity. Also, in the moisture and oxidizing conditions under which the soil was maintained in this study, NH4+ was probably slowly oxidized to NO3- (Malavolta, 1981), providing a NO3- supply just enough to induce a slight NRA increase (Kawachi et al., 2002).
When Mo was added, plant development (leaf and stem fresh weight) was partially suppressed, as well as total NRA in leaf tissue. NRA in stem tissue showed no differences (P < 0.05), but data suggested a similar pattern as that observed for leaves (Tables 1, 2, and 3). The growth suppression imposed by Mo application may be due to partial toxicity. No injury symptoms were observed in the plants used in this study, although Gupta (2001) stated that symptoms become apparent only under very high Mo levels.
When Mo was applied in the absence of N (control), it improved NRA, but stalk development tended to be suppressed (Tables 2 and 3). Leaf fresh weigh tended to show similar results, although the differences did not reach significant levels (P < 0.05) (Tables 2 and 3). It seems that Mo improved NRA because it is a NR cofactor; however, the rate of Mo applied to the soil in this research probably was too high for this species, and may explain the partial suppression of plant development.
The addition of Mo tended to reduce leaf fresh weight of plants under NaNO3 and urea fertilization (Tables 2 and 4 ). Also, urea promoted better leaf growth than NH4Cl when Mo was not applied, but did not differ under Mo fertilization (Table 4 ). Sources of N did not affect stem NRA when plants were grown without Mo fertilization (Table 5). However, for Mo fertilized plants, NaNO3 induced higher NRA when compared to NH4Cl, while urea produced intermediate values. On the other hand, the effect on NRA was increased by NaNO3, decreased by NH4Cl, and was not altered by urea under Mo fertilization.
These findings together suggest that in this research the limitation imposed by Mo application would not be directly related to NR, but involved toxicity in other ways.
In summary, the objective of the NRA assay optimization was met. Tanner grass plants produced much more stem fresh weight than leaf fresh weight, with most NRA found in the leaves. Although it would be expected that, to some extent, tanner grass plants could avoid NO3- reduction by absorbing reduced N directly from fertilizers, NH4Cl and urea induced less plant development when compared to NaNO3, showing that this species has a lower tolerance to these sources of N, mainly NH4Cl. The application of 20 kg ha-1 of Mo as H2Mo2O4 to the soil under the conditions used in this research, affected plant development and NRA to some extent; however, it seems that the reduced plant growth observed did not directly involve NRA.
Further research is needed to unveil the plant toxicity mechanism triggered by high levels of Mo in the soil, and how it effects N utilization. Experiments consisting of combinations of different NO3- and Mo levels might help clarify and better understand this matter.
ACKNOWLEDGEMENTS
This study was supported in part by a scholarship awarded to L.C.V. Villela by FAPESP.
Received Fevruary 14, 2003
Accepted September 13, 2004
- ASLAM, M.; HUKFFAKER, R.C.; RAINS, D.W. Early effects of salinity on nitrate assimilation in barley seedlings. Plant Physiology, v.76, p.321-325, 1984.
- BAER, G.R.; COLLET, G.F. In vivo determination of parameters of nitrate utilization in wheat (Triticum aestivum L.) seedling grown with low concentration of nitrate in the nutrient solution. Plant Physiology, v.68, p.1237-1243, 1981.
- BARTHES, L.; DELEENS, E.; BOUSSER, A.; HOARAU, J.; PRIOL, J.L. Xylem exudation is related to nitrate assimilation pathway in developing maize seedlings: Use of nitrate reductase and glutamine synthetase inhibitors as tools. Journal of Experimental Botany, v.47, p.485-495, 1996.
- BLONDEL, A.M.; BLANC, D. Mise au point dúne methode de mesure in vivo de lactivité de la nitrate reductase. Annals of Agronomy, v.26, p.309-322, 1975.
- BRUNETTI, N.; HAGEMAN, R.H. Comparison of in vivo and in vitro assays of nitrate reductase in wheat (Triticum aestivum L.) seedlings. Plant Physiology, v.58, p.583-587, 1976.
- DORDAS, C.; SAH, R.; BROWN, P.H.; ZENG, Q.; HU, H. Remobilização de micronutrientes e elementos tóxicos em plantas superiores. In: FERREIRA, M.E.; CRUZ, M.C.P.; RAIJ, B.V.; ABREU, C.A. (Ed.) Micronutrientes e elementos tóxicos na agricultura Jaboticabal: CNPq; FAPESP; Potafos, 2001. cap.3, p.43-61.
- FALEIROS, R.R.S.; CAZETTA, J.O. Optimization of the in vivo assay for nitrate reductase activity determination in pigeon pea [Cajanus cajan (L.) Millsp] leaves. Científica, v.24, p.385-396, 1996.
- GUPTA, U.C. Micronutrientes e elementos tóxicos em plantas e animais. In: FERREIRA, M.E.; CRUZ, M.C.P.; RAIJ, B.V.; ABREU, C.A. (Ed.) Micronutrientes e elementos tóxicos na agricultura Jaboticabal: CNPq; FAPESP; Potafos, 2001. cap.2, p.13-42.
- HARPER, J.E. Evolution of nitrogen oxide(s) during in vivo nitrate reductase assay of soybean leaves. Plant Physiology, v.68, p.1488-1493, 1981.
- HEUER, B.; PLAUT, Z. Reassessment of the in vivo assay for nitrate reductase in leaves. Physiologia Plantarum, v.43, p.306-312, 1978.
