Abstracts
The nutrient accumulation curves of apple trees are good indicators of plant nutrient demand for each developmental stage. They are also a useful tool to evaluate orchard nutritional status and to estimate the amount of soil nutrient removal. This research aimed at evaluating the seasonality of nutrients in commercial apple orchards during the agricultural years of 1999, 2000, and 2001. Therefore, apple tree leaves and fruits of three cultivars 'Gala', 'Golden Delicious' and 'Fuji' were weekly collected and evaluated for fresh and dry matter, fruit diameter and macronutrient (N, P, K, Ca and Mg) and micronutrient (B, Cu, Fe, Mn, and Zn) concentrations. Leaf and fruit sampling started one or two weeks after full bloom, depending on the cultivar, and ended at fruit harvest or four weeks later (in the case of leaf sampling). In general, leaf concentrations of N, P, K, Cu, and B decreased; Ca increased; and Mg, Fe, Mn, and Zn did vary significantly along the plant vegetative cycle. In fruits, the initial nutrient concentrations decreased quickly, undergoing slow and continuous decreases and then remaining almost constant until the end of fruit maturation, indicating nutrient dilution, once the total nutrient accumulation increased gradually with fruit growth. Potassium was the nutrient present in highest quantities in apple tree fruits and thus, the most removed from the soil.
Malus domestica; macronutrients; micronutrients; accumulation
As curvas de acúmulo de nutrientes pela macieira têm se mostrado como um bom parâmetro indicador da demanda da cultura em cada etapa de desenvolvimento, sendo importante para avaliar o estado nutricional do pomar e estimar a remoção de nutrientes do solo. O objetivo do presente trabalho foi avaliar a sazonalidade de nutrientes na macieira, em pomares comerciais, durante as safras agrícolas de 1999, 2000 e 2001. Para tanto, amostras de folhas e frutos de três cultivares 'Gala', 'Golden Delicious' and 'Fuji' foram coletadas semanalmente e avaliadas quanto à massa fresca e seca, diâmetro dos frutos e concentrações de macronutrientes (N, P, K, Ca e Mg) e micronutrientes (B, Cu, Fe, Mn e Zn). A amostragem de folhas e frutos teve início uma ou duas semanas após a plena floração, dependendo do cultivar, e prolongou-se até a colheita ou até quatro semanas depois, no caso da amostragem de folhas. Nas folhas, de modo geral, as concentrações de N, P, K, Cu e B diminuíram, a concentração de Ca aumentou e as concentrações de Mg, Fe, Mn e Zn pouco variaram ao longo do ciclo vegetativo da cultura. Nos frutos, inicialmente, as concentrações dos nutrientes diminuíram rapidamente e depois decresceram lenta e continuamente, chegando quase constantes à fase final da maturação, indicando diluição dos nutrientes, uma vez que a quantidade total de nutrientes acumulada aumentou gradativamente com o crescimento dos frutos. O potássio foi o nutriente presente em maior quantidade nos frutos de macieiras, tendo sido, portanto, o mais removido do solo.
