Abstracts
Since citrus flowering is a key process in citriculture and its evaluation is often difficult due to the canopy structure and field sampling, the aim of this research was to give some directions regarding the evaluation of flowering in field-grown sweet orange plants. This study was conducted in a citrus orchard of sweet orange plants cv. 'Valencia' [Citrus sinensis (L.) Osbeck] grafted on 'Cleopatra' mandarin (Citrus reshni hort. ex Tanaka) or 'Rangpur' lime (Citrus limonia Osbeck) rootstocks, with North-South orientation. Generative structures [buds, flowers and fruitlets (diameter < 3 cm)] were quantified weekly between August and November 2005, by using a 1 m² frame positioned at the middle third of plant canopy, sampling a volume of about 1 m³. Frames were divided in two parts so that two people could take measurements, and were positioned at Southeast, Southwest, Northeast and Northwest orientations, using seven plants. The following flowering parameters were: (i) number of plants necessary for a representative evaluation of flowering; (ii) plant canopy position to be sampled, and (iii) volume to be evaluated. When considering practical aspects of crop production, a rapid, simple and representative method for flowering evaluation is necessary, especially for growers that frequently have to make strategic decisions about the management of citrus groves. The flowering of field-grown 'Valencia' sweet orange plants can be rapidly assessed by considering at least five plants and sampling canopy volumes of 0.5 m³ in at least one canopy position on each side of the plant row.
Citrus spp.; bloom; method; sampling
O florescimento dos citros é um processo chave na citricultura e sua avaliação é dificultada devido à estrutura da copa e amostragem em campo. O objetivo desse artigo foi fornecer algumas indicações de como avaliar o florescimento de laranjeiras em condição de campo. Esse estudo foi conduzido em um pomar de laranjeiras 'Valência' [Citrus sinensis (L.) Osbeck] enxertadas em tangerineira 'Cleópatra' (Citrus reshni hort. ex Tanaka) ou limoeiro 'Cravo' (Citrus limonia Osbeck), com orientação Norte-Sul. As estruturas reprodutivas [botões florais, flores e frutos (diâmetro < 3 cm)] foram quantificadas semanalmente entre agosto e novembro de 2005, utilizando guias de 1 m² posicionadas no terço médio da copa das plantas, amostrando aproximadamente um volume de 1 m³. As guias foram divididas em duas partes para que duas pessoas pudessem realizar as avaliações, e posicionadas nas orientações sudeste, sudoeste, nordeste e noroeste, em sete plantas. Alguns aspectos do florescimento dos citros foram avaliados: (i) quantas plantas são necessárias para uma amostragem representativa do florescimento; (ii) em qual orientação deve ser feita a medida e (iii) qual volume da copa das plantas que deve ser amostrado. Ao se considerar os aspectos práticos da produção dos citros, um método rápido, simples e representativo é necessário para avaliar o florescimento, especialmente quando produtores têm que freqüentemente tomar decisões estratégicas no manejo dos pomares. O florescimento de laranjeiras 'Valência' em condição de campo pode ser rapidamente avaliado considerando-se pelo menos cinco plantas e amostrando-se volumes de copa de 0,5 m³ em pelo menos uma posição da copa em cada lado da linha de plantio.
Citrus spp.; brotação; método; amostragem
PLANT PATHOLOGY AND BIOCHEMISTRY
'Valencia' sweet orange tree flowering evaluation under field conditions
Avaliação do florescimento de laranjeiras valência em condição de campo
Rafael Vasconcelos RibeiroI, * * Corresponding author < rafael@iac.sp.gov.br> ; Glauco de Souza RolimI; Fernando Alves de AzevedoII; Eduardo Caruso MachadoI
IAPTA/IAC - Centro de Pesquisa e Desenvolvimento em Ecofisiologia e Biofísica, C.P. 28 - 13012-970 - Campinas, SP - Brasil
IIAPTA/IAC - Centro Avançado de Pesquisa Tecnológica do Agronegócio de Citros 'Sylvio Moreira', C.P. 04 - 13490-970 - Cordeirópolis, SP - Brasil
ABSTRACT
Since citrus flowering is a key process in citriculture and its evaluation is often difficult due to the canopy structure and field sampling, the aim of this research was to give some directions regarding the evaluation of flowering in field-grown sweet orange plants. This study was conducted in a citrus orchard of sweet orange plants cv. 'Valencia' [Citrus sinensis (L.) Osbeck] grafted on 'Cleopatra' mandarin (Citrus reshni hort. ex Tanaka) or 'Rangpur' lime (Citrus limonia Osbeck) rootstocks, with North-South orientation. Generative structures [buds, flowers and fruitlets (diameter < 3 cm)] were quantified weekly between August and November 2005, by using a 1 m2 frame positioned at the middle third of plant canopy, sampling a volume of about 1 m3. Frames were divided in two parts so that two people could take measurements, and were positioned at Southeast, Southwest, Northeast and Northwest orientations, using seven plants. The following flowering parameters were: (i) number of plants necessary for a representative evaluation of flowering; (ii) plant canopy position to be sampled, and (iii) volume to be evaluated. When considering practical aspects of crop production, a rapid, simple and representative method for flowering evaluation is necessary, especially for growers that frequently have to make strategic decisions about the management of citrus groves. The flowering of field-grown 'Valencia' sweet orange plants can be rapidly assessed by considering at least five plants and sampling canopy volumes of 0.5 m3 in at least one canopy position on each side of the plant row.
Key words:Citrus spp., bloom, method, sampling
RESUMO
O florescimento dos citros é um processo chave na citricultura e sua avaliação é dificultada devido à estrutura da copa e amostragem em campo. O objetivo desse artigo foi fornecer algumas indicações de como avaliar o florescimento de laranjeiras em condição de campo. Esse estudo foi conduzido em um pomar de laranjeiras 'Valência' [Citrus sinensis (L.) Osbeck] enxertadas em tangerineira 'Cleópatra' (Citrus reshni hort. ex Tanaka) ou limoeiro 'Cravo' (Citrus limonia Osbeck), com orientação Norte-Sul. As estruturas reprodutivas [botões florais, flores e frutos (diâmetro < 3 cm)] foram quantificadas semanalmente entre agosto e novembro de 2005, utilizando guias de 1 m2 posicionadas no terço médio da copa das plantas, amostrando aproximadamente um volume de 1 m3. As guias foram divididas em duas partes para que duas pessoas pudessem realizar as avaliações, e posicionadas nas orientações sudeste, sudoeste, nordeste e noroeste, em sete plantas. Alguns aspectos do florescimento dos citros foram avaliados: (i) quantas plantas são necessárias para uma amostragem representativa do florescimento; (ii) em qual orientação deve ser feita a medida e (iii) qual volume da copa das plantas que deve ser amostrado. Ao se considerar os aspectos práticos da produção dos citros, um método rápido, simples e representativo é necessário para avaliar o florescimento, especialmente quando produtores têm que freqüentemente tomar decisões estratégicas no manejo dos pomares. O florescimento de laranjeiras 'Valência' em condição de campo pode ser rapidamente avaliado considerando-se pelo menos cinco plantas e amostrando-se volumes de copa de 0,5 m3 em pelo menos uma posição da copa em cada lado da linha de plantio.
Palavras-chave: Citrus spp., brotação, método, amostragem
INTRODUCTION
Fruit production is one of the main economical aspects in citriculture, which is an important agricultural activity in many countries around the world (FAO, 2006). Adequate environmental conditions and suitable plant nutrition are two essential needs for citrus production; nevertheless, a key developmental process that determines the orchard yield is flowering. Since flowering and fruit set are important elements involved in the forecast of citrus production, many studies have been carried out to evaluate these phenological phases (Araújo et al., 1999; Agustí et al., 2000; Koller et al., 2000; Tonietto & Tonietto, 2005).
The evaluation of citrus flowering is complex due to the canopy structure, uneven flowering, and sampling issues in field-grown plants. An important point is the flowering evaluation method, which is not standardized and thus forces comparisons to previous literature. Regarding flowering evaluation, some authors have considered only branches or shoots of the plant canopy (Lomas & Burd, 1983; Rocha et al., 1990; Nuñez et al., 1992; Koshita et al., 1999; Tonietto & Tonietto, 2005), while others have used a visual scale or taken measurements with frames (hollow squares with varying areas) positioned at single or multiple regions around the plant canopy (Holguín et al., 1992; Nuñez et al., 1992; Araújo et al., 1999; Koller et al., 2000; Sanches et al., 2001).
The aim of this research is to give some directions in relation to the evaluation of flowering in field-grown sweet orange plants, answering some questions such as: how many plants should be evaluated? Which canopy positions should be considered for sampling? Which volume of the canopy should be evaluated? The above aspects were addressed to optimize the method for studying citrus flowering, maximizing the field work and obtaining reliable data on flowering intensity. Since flowering may be influenced by rootstock (Holguín et al., 1992; Tonietto & Tonietto, 2005), this work was developed with 'Valencia' sweet orange plants grafted on 'Cleopatra' mandarin and 'Rangpur' lime rootstocks.
MATERIAL AND METHODS
Experimental site
This study was conducted in a 15-year-old citrus orchard grove of sweet orange plants cv. 'Valencia' [Citrus sinensis (L.) Osbeck] grafted on 'Cleopatra' mandarin (Citrus reshni hort. ex Tanaka) and 'Rangpur' lime (Citrus limonia Osbeck) rootstocks, spaced 8 × 5 m, with North-South row orientation. The orchard is located in Cordeirópolis, São Paulo State, Brazil (22°32' S; 47°27' W; altitude 639 m). The climate is subtropical, with an average annual rainfall of 1366 mm and mean monthly air temperature varying between 23.8ºC and 17.8ºC. Plants were grown under natural environmental conditions, i.e. non-irrigated and exposed to natural variations of solar radiation, air temperature and humidity.
Flowering evaluation
Generative structures were classified as buds (only sepals visible, corresponding to types 56, 57 and 59 [Agustí et al., 2000]), flowers (whenever exists a single flower structure, such as petal, sepal, stamen or pistil, corresponding to types 60, 61, 65, 67 and 69 [Agustí et al., 2000]) and fruitlets (without flower structure and with fruit up to 3.0 cm of diameter, corresponding to types 71, 72 and 73 [Agustí et al., 2000]). These structures were quantified weekly between August and November 2005, using square frames of 1 m2 (1 × 1 m) positioned at the middle third of tree canopy (sampling the volume portion between 1.5 and 2.5 m height). Mid-canopy was sampled since a large variation in some fruit quality attributes has been reported between top and bottom canopy positions (Syvertsen & Albrigo, 1980; Davies & Zalman, 2004).
Frames were positioned at Southeast (SE), Southwest (SW), Northeast (NE) and Northwest (NW) orientations. Frames were divided in two areas of 0.5 m2 in order to allow two people to take measurements. Considering the quantification of generative structures down to 1 m inside tree canopy, the sampled volumes were 0.5 or 1 m3. Seven plants were used for evaluating flowering per planting row. The central tree (located at the middle of the plant row) was fixed and then three trees at each side were evaluated. Data refers to evaluations made before, during and after full flowering, between Sept. 16 and Nov. 11 for Valencia/Cleopatra and between Sept. 16 and Nov. 24 for Valencia/Rangpur. The flowering peaks were defined as the dates on which the highest sum of generative structures (buds + flowers + fruitlets) occurred. An additional evaluation was carried out on March 31, 2006 to determine the amount of fruit in each canopy position. This data was compared to the amount of generative structures observed at full flowering and then fruit set (%) was calculated.
Data analysis
The analysis to determinate how many plants are necessary for a representative evaluation of 'Valencia' sweet orange flowering was made by an iterative process that compared the mean value and its standard error (SE) of one to seven plant samplings, considering all generative structures quantified in 1 m3. The minimum number of plants to be evaluated was defined as the one with mean value close to the mean value of seven samplings and with small variation of SE (< ± 15%) in relation to the data variability found in the sampling of seven plants. Considering seven trees in one planting row, the tree located at the central part was fixed in all samplings (n = 1 to 7), giving one combination for groups of one (central tree) and seven trees (all trees) (n = 1 and n = 7), six combinations for groups of two and six trees (n = 2 and n = 6), 15 combinations for groups of three and five trees (n = 3 and n = 5), and 20 combinations for groups of four trees (n = 4).
The flowering in relation to plant canopy position was evaluated by comparing the mean value (n = 7) of all generative structures quantified at each position (SE, SW, NE and NW) and by evaluating fruit set in these positions. Another analysis made in this work was the sampling of generative structures taking into account 0.5 (half guide) and 1.0 m3 (entire guide) of sampled canopy volume. Data were submitted to ANOVA procedure and mean values were compared by the Tukey test (p < 0.05).
RESULTS AND DISCUSSION
Sampling size
In the first evaluations, approximately 15 and 30 days before the flowering peak (Figure 1A,B), the mean value of total generative structures was almost constant regardless of the sample size for Valencia/Cleopatra. For the other evaluation dates, the sampling of a single plant led to underestimation (Figure 1C,D) or to overestimation (Figure 1E) of the mean value as compared to the sampling of seven plants. For Valencia/Rangpur the sampling of one plant was always unreliable in relation to the sampling of seven plants (Figure 1F,G), giving a higher overestimation in the first evaluation date, i.e. 30 days before full flowering. Before the flowering peak, there were decreasing (Figure 1F) or increasing (Figure 1G) trends of generative structures in relation to sample size increase, depending on the evaluation date. These trends were mostly due to the chance of the first two selected trees having higher or lower amounts of generative structures than the average.
In general, the sampling size considering five plants gave mean values of generative structures (buds + flowers + fruitlets) similar to samplings of six and seven plants, in spite of scion/rootstock combination. However, one important and essential aspect of flowering characterization is to define the number of plants to be evaluated to obtain a representative analysis of data variability, considered as the standard error of the mean (SE). In fact, SE values tended to stabilize and remain constant when considering samples of five to seven plants (Figure 1).
According to the number of reproductive structures per m3 of plant canopy (Figure 1), the flowering evaluation in field-grown 'Valencia' sweet orange plants should consider a sample of at least five plants in order to obtain reliable and representative data, based on similar mean values and data variability (SE) of the five and seven plant samplings. This sample size minimizes the time spent with this evaluation and improve the orchard management when one should make decisions that are dependent on the flowering stage.
One interesting point is that the number of total generative structures (buds + flowers + fruitlets) per m3 of plant canopy at full flowering was around 500 in Valencia/Cleopatra and 560 in Valencia/Rangpur (Figure 1C,H), suggesting an influence of the rootstock on flowering. In fact, the number of generative structures is related to citrus production (Guardiola et al., 1984) and it is known that the 'Rangpur' lime rootstock induces more crop loading in sweet orange shoots and tolerance to water deficit, being the most used rootstock in Brazil (Davies & Albrigo, 1994). Scion/rootstock combination also affects the occurrence of vegetative/generative flushes and the duration and period of reduced plant growth, commonly observed during winter (June to August in the South hemisphere) (Volpe, 1992).
Sampling position
An important aspect in citrus flowering evaluation is which canopy position in relation to cardinal orientation should be considered for sampling. However, little importance has been given to this detail, which is a probable reason for some contradictory results in relation to citrus flowering (Krajewski & Rabe, 1995). No differences were detected between canopy positions in Valencia/Cleopatra (Figure 2A,B,D,E), except for evaluations carried out near to flowering peak (Figure 2C). However, some authors have found significant differences between canopy positions when considering flowering (Rocha et al., 1990) or fruit quality attributes (Syvertsen & Albrigo, 1980; Davies & Zalman, 2004). Such discrepancy is probably due to differential exposure of tree canopy to solar radiation and/or tree architecture, which varies depending on the orchard management practices. On October 14 (Figure 2C,H), the Northeast (NE) and Southeast (SE) positions exhