Abstracts
Guava (Psidium guajava L.) is a tropical fruit that presents fast post-harvest ripening; therefore it is a very perishable product. Inappropriate storage temperature and retail practices can accelerate fruit quality loss. The objective of this study was to evaluate the respiratory activity (RA), the ethylene production (EP) and Q10 of guava fruit at different storage temperatures. 'Paluma' guava fruits were harvested at maturity stage 1 (dark-green skin) and stored at either 1, 11, 21, 31 or 41ºC; RA and EP were determined after 12, 36, 84 and 156 h of storage. RA and EP rates at 1 and 11ºC were the lowest - 0.16 and 0.43 mmol CO2 kg-1 h-1 and 0.003 and 0.019 µmol C2H4 kg-1 h-1, respectively. When guavas were stored at 21ºC, a gradual increase occurred in RA and EP, reaching 2.24 mmol CO2 kg-1 h-1 and 0.20 µmol C2H4 kg-1 h-1, after 156 h of storage. The highest RA and EP were recorded for guavas stored at 31ºC. In spite of high RA, guavas stored at 41ºC presented EP similar to guavas stored at 11ºC, an indicator of heat-stress injury. Considering the 1-11ºC range, the mean Q10 value was around 3.0; the Q10 value almost duplicated at 11-21ºC range (5.9). At 21-31ºC and 31-41ºC, Q10 was 1.5 and 0.8, respectively. Knowing Q10, respiratory variation and ripening behavior in response to different temperatures, fruit storage and retail conditions can be optimized to reduce quality losses.
Psidium guajava; ethylene; ripening
A goiaba (Psidium guajava L.) é um fruto tropical que apresenta rápido amadurecimento, o que a torna um produto muito perecível. Temperaturas inapropriadas durante o armazenamento e comercialização podem acelerar a perda da qualidade dos frutos. O objetivo desse trabalho foi avaliar a atividade respiratória (AR), a produção de etileno (PE) e o coeficiente Q10 de goiabas em diferentes temperaturas de armazenamento. Goiabas do cultivar Paluma foram colhidas no estádio 1 de maturação (casca verde escura) e armazenadas a 1, 11, 21, 31 e 41ºC. A AR e a PE foram determinadas com 12, 36, 84 e 156 h de armazenamento. As taxas de AR e PE a 1 e 11ºC foram as menores, atingindo valores ao redor de 0,16 e 0,43 mmol CO2 kg-1 h-1 e 0,003 e 0,019 µmol C2H4 kg-1 h-1, respectivamente. Quando as goiabas foram armazenadas a 21ºC, observou-se aumento gradual em AR e PE, as quais alcançaram valores de 2,24 mmol CO2 kg-1 h-1 e 0,20 µmol C2H4 kg-1 h-1 após 156 h de armazenamento. As maiores AR e PE foram observadas em goiabas armazenadas a 31ºC. Apesar de alta AR, goiabas armazenadas a 41ºC tiveram baixa PE, similarmente àquelas armazenadas a 11ºC, indicando dano por alta temperatura. Na faixa de 1-11ºC, o valor médio de Q10 foi de 3,0, enquanto esse valor quase duplicou na faixa de 11-21ºC, atingindo 5,9. O Q10 decresceu nas faixas de 21-31ºC e 31-41ºC, apresentando valores de 1,5 e 0,8, respectivamente. Conhecendo-se a variação do Q10, da taxa respiratória e do comportamento do amadurecimento em resposta a diferentes temperaturas, as condições de armazenamento e comercialização dos frutos podem ser otimizadas para reduzir as perdas na qualidade.