- JAWORSKI, E.K. Nitrate reductase assay in intact plant tissues. Biochemical and Biophysical Research Communications, v.43, p.1274-1279, 1971.
- JONES, R.W.; SHEARD, R.W. Conditions affecting in vivo nitrate reductase activity in chlorophyllous tissues. Canadian Journal of Botany, v.55, p.896-901, 1977.
- KAWACHI, T.; SHOJI, Y.; SUGIMOTO, T.; OJI, Y.L.; KLEINHOFS, A.; WARNER, R.L.; OHTAKE, N.; OHYAMA, T.; SUEIOSHI, K. Role of xylem sap nitrate in regulation of nitrate reductase gene expression in leaves of barley (Hordeum vulgare L.) seedlings. Soil Science and Plant Nutrition, v.48, p.79-85, 2002.
- KLEPPER, L.; FLESHER, D.; HAGEMAN, R.H. Generation of reduced Nicotinamide Adenine Dinucleotide for nitrate reduction in green leaves. Plant Physiology, v.48, p.580-590, 1971.
- LAWRENCE, J.M.; HERRICK, H.E. Media for in vivo nitrate reductase assay of plant tissues. Plant Science Letters, v.24, p.17-26, 1982.
- LEA, P.J. Primary nitrogen metabolism. In: DAY, P.M.; J.B. HARBORN (Ed.) Plant biochemistry New York: Academic Press, 1997. cap.7, p.273-313.
- LILLO, C. Studies of diurnal variations of nitrate reductase activity in barley leaves using various assay methods. Physiologia Plantarum, v.57, p.357-362, 1983.
- LIN, W.H.; KAO, C.H. Factors affecting nitrate reductase activity in triticale. Physiologia Plantarum, v.48, p.361-364, 1980.
- MALAVOLTA, E. Elementos de nutrição mineral de plantas São Paulo: Ed. Agronômica Ceres, 1981.
- MITIDIERI, J. Manual de gramíneas e leguminosas para pastos tropicais São Paulo: EDUSP, 1983.
- MULLER, E.K.H.; JANIESHCH, P. In vivo nitrate reductase activity in Carex pseudocyperus L.: the influence of nitrate-ammonium concentration ratios and correlation with growth. Journal of Plant Nutrition, v.16, p.1357-1372, 1993.
- NICHOLAS, J.C.; HARPER, J.E.; HAGEMAN, R.H. Nitrate reductase activity in soybeans (Glycine max [L] Merr.): 1. Effects of light and temperature. Plant Physiology, v.58, p.731-735, 1976.
- PURANIK, R.M.; SRIVASTAVA, H.S. Increase in nitrate reductase activity in the presence of sucrose in bean leaf segments. Phytochemistry, v.22, p.2383-2387, 1983.
- REYNOLDS, C.M.; WOLF, D.C.; ARMBRUSTER, J.A. Factors related to urea hydrolysis in soils. Soil Science Society of America Journal, v.49, p.104-108, 1985.
- ROBERTS, N.J.; MENARY, R.C. Effect of nitrogen on growth, flower yield, oil composition, and yield in Boronia megastigma Nees. Journal of Plant Nutrition, v.17, p.2035-2052, 1994.
- SKERMAN, P.J.; RIVEROS, F. Tropical grasses Rome: FAO, 1990. 254p. (FAO (Plant Production and Protection Series, 23).
- STITT, M.; MÜLLER, C.; MATT, P.; GIBSON, Y.; CARILLO, P.; MORCUENDE, R.; SCHEIBLE, W.; KRAPP, A. Steps towards an integrated view of nitrogen metabolism. Journal of Experimental Botany, v.53, p.959-970, 2002.
- SOUZA, F.H.D. Brachiaria spp in Brazil. In: LOCH, D.S.; FERGUSSON, J.E. (Ed.) Forage seed production: tropical and subtropical species. New York: CAB International Publishing, 1999. v.2, p.371-379.
- SRIVASTAVA, H.S. Regulation of nitrate reductase activity in higher plants. Phytochemistry, v.19, p.725-733, 1990.
- STOLTZ, J.F.; BAZU, P. Evolution of nitrate reductase: Molecular and structural variations on a common function. Chembiochem, v.3, p.198-206, 2002.
- STREETER, J.G.; BOSLER, M.E. Comparison of in vitro and in vivo assays for nitrate reductase in soybeans leaves. Plant Physiology, v.49, p.448-450, 1972.
- TINGEY, D.T.; FITES, R.C.; BAHARSJAH, J. Factors influencing nitrate reductase in soybean foliage. New Phytologyst, v.73, p.21-29, 1974.
- VIÉGAS, R.A.; MELLO, A.R.B.; SILVEIRA, J.A.G. Nitrate reductase activity and proline accumulation in cashew in response to NaCl salt shock. Revista Brasileira de Fisiologia Vegetal, v.11, p.21-28, 1999.
- WATANABE, M.; HAYASHI, M.; SUGIYAMA, T. Effects of supplemental nitrate application on the activity of some nitrogen assimilation enzymes and leaf tissue productivity in maize seedlings. Soil Science and Plant Nutrition, v.31, p.573-580, 1985.

Publication Dates
-
Publication in this collection
29 Nov 2004 -
Date of issue
Dec 2004
History
-
Received
14 Feb 2003 -
Accepted
13 Sept 2004