Malus domestica; macronutrientes; micronutrientes; acumulação
SOILS AND PLANT NUTRITION
Seasonality of nutrients in leaves and fruits of apple trees
Sazonalidade de nutrientes em folhas e frutos de macieira
Gilmar Ribeiro NachtigallI, * * Corresponding author < gilmar@cnpuv.embrapa.br> ; Antonio Roque DechenII
IEmbrapa Uva e Vinho, C.P. 130 - 95700-000 - Bento Gonçalves, RS - Brasil
IIUSP/ESALQ - Depto. de Ciência do Solo, C.P. 9 - 13418-900 - Piracicaba, SP - Brasil
ABSTRACT
The nutrient accumulation curves of apple trees are good indicators of plant nutrient demand for each developmental stage. They are also a useful tool to evaluate orchard nutritional status and to estimate the amount of soil nutrient removal. This research aimed at evaluating the seasonality of nutrients in commercial apple orchards during the agricultural years of 1999, 2000, and 2001. Therefore, apple tree leaves and fruits of three cultivars 'Gala', 'Golden Delicious' and 'Fuji' were weekly collected and evaluated for fresh and dry matter, fruit diameter and macronutrient (N, P, K, Ca and Mg) and micronutrient (B, Cu, Fe, Mn, and Zn) concentrations. Leaf and fruit sampling started one or two weeks after full bloom, depending on the cultivar, and ended at fruit harvest or four weeks later (in the case of leaf sampling). In general, leaf concentrations of N, P, K, Cu, and B decreased; Ca increased; and Mg, Fe, Mn, and Zn did vary significantly along the plant vegetative cycle. In fruits, the initial nutrient concentrations decreased quickly, undergoing slow and continuous decreases and then remaining almost constant until the end of fruit maturation, indicating nutrient dilution, once the total nutrient accumulation increased gradually with fruit growth. Potassium was the nutrient present in highest quantities in apple tree fruits and thus, the most removed from the soil.
Key words:Malus domestica, macronutrients, micronutrients, accumulation
RESUMO
As curvas de acúmulo de nutrientes pela macieira têm se mostrado como um bom parâmetro indicador da demanda da cultura em cada etapa de desenvolvimento, sendo importante para avaliar o estado nutricional do pomar e estimar a remoção de nutrientes do solo. O objetivo do presente trabalho foi avaliar a sazonalidade de nutrientes na macieira, em pomares comerciais, durante as safras agrícolas de 1999, 2000 e 2001. Para tanto, amostras de folhas e frutos de três cultivares 'Gala', 'Golden Delicious' and 'Fuji' foram coletadas semanalmente e avaliadas quanto à massa fresca e seca, diâmetro dos frutos e concentrações de macronutrientes (N, P, K, Ca e Mg) e micronutrientes (B, Cu, Fe, Mn e Zn). A amostragem de folhas e frutos teve início uma ou duas semanas após a plena floração, dependendo do cultivar, e prolongou-se até a colheita ou até quatro semanas depois, no caso da amostragem de folhas. Nas folhas, de modo geral, as concentrações de N, P, K, Cu e B diminuíram, a concentração de Ca aumentou e as concentrações de Mg, Fe, Mn e Zn pouco variaram ao longo do ciclo vegetativo da cultura. Nos frutos, inicialmente, as concentrações dos nutrientes diminuíram rapidamente e depois decresceram lenta e continuamente, chegando quase constantes à fase final da maturação, indicando diluição dos nutrientes, uma vez que a quantidade total de nutrientes acumulada aumentou gradativamente com o crescimento dos frutos. O potássio foi o nutriente presente em maior quantidade nos frutos de macieiras, tendo sido, portanto, o mais removido do solo.
Palavras-chave:Malus domestica, macronutrientes, micronutrientes, acumulação
INTRODUCTION
Foliar analysis is widely used to evaluate the nutritional status of apple tree plants. Lately, fruit chemical analysis has received special attention, since it can provide information on fruit quality based on previously known adequate and critical nutrient levels and, therefore, prevent deficiencies and physiological disturbances in fruits (Marcelle, 1984; Suzuki & Argenta, 1994; Nachtigall & Freire, 1998; Ernani et al., 2002).
The nutrient accumulation curves of apple trees are good indicators of nutrient demand in each plant developmental stage. There are few studies on leaf and fruit nutrient seasonality of apple trees. In Washington (US) N, P, K, Ca, and Mg accumulated quantities were evaluated along the vegetative cycle of six apple tree orchards (Rogers et al., 1953). In North Carolina (US) the seasonal N, P, K, Ca, and Mg accumulation of an apple tree cultivar of 'Delicious' group were evaluated (Chuntanaparb & Cummings, 1980). In Virginia (US) K, Ca, and Mg leaf accumulation of eight cultivars of 'Delicious' group and eight of 'Virgínia Gold' group were evaluated (Hilmelrich & Walker, 1982). In Poland the seasonality of N, P, K, Ca, and Mg accumulation was evaluated in 40 orchards during three successive crops (Nurzynski et al., 1990).
Similarly in Brazil, the nutrient accumulation in apple fruits of 'Ohio Beauty' and 'Brasil', at Buri County, State of São Paulo was evaluated (Trani et al., 1981), and at Palmas County, State of Paraná the nutrient accumulation seasonality in fruits of Gala, Golden Delicious and Fuji cultivars was evaluated (Pavan et al., 1988). However, these studies are frequently out-of-date due to the release of new cultivars and grafts, and also, to new techniques for crop management that can change the apple tree pattern of leaf and fruit nutrient accumulation and levels (Eaton & Robinson, 1977). Furthermore, no information about seasonality of leaf nutrient accumulation in apple trees is available for the States of Rio Grande do Sul and Santa Catarina, in the south region of Brazil, where most of the Brazilian apple production is concentrated.
The objective of this paper was to evaluate the seasonality of nutrients in leaves and fruits of apple trees of Gala, Golden Delicious and Fuji cultivars, during the period between bloom and fruit harvest.
MATERIAL AND METHODS
This research was carried out during the agricultural years of 1998/99, 1999/00 and 2000/01, at Vacaria, RS, Brazil (28°30' S, 50°42' W and 955 m of altitude), in Gala, Golden Delicious and Fuji apple cultivar orchards, grafted on EM VII rootstocks. The orchards were established in 1982 on a Humic Xanthic Hapludox (Streck et al., 2002), whith the following characteristics: clay = 540 g kg-1; organic matter = 58.8 g dm-3, pHH2O (1:1) = 5.8; P = 3.5 mg dm-3; K = 88 mg dm-3; Al = 0.4 mmolc dm-3; and Ca + Mg = 2.7 mmolc dm-3. Crop management practices for weed and pest control (fungicide and insecticide applications), tree formation and orchard maintenance were the usual ones for apples, plus soil and foliar fertilization monitored by means of leaf analysis.
In order to monitor the seasonality of nutrient accumulation, apple tree samples of leaves and fruits were weekly collected, starting one or two weeks after full bloom, depending on the cultivar, until fruit harvest or until four weeks later for leaf sampling, as follows: (1) for the 1st crop (1998/99) sampling started in 29/10/98 and continued for 21, 24 and 29 weeks (for leaves) and for 15, 19 and 21 weeks (for fruits) of Gala, Golden Delicious and Fuji cultivars, respectively; (2) for the 2nd crop (1999/00) sampling started in 11/10/99 and continued for 22, 24 and 30 weeks (for leaves) and for 18, 20 and 26 weeks (for fruits) for cultivars Gala, Golden Delicious and Fuji, respectively; (3) in the 3rd crop sampling started in 19/10/00 and continued for 21, 25 and 25 weeks (for leaves) and for 16, 20 and 25 weeks (for fruits) for cultivars Gala, Golden Delicious and Fuji, respectively.
Leaves were collected from the medium third of branches of the year (one leaf from each plant quarter) of 10 plants to obtain one composite sample (40 leaves). The first fruit sampling was made during the first up to third week after full bloom: 12 fruits plus peduncle were taken per sample. During the fourth until the eighth week after full bloom five fruits plus peduncle were taken per sample and after that until fruit harvest, three fruits plus peduncle were collected per sample. Fruits were taken from four plant quarters, at medium plant height (using as standards, fruits from marked floral bunches at bloom in order to obtain fruits of same age). Fruit samples were measured for diameter and fresh mass. Both leaf and fruit samples were dried in a forced air oven at 60°C until constant mass, for dry mass determination. After that, samples were ground in a Willey type mill and submitted to procedures for macro and micronutrient analyses, as follows: leaf nitrogen (N) was determined by the semi-micro Kjeldahl method, and leaf phosphorus (P), potassium (K), calcium (Ca) and magnesium (Mg) concentrations were determined in extracts obtained through nitric-perchloric digestion. Phosphorus was determined by spectrophotometry, potassium by flame photometry and calcium and magnesium by atomic absorption spectrophotometry, following procedures described by Freire (1998). Leaf micronutrient concentrations copper (Cu), iron (Fe), manganese (Mn) and zinc (Zn) were determined by atomic absorption spectrophotometry in extracts obtained through nitric/sulfuric/perchloric acid digestion. Leaf boron (B) concentration was determined by spectrophotometry using the Azomethine-H method, in extracts obtained from leaf ashes (oven digestion) according to procedure described by Freire (1998).