Psidium guajava; amadurecimento; etileno
CROP SCIENCE
Temperature-related changes in respiration and Q10 coefficient of Guava
Mudanças na respiração e no coeficiente Q10 de goiaba relacionadas à temperatura
Ilana Urbano BronI, * * Corresponding author < ilana@iac.sp.gov.br> ; Rafael Vasconcelos RibeiroII; Flávia Cristina CavaliniIII; Angelo Pedro JacominoIII; Marcos José TrevisanIII
IIAC - Centro APTA de Engenharia e Automação, C.P. 26 - 13201-970 - Jundiaí, SP - Brasil
IIIAC - Centro de Pesquisa e Desenvolvimento de Ecofisiologia e Biofísica, C.P. 28 - 13001-970 - Campinas, SP - Brasil
IIIUSP/ESALQ - Depto. de Produção Vegetal - Lab. de Pós-colheita de Produtos Hortícolas, C.P. 09 - 13418-900 - Piracicaba, SP - Brasil
ABSTRACT
Guava (Psidium guajava L.) is a tropical fruit that presents fast post-harvest ripening; therefore it is a very perishable product. Inappropriate storage temperature and retail practices can accelerate fruit quality loss. The objective of this study was to evaluate the respiratory activity (RA), the ethylene production (EP) and Q10 of guava fruit at different storage temperatures. 'Paluma' guava fruits were harvested at maturity stage 1 (dark-green skin) and stored at either 1, 11, 21, 31 or 41ºC; RA and EP were determined after 12, 36, 84 and 156 h of storage. RA and EP rates at 1 and 11ºC were the lowest - 0.16 and 0.43 mmol CO2 kg-1 h-1 and 0.003 and 0.019 µmol C2H4 kg-1 h-1, respectively. When guavas were stored at 21ºC, a gradual increase occurred in RA and EP, reaching 2.24 mmol CO2 kg-1 h-1 and 0.20 µmol C2H4 kg-1 h-1, after 156 h of storage. The highest RA and EP were recorded for guavas stored at 31ºC. In spite of high RA, guavas stored at 41ºC presented EP similar to guavas stored at 11ºC, an indicator of heat-stress injury. Considering the 1-11ºC range, the mean Q10 value was around 3.0; the Q10 value almost duplicated at 11-21ºC range (5.9). At 21-31ºC and 31-41ºC, Q10 was 1.5 and 0.8, respectively. Knowing Q10, respiratory variation and ripening behavior in response to different temperatures, fruit storage and retail conditions can be optimized to reduce quality losses.
Key words:Psidium guajava, ethylene, ripening
RESUMO
A goiaba (Psidium guajava L.) é um fruto tropical que apresenta rápido amadurecimento, o que a torna um produto muito perecível. Temperaturas inapropriadas durante o armazenamento e comercialização podem acelerar a perda da qualidade dos frutos. O objetivo desse trabalho foi avaliar a atividade respiratória (AR), a produção de etileno (PE) e o coeficiente Q10 de goiabas em diferentes temperaturas de armazenamento. Goiabas do cultivar Paluma foram colhidas no estádio 1 de maturação (casca verde escura) e armazenadas a 1, 11, 21, 31 e 41ºC. A AR e a PE foram determinadas com 12, 36, 84 e 156 h de armazenamento. As taxas de AR e PE a 1 e 11ºC foram as menores, atingindo valores ao redor de 0,16 e 0,43 mmol CO2 kg-1 h-1 e 0,003 e 0,019 µmol C2H4 kg-1 h-1, respectivamente. Quando as goiabas foram armazenadas a 21ºC, observou-se aumento gradual em AR e PE, as quais alcançaram valores de 2,24 mmol CO2 kg-1 h-1 e 0,20 µmol C2H4 kg-1 h-1 após 156 h de armazenamento. As maiores AR e PE foram observadas em goiabas armazenadas a 31ºC. Apesar de alta AR, goiabas armazenadas a 41ºC tiveram baixa PE, similarmente àquelas armazenadas a 11ºC, indicando dano por alta temperatura. Na faixa de 1-11ºC, o valor médio de Q10 foi de 3,0, enquanto esse valor quase duplicou na faixa de 11-21ºC, atingindo 5,9. O Q10 decresceu nas faixas de 21-31ºC e 31-41ºC, apresentando valores de 1,5 e 0,8, respectivamente. Conhecendo-se a variação do Q10, da taxa respiratória e do comportamento do amadurecimento em resposta a diferentes temperaturas, as condições de armazenamento e comercialização dos frutos podem ser otimizadas para reduzir as perdas na qualidade.
Palavras-chave:Psidium guajava, amadurecimento, etileno
INTRODUCTION
Temperature heavily influences metabolic activity of plant tissues and organs, such as fruit (Chitarra & Chitarra, 1990). Metabolic reactions, such as respiration and ethylene production, are fundamental for fruit ripening, but harmful to fruit conservation. These reactions ordinarily increase with increasing temperature up to 40ºC, when ethylene biosynthesis is impaired by heat stress (Eaks, 1978). On the other hand, enzymatic reactions occur more slowly at low temperatures, extending fruit shelf life (Chitarra & Chitarra, 1990). Low temperatures can cause chilling injuries, but the threshold temperature is specific for each species and depends on the ripening stage (Wang, 1982).
The Q10 coefficient commonly used in postharvest studies regarding the fruit respiratory activity, represents the increase in the rate of a process with a 10ºC increase in temperature. Enzymatic and physiological processes are twice to three times faster for each 10ºC of temperature increase. However, the range of Q10 can go from 1 to 10 or more (Ting, 1982).
The increase or reduction in fruit respiration can vary with the exposure to temperature. Kader (1985) and Kluge et al. (2002) have reported that Q10 values of some fruits change as function of the considered temperature range. The recommended storage temperature for guava fruit (Psidium guajava L.) varies from 8 to 10ºC (Carraro & Cunha, 1994; Castro & Sigrist, 1988). In typical, tropical Brazilian climate, guava fruit can easily be exposed to temperatures higher than 10ºC during storage and commercialization period, and undergo physiological stress and loss of shelf life and quality. The objective of this study was to evaluate the respiratory activity, ethylene production and Q10 of Paluma guava cultivar at different storage temperatures.
MATERIAL AND METHODS
Studied material - 'Paluma' guava fruits (Psidium guajava L.) were harvested at maturity stage 1 in a commercial orchard in Vista Alegre do Alto County, SP Brazil (21º10'S; 48º38'W; 700 m) and transported in a refrigerated truck at 15ºC during 4 hours to Piracicaba, SP Brazil. The maturity stage was defined by skin color as a fruit with dark-green skin (Azzolini et al., 2004).
Temperature treatments - Seven guava fruits were stored in each temperature-controlled chamber at 1, 11, 21, 31 and 41ºC during 156 h.
Respiration and ethylene measurements - Respiration rate and ethylene production were determined after 12, 36, 84 and 156 h of storage in each temperature. Guava fruits were enclosed in a 0.45 L hermetic flask during 1 h, then gas samples of 0.001 L were collected from flasks with a gas-tight syringe through a silicone septum, and analyzed in a gas chromatographer model Trace 2000GC equipped with a capillary Porapack, 2 m column set at 100ºC with hydrogen as carrier gas, pressure 105 Pa. Respiration rate and ethylene production were determined by measuring the difference between the initial (when flasks were closed) and the final gas concentration (after 1 h), and expressed as mmol CO2 kg-1 h-1 and mmol C2H4 kg-1 h-1, respectively. Q10 values for respiration were calculated as the quotient between respiration rates (RR) measured at two different temperatures as follows:

The Q10 was calculated considering the temperature of fruit surface in each measurement. The fruit temperature (FT) was measured with an infrared thermometer model 4000-4GL positioned at 5 cm from fruit surface and attached to a micrologger model CR23X. In addition, log (RR) was plotted against FT: log(RR)=a*FT+b, being Q10 calculated from the a coefficient: Q10=10(10*a) (Benkeblia et al., 2000).
After 156 h of storage, guava fruit stored at 1 and 11ºC were transferred to 25ºC for ripening visual observation. Weight loss was determined by weighing individual fruit before respiration and ethylene analyses. The seven guava fruit stored in each temperature were weighed after 12, 36, 84 and 156 h of storage.
The experiment was arranged in random block design (n = 7). Data were analyzed using the ANOVA procedure and the Tukey test (a = 0.01; 0.05) was used to compare means.
RESULTS AND DISCUSSION
Respiratory rates of guavas stored at 1 and 11ºC were the lowest (Figure 1a). The respiration in guavas stored at 1ºC increased in the first 36 h, reaching a stable level between 36 and 84 h, and increasing again after 156 h of storage, reaching 0.16 mmol CO2 kg-1 h-1. After an initial increase in the respiratory rate, fruit stored at 11ºC presented stable respiration until the end of storage period, values varying around 0.4 mmol CO2 kg-1 h-1.

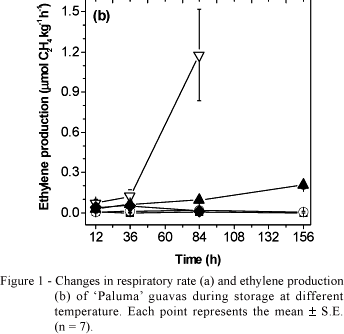
Ethylene production in guavas stored at 1 and 11ºC was very low - 0.003 and 0.019 µmol C2H4 kg-1 h-1 after 84 h of storage, respectively, decreasing to 0.001 and 0.007 µmol C2H4 kg-1 h-1 thereafter, respectively (Figure 1b). Although low ethylene levels were measured in fruit stored at those temperatures, this plant gaseous hormone is biologically active in trace amounts, being active in concentrations of 0.01 µL L-1 (Abeles et al., 1992). Since guavas stored at 1 and 11ºC produced very low ethylene, there was no progress of ripening during storage, as confirmed by visual evaluation after 156 h of storage (Figure 2).
In fact, guavas stored at 1 and 11ºC exhibited green skin with non-significant alteration of pulp color (Figure 2). Even after 4 days at 25ºC, those fruits did not develop the typical, ripe-fruit skin and pulp color (data not shown). Since 11ºC is the recommended storage temperature, it was expected that guavas stored at this condition would complete ripening when transferred to higher temperatures. However the fruits did not ripen, possibly because of the maturity stage in which they were harvested. This can indicate a physiological disturbance caused by low temperature exposure on underlying processes that trigger fruit ripening. Prasanna et al. (2000) reported that pulp color, texture, taste and flavor of ripe custard apples held at 25 and 20ºC were superior when compared with those stored at 10ºC, which did not normally ripe.
The failure in ripening is a common symptom of chilling injury in tropical fruit (Couey, 1982). Temperatures below 10°C are generally responsible for cold damages in chilling-sensitive tropical fruits, as guavas. As a matter of fact, Osman & Ayub (1998) verified that guavas stored at 3°C did not ripen satisfactorily, and that was related to chilling injury. According to Kader (1985), the ideal storage temperature for those fruits can vary from 10 to 15°C, depending on species and maturity stage. Physical change of membranes, from a flexible, liquid-crystalline to a solid-gel structure, is the primary response in chilled fruit. Secondary responses include decrease in the rate of mitochondrial oxidative activity as well as increasing activation energy of membrane-associated enzymes until a complete disorganization of cellular structure (Wang, 1982).
When guavas were stored at 21ºC, the respiratory activity and ethylene production gradually increased, reaching 2.24 mmol CO2 kg-1 h-1 and 0.20 µmol C2H4 kg-1 h-1 after 156 h of storage respectively, (Figure 1). Cavalini (2004) also observed gradual increase in respiration and ethylene production in 'Paluma' guavas harvested at maturity stage 2 (light-green skin) and stored at 25ºC, rates being similar to those reported herein (Figure 1). After 156 h of storage at 21ºC, guavas were visually ripe, exhibiting a complete yellow skin and red pulp (Figure 2).
The results are in accordance to those reported by Akamine & Goo (1979) for Psidium cultivars, reaching maximum mean values around 75 mL CO2 kg-1 h-1 (3 mmol CO2 kg-1 h-1) and 4 µL C2H4 kg-1 h-1 (0.16 µmol C2H4 kg-1 h-1). However, the fruit did not show the climacteric behavior pattern found by Akamine & Goo (1979) in any storage temperature. Guava fruit had same initial quality, and were all at the same maturity stage, i.e. dark-green stage (Azzolini et al., 2004), which excludes those influences on ripening behavior of 'Paluma' guava fruit. Therefore, the climacteric or non-climacteric behavior of guava fruit should be clarified in further studies when an intermediate ripening behavior could be proposed, based on a systemic evaluation of postharvest fruit characteristics in addition to respiration and ethylene production.
The highest respiratory rates were recorded for guavas stored at 31 and 41ºC, but respiration rates were always comparatively lower in guavas stored at 41ºC (Figure 1a). Ethylene production of guavas stored at 41ºC was very low (Figure 1b). In the first 36 h of storage, the ethylene production showed increasing trend, reaching 0.053 µmol C2H4 kg-1 h-1. However, after 84 h storage, production of ethylene decreased to 0.010 µmol C2H4 kg-1 h-1 a pattern similar to that observed for guavas stored at 11ºC (Figure 1b). Similar results were observed for kiwi (Antunes & Sfakiotakis, 2000) and apple (Lurie & Klein, 1990), in which high temperature stress decreased ripening and ethylene production, increasing respiration rate.
In spite of high respiratory rates, guavas stored at 31 and 41ºC did not show normal ripening during storage period, retaining greener peel and light pulp color, in comparison to guavas stored at 21ºC (Figure 2). High temperatures tend to disrupt physiological processes by thermal denaturation of enzymes, and perhaps alteration of important cellular and sub-cellular structures (Ting, 1982). According to Paull & Chen (2000), cell wall degrading enzymes and ethylene production are frequently the most disrupted processes. Failure of fruit to ripe at high temperatures has been attributed to the reduction of ethylene biosynthesis (Eaks, 1978).
The stress resulting from high temperature appears to inhibit 1-aminocyclopropane-1-carboxylate oxidase (ACC oxidase) more than 1-aminocyclopropane-1-carboxylate synthase (ACC synthase) (Yu et al., 1980). Apelbaum et al. (1981) proposed that high temperatures cause impairment of ethylene production by disturbing cellular membranes, resulting in inhibition of the membrane-associated ACC oxidase. In apple and tomato, Yu et al. (1980) and Atta Aly (1992) reported that temperatures of 35-38°C caused endogenous ACC to accumulate concomitantly with the decrease in ethylene. Therefore, the lower ethylene production of guavas stored at 41ºC was probably caused by heat-damage, that is, this temperature exceeds the threshold temperature for heat injury.
Fruits stored at 31ºC and 41ºC presented incidence of anthracnosis, a disease caused by Colletotrichum gloeosporioides (Penz) Penz & Sacc. (Piccinin & Pascholati, 1997). The highest incidence of this disease in guavas stored at 31ºC can be explained by the fact that 30ºC is the optimum temperature for the development of Colletotricum, (Piccinin & Pascholati, 1997). Therefore, the high ethylene production (1.17 µmol C2H4 kg-1 h-1), recorded at 84 h after storing guavas at 31ºC (Figure 1b), was probably caused by fungi presence. To avoid the interference of fungus respiration, fruit respiration and ethylene production were not measured after 84 h both at 31 and 41ºC.
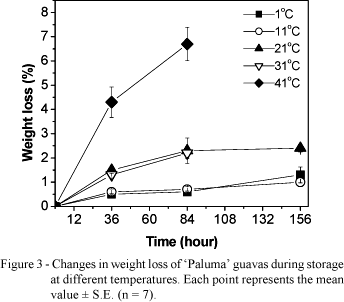
Guavas stored at 1ºC and 11ºC as well as those stored at 21ºC and 31ºC had 1.3% and 2.4% weight loss, respectively (Figure 3). These losses were very low and did not cause any visual depreciation. On the other hand guavas stored at 41ºC had 6.7% weight loss, after 84 h. In fact, turning visually smaller than fruits of the other groups after 156 h storage (Figure 2).
Regarding Q10 values, significant changes were related to temperature range (Table 1). At the 1-11ºC range, mean Q10 = 3.0; at 11-21ºC range this value almost duplicated, (Q10 = 5.9). The increase of respiration rate depends on temperature range, being the highest Q10 values found at 11-21ºC range (P < 0.01) (Table 1). Considering that respiration increases two to three times for every 10ºC in temperature rise (Ting, 1982), discretion is needed when referring to guava fruit. Only at 1-11ºC range the Q10 values lied between two and three. One possible consequence of misleading Q10 values would be the reduction in shelf life conservation, once the deterioration rate would be twice as fast as expected.
At 11-21ºC, Q10 = 5.9. It is thus possible to foresee that the respiratory rate of guavas stored, as recommended (i.e. around 11ºC), would increases 6 times when transferring fruits to higher temperatures (i.e. around 21ºC), similar to those found at market stands. This fact helps to understand why guava is considered a very perishable fruit. At higher temperature ranges - 21-31ºC and 31-41ºC - Q10 values decreased to 1.5 and 0.8, respectively (Table 1). Occurrence of physiological damage at 41ºC is evidenced by records of Q10 smaller than those observed at 31-41ºC range.
According to Larcher (2000), Q10 values are high at lower temperatures because in such conditions biochemical reaction are limited by low enzymatic activity. At high temperatures, the Q10 values are low because in such conditions physical processes, e.g. gaseous diffusion, limit the speed of the reactions. Regarding Q10 values within each temperature range, there were no significant (P < 0.05) changes during the storage period, except at 11-21ºC range (Table 1). In this condition, the highest Q10 values were observed after 12 h of storage.
Benkeblia et al. (2000) suggested respiration rates increase linearly with increasing temperature (dotted lines in Figure 4). However, this behavior did not happen when fruit was damaged by heating, as occurred at 41ºC (Figure 4). This fact probably happens when broad temperature ranges (i.e. 40ºC) are evaluated. The estimation of Q10, by the linear regression approach leads to lower values in comparison to the mean Q10 value of each evaluation time (Table 1 and Figure 4). Also higher standard errors were observed when Q10 was calculated by the linear regression technique (Table 1 and Figure 4).
Once the visual damage (i.e. fruit did not ripe when transferred to room temperature) was noticed in guavas stored at 11ºC and 1ºC after 156 h of storage, it is fain to infer that 'Paluma' guava, harvested in stage 1, can be stored at temperatures between 11 and 21ºC instead of the general guava recommendation of storage between 8 and 10ºC (Carraro & Cunha, 1994; Castro & Sigrist, 1988). However, further studies are needed to define the optimum temperature to refrigerated conservation of guava fruit, which probably varies according to maturity stage, cultivar, and storage time. Knowing Q10, respiratory variation and ripening behavior in response to different temperatures, fruit storage and retail conditions can be optimized to reduce quality losses.
Received April 08, 2005
Accepted July 19, 2005
- ABELES, F.B.; MORGAN, P.W.; SALTVEIT, M.E. Ethylene in plant biology San Diego: Academic Press, 1992. 414p.
- AKAMINE, E.K.; GOO, T. Respiration and ethylene production in fruits of species and cultivars of Psidium and species of Eugenia Journal of the American Society for Horticultural Science, v.104, p.632-635, 1979.
- ANTUNES, M.D.C.; SFAKIOTAKIS, E.M. Effect of high temperature stress on ethylene biosynthesis, respiration and ripening of 'Hayward' kiwifruit. Postharvest Biology and Technology, v.20, p.251-259, 2000.
- APELBAUM, A.; BURGOON, A.C.; ANDERSON, J.D.; SOLOMOS, T.; LIEBERMAN, M. Some characteristics of the system converting 1-aminocyclopropane-1-carboxylic acid to ethylene. Plant Physiology, v.67, p.80-84, 1981.
- ATTA Aly, M.A. Effect of high temperature on ethylene biosynthesis by tomato fruit. Postharvest Biology and Technology, v.2, p.19-24, 1992.
- AZZOLINI, M.; JACOMINO, A.P.; BRON, I.U. Índices para avaliar qualidade pós-colheita de goiabas em diferentes estádios de maturação. Pesquisa Agropecuária Brasileira, v.39, p.139-145, 2004.
- BENKEBLIA, N.; VAROQUAUX, P.; GOUBLE, B.; SELSELET-ATTOU, G. Respiratory parameters of onion bulbs (Allium cepa) during storage. Effects of ionizing radiation and temperature. Journal of the Science of Food and Agriculture, v.80, p.1772-1778, 2000.
- CARRARO, A.F.; CUNHA, M.M. Manual de exportação de frutas Brasília: Iica, 1994. 252p.
- CASTRO, J.V.; SIGRIST, J.M.M. Matéria-prima. In: MEDINA, J.C. (Ed.) Goiaba: cultura, materia-prima, processamento e aspectos econômicos. Campinas: ITAL, 1988. cap.2, p.121-140.
- CAVALINI, F.C. Índices de maturação, ponto de colheita e padrão respiratório de goiabas 'Kumagai' e 'Paluma'. Piracicaba: USP/ESALQ, 2004. 69p. (Dissertação - Mestrado).
- CHITARRA, M.I.F.; CHITARRA, A.B. Pós-colheita de frutos e hortaliças: fisiologia e manuseio. Lavras: ESAL/FAEPE, 1990. 360p.
- COUEY, H.M. Chilling injury of crops of tropical and subtropical origin. HortScience, v.17, p.162-165, 1982.
- EAKS, I.L. Ripening, respiration, and ethylene production of 'Hass' avocado fruit at 20 to 40°C. Journal of the American Society for Horticultural Science, v.103, p.576-578, 1978.
- KADER, A.A. Postharvest biology and technology: an overview. In: KADER, A.A.; KASMIRE, R.F.; MITCHELL, F.G.; REID, M.S.; SOMMER, N.F.; THOMPSON, J.F. (Ed.) Postharvest technology of horticultural crops Davis: University of California, 1985. cap.1, p.3-7.
- KLUGE, R.A.; NACHTIGAL, J.C.; FACHINELLO, J.C.; BILHALVA, A.B. Fisiologia e manejo pós-colheita de frutas de clima temperado Campinas: Livraria e Editora Rural, 2002. 214p.
- LARCHER, W. O balanço de carbono das plantas. In: LARCHER, W. Ecofisiologia vegetal São Carlos: RIMA Artes e Textos, 2000. cap.2, p.69-182.
- LURIE, S.; KLEIN, J.D. Heat treatment of ripening apples: differential effects on physiology and biochemistry. Physiologia Plantarum, v.78, p.181-186, 1990.
- OSMAN, A.; AYUB, M.N.A. Effects of different postharvest treatments on the respiration patterns of guava (Psidium guajava L.). Acta Horticulturae, n.464, p.502, 1998.
- PAULL, R.E.; CHEN, N.J. Heat treatments and fruit ripening. Postharvest Biology and Technology, v.21, p.21-37, 2000.
- PICCININ, E.; PASCHOLATI, S.F. Doenças da goiabeira (Psidium guajava). In: KIMATI, H.; AMORIM, L.; BERGAMIN FILHO, A.; CAMARGO, L.E.A.; REZENDE, J.A.M. (Ed.) Manual de fitopatologia: doenças das plantas cultivadas. São Paulo: Editora Agronômica Ceres, 1997. cap.41, p.450-455.
- PRASANNA, K.N.V.; RAO, D.V.S.; KRISHNAMURTHY, S. Effect of storage temperature on ripening and quality of custard apple (Annona squamosa L.) fruits. Journal of Horticultural Science and Biotechnology, v.75, p.546-550, 2000.
- TING, I.P. The energy budget of plants. In: TING, I.P. Plant physiology Reading: Addison-Wesley Publishing Company, 1982. cap.7, p.170-188.
- WANG, C.Y. Physiological and biochemical responses of plants to chilling stress. HortScience, v.17, p.173-186, 1982.
- YU, Y.B.; ADAMS, D.O.; YANG, S.F. Inhibition of ethylene production by 2,4 dinitrophenol and high temperature. Plant Physiology, v.66, p.286-290, 1980.
Publication Dates
-
Publication in this collection
26 Sept 2005 -
Date of issue
Oct 2005
History
-
Accepted
19 July 2005 -
Received
08 Apr 2005