The data were submitted to regression analysis and the model adopted was chosen using the criterion of determination coefficient magnitude (P < 0.05). The t test (P < 0.05) was used in order to compare mean cultivars. The accumulated nutrient quantities by the crop in fresh fruits were submitted to analysis of variance.
RESULTS AND DISCUSSION
In general, leaf N, P, K, Cu and B concentrations decreased along the apple tree vegetative cycle, calcium increased and little variation was observed in Mg, Fe, Mn and Zn concentrations (Figure 1). These results are similar to those obtained by Hilmelrich & Walker (1982), when working with eight cultivars of the 'Delicious' group and eight of the 'Virginia Gold' group: the authors observed that leaf Ca concentration increased, K decreased and Mg remained constant during the vegetative cycle. In the same way, Rogers et al. (1953) observed that N concentration decreased slightly up to 140 days after bloom passing to a sharp decrease starting from this period; P decreased along the cycle, mainly after 140 days after bloom, K decreased in a gradual and continuous way along the cycle and Ca increased quickly during the first 65 days after bloom remaining practically constant until 155 days after bloom and increasing again from this date on. The reduction observed for N and P concentrations followed a cubic model, where the concentrations were higher during the first five weeks after full bloom, decreased slowly between the fifth and the twentieth week after full bloom and presented a sharp fall from the twentieth week after full bloom. The reduction in leaf K concentrations followed a quadratic model, except for the Gala cultivar, for which leaf K concentrations fit a linear model. Leaf K reduction was practically uniform along the vegetative cycle except for the Golden Delicious cultivar, for which leaf K increased until the eighth week after full bloom and continuously decreased from this period on. Leaf Cu and B concentrations decreased more sharply in the first five weeks after full bloom and from then on, slowly and gradually along the vegetative cycle. Leaf Cu concentration decreases followed a cubic model for the three cultivars, meanwhile leaf B decreases fit to a linear model, significant only for the Gala cultivar.
Young leaf tissues usually present lower water content and higher N, P and K concentrations, meanwhile older tissues are richer in Ca, Mn, Fe and B mainly (Mengel & Kirkby, 2001). Therefore, the nutrient concentration decreases, mainly N, P and K, observed in apple leaves along the vegetative cycle, can be related to a dilution effect occurred with leaf growth and to the nutrient redistribution to other plant organs until the end of the cycle. The increase in leaf Ca concentration along this period can be explained by Ca immobility in plant tissues and no redistribution to other plant organs. Magnesium increases, however, are probably a consequence of lower K competition, since leaf K decreased along the cycle.
Leaf Ca concentrations increased faster during the first five weeks after full bloom, passing to slow and continuous increases from this period on, fitting to cubic models for the three cultivars. Golden Delicious cultivar presented lower leaf Ca concentrations, which practically did not vary along the cycle, what might be explained by the antagonism with K, present in higher concentrations in this cultivar.
Leaf Mg concentrations increased according to cubic models for the Gala and Fuji cultivars, showing sharper increases in the first five weeks after full bloom, passing to slow increases from this period on. For the cultivar Golden Delicious no clear tendency along the cycle was verified.
Leaf Fe, Mn and Zn concentrations fit to cubic models only for the Fuji cv: Fe increased along de cycle; Mn and Zn decreased until the twenty fifth week after full bloom and increased from then on. The variability in leaf Fe, Mn and Zn concentrations observed along the vegetative cycle of the three cultivars might be attributed to leaf contamination with products sprayed in the orchard for pest and disease control. Although this contamination has impaired a clear trend definition of these micronutrient concentrations in apple leaves, it might be considered that this is a frequent and usual occurrence in apple orchards as a function of crop management procedures and has been reported in other works (Pavan et al., 1988). Thus, the leaf micronutrient distribution along the cycle will vary according to the referred practices and products sprayed.
The obtained apple leaf nutrient distribution, in practical terms, allowed to infer that leaf sampling for nutritional diagnosis might be anticipated for 30 days in relation to the period used in the present experiment and reported in the literature (14th to 18th week after full bloom), because there is a relative nutrient concentration stability from the 10th week after full bloom on. Moreover, these results might be useful as standard reference values of different apple tree phenological stages for the leaf analysis interpretation of current laboratory data.
Small but significant differences were observed among cultivars for leaf K, Ca, Mg, Zn and Mn concentrations. Thus, the three cultivars differed for leaf K concentrations (P < 0.01) and the highest leaf K was found for the 'Golden Delicious' cultivar. For Ca, 'Golden Delicious' presented higher leaf Ca than 'Gala' (P < 0.05) and than 'Fuji' (P < 0.01), while for Mg, 'Golden Delicious' differed from the others (P < 0.01) presenting the lowest leaf Mg concentration. For Zn, 'Gala' had higher leaf Zn than the other cultivars (P < 0.01); and for Mn, significant differences among cultivars were also observed (P < 0.01). Although such differences were found among cultivars for some leaf nutrient concentrations, these results are not an indicative of requirement for individual cultivar diagnosis.
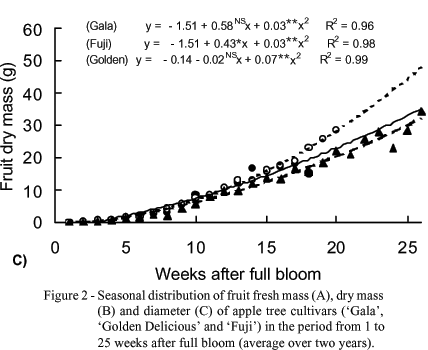
Fruit fresh and dry mass accumulation was small until the 7th week after full bloom representing 14, 13 and 8% of the fresh or final dry mass for 'Gala', 'Golden Delicious' and 'Fuji', respectively (Figure 2, A and B ). From this period on, more expressive increments were obtained, so that, in the 12th week after full bloom the fruit mass (dry or fresh) represented 58, 46 and 32% of the final mass for 'Gala', 'Golden Delicious' and 'Fuji', respectively. These results are in agreement with those obtained by Pavan et al. (1988). Since 'Fuji' is a late cycle cultivar, from the 14th week after full bloom on, lower fresh and dry mass accumulation was observed as compared to the other cultivars. However, at the end of the cycle, the obtained values for fruit fresh and dry mass accumulation were not significantly different among the three cultivars.

Fruit diameters increased continuously up to the 16th week after full bloom and from this period on, smaller increments in fruit diameter were obtained for the three cultivars (Figure 2 C), but no significant differences among cultivars were observed.
Apple fruit nutrient concentrations (N, P, K, Ca, Mg, Fe, Mn, Zn, Cu and B) of 'Gala', 'Golden Delicious' and 'Fuji' were relatively high in the initial fruit development, decreasing systematically with growth (Figure 3). The observed pattern suggested three different phases for fruit nutrient accumulation: a) in the first three weeks after full bloom there was a fast decrease in nutrient concentrations with fruit growth; b) from the 4th to the 10th week after full bloom, slow and continuous decreases in nutrient concentrations were observed; and c) from the 10th week after full bloom until the end of fruit maturation, nutrient concentrations remained almost constant. The first phase occurs during the cellular division period, while the other phases are associated with the period of cellular expansion (Hilmelrick & McDuffie, 1983). This is the so-called effect of chemical dilution, that is, reduction in the nutrient concentrations caused by the increase in fruit dry matter.
No significant differences were observed among cultivars in to relation fruit nutrient concentrations, except for K ('Golden Delicious' presented higher fruit K than 'Fuji', P < 0.05), Mn and Cu ('Gala' had higher values than 'Fuji', P < 0.01 and P < 0.05, respectively). The same tendency was reported by Hilmelrick & Walker (1982) and Pavan et al. (1988). However, Pavan et al. (1988) observed the lowest nutrient concentrations for 'Gala' as compared to 'Golden Delicious' and 'Fuji'.
Fruit Fe concentration variability along the vegetative cycle can be attributed to leaf sample contamination during orchard spraying for pest and disease control.
The total N, P, K, Ca, Mg, Fe, Mn, Zn, Cu and B quantities accumulated in fruits of the tree cultivars increased gradually with fruit growth (Figure 4). This was also obtained by Trani et al. (1981) and Pavan et al. (1988). For the 'Gala' orchard, significant linear regressions were obtained for total N, P, Ca, Mg and B quantities accumulated in fruits, while for K, Cu and Mn, the data significantly fitted to quadratic models. For the 'Golden Delicious', significant positive linear regressions were obtained for total N, P, Ca and Mn in the fruits, while for K, Mg, B, Cu and Fe, the data significantly adjusted to quadratic models. For the 'Fuji', significant positive linear regressions were obtained for total N, K, Mg and Zn, while for P, Ca, B and Cu fitted to quadratic models.
The total nutrient quantities accumulated in dry fruits and removed from orchards during harvest followed a decreasing order: a) 'Gala': K> N> P> Mg = Ca> Fe> Mn> B> Cu> Zn; b) 'Golden Delicious': K> N> Mg = P> Fe> Ca> B> Mn> Cu> Zn; c) 'Fuji': K> N> P> Mg> Ca> Fe> B> Cu> Mn> Zn. Most K accumulation in apple tree fruits was also observed by Trani et al. (1981) and Pavan et al. (1988). The period of higher fruit nutrient accumulation increments, for the three cultivars, occurred from the 8th to 16th week after full bloom, which corresponded to the same period of higher fruit dry mass increments. Similar results were described by Pavan et al. (1988). Except for Ca, there were no significant differences in nutrient accumulation among cultivars. The 'Golden Delicious' total Ca accumulated in the fruits differed significantly (P < 0.05) from the 'Fuji' cultivar.
Nutrients removed by apple tree fresh fruits at harvest of cultivars 'Gala', 'Golden Delicious' and 'Fuji' are presented in Table 1. The data on fruit nutrient quantity removed at harvest in the present experiment were inferior in relation to the ones reported by Pavan et al. (1988), for the same cultivars, but similar to the ones reported by Trani et al. (1981).
Regardless of the cultivar, the apple tree fruits removed higher quantities of potassium as compared to the other nutrients. Although no significant differences were found among cultivars for the total nutrient quantities removed during fruit harvest, it is pointed out that the 'Golden Delicious' total K/Ca ratio was higher in comparison to the other cultivars, because this cultivar removed high amounts of K and relatively low amounts of Ca, which might induce K-Ca unbalance. These results suggested that 'Golden Delicious' is more susceptible to present Ca deficiency in the fruits and requires more careful monitoring of Ca nutrition.
CONCLUSIONS
The apple leaf nutrient concentrations along the vegetative cycle presented relative stability after the 10th week after full bloom, indicating that leaf sampling for nutritional diagnosis might be anticipated for 30 days.
Leaf nutrient concentration can be used as standard reference values of different apple tree phenological stages for leaf analysis interpretation of current laboratory data.
The quantities of accumulated nutrients increased gradually with fruit growth. Potassium was the most accumulated nutrient in fruits, followed by N and P, regardless of the cultivar.
'Golden Delicious' requires more careful monitoring of the Ca nutrition to prevent deficiency in the fruits.
Received June 08, 2005
Accepted July 17, 2006
- CHUNTANAPARB, N.; CUMMINGS, G. Seasonal trends in concentration of nitrogen, phosphorus potassium, calcium, and magnesium in leaf portions of apple, blueberry, grape, and peach. Journal of the America Society for Horticultural Science, v.105, p.933-935, 1980.
- EATON, G.W.; ROBINSON, M.A. Interstock effects upon apple leaf and fruit mineral content. Canadian Journal of Plant Science, v.57, p.227-234, 1977.
- ERNANI, P.R.; AMARANTE, C.V.T.; DIAS, J.; BESSEGATO, A.A. Preharvest calcium sprays improve fruit quality of 'Gala' apples in Southern Brazil. Acta Horticulturae, n.594, p.481-486, 2002.
- FREIRE, C.J.S. Manual de métodos de análise de tecido vegetal, solo e calcário. Pelotas: EMBRAPA, CPACT, 1998. 208p.
- HILMELRICH, D.G.; WALKER, C.E. Seasonal trends of calcium, magnesium, and potassium fractions in apple leaf and fruit tissues. Journal of the America Society for Horticultural Science, v.107, p.1078-1080, 1982.
- HILMELRICH, D.G.; McDUFFIE, R.F. The calcium cycle: Uptake and distribution in apple trees. HortScience, v.18, p.147-151, 1983.
- MARCELLE, R.D. Mineral analysis and storage properties of fruit. In: INTERNATIONAL COLLOQUIUM FOR THE OPTIMIZATION OF PLANT NUTRITION, 6., Montpellier, 1984. Proceedings. Montpellier, 1984. p.365-371.
- MENGEL, K.; KIRKBY, E.A. Principles of plant nutrition. 5.ed. Dordrecht: Kluwer Academic Publishers, 2001. 849p.
- NACHTIGALL, G.R.; FREIRE, C.J.S. Previsão da incidência de "Bitter pit" em maçãs através dos teores de cálcio em folhas e frutos. Revista Brasileira de Fruticultura, v.20, p.158-166, 1998.
- NURZYNSKI, J.; KEPTA, M.; KOMOSA, A.; KOZERA, G. Seasonal changes of N, P, K, Ca and Mg content in apple tree leaves during vegetation period. Acta Horticulturae, n.274, p.365-373, 1990.
- PAVAN, M.A.; MARUR, C.J.; MYAZAWA, M. Composição mineral e acúmulo de matéria seca nos frutos das macieiras 'Gala', 'Fuji' e 'Golden Delicious'. Revista Brasileira de Fruticultura, v.10, p.7-14, 1988.
- ROGERS, B.L.; BATJER, L.P.; THOMPSON, A.H. Seasonal trend of several nutrient elements in Delicious apple leaves expressed on a per cent and unit area basis. Proceedings of the America Society for Horticultural Science, v.61, p.1-5, 1953.
- STRECK, E.V.; KÄMPF, N.; DALMOLIN, R.S.D.; KLAMT, E.; NASCIMENTO, P.C. do; SCHNEIDER, P. Solos do Rio Grande do Sul. Porto Alegre: EMATER/RS; UFRGS, 2002. 107p.
- SUZUKI, A.; ARGENTA, L.C. Teores minerais na polpa de maçã das cultivars. Gala, Golden Delicious e Fuji. Revista Brasileira de Fruticultura, v.16, p.92-104, 1994.
- TRANI, P.E.; HAAG, H.P.; DECHEN. A.R.; SARRUGE, J.R.; CATANI, C.B. Nutrição mineral de macieira. VI. Acúmulos de nutrientes pelos frutos. Anais da ESALQ, v.38, p.723-801, 1981.

Publication Dates
-
Publication in this collection
20 Sept 2006 -
Date of issue
Oct 2006
History
-
Received
08 June 2005 -
Accepted
17 July 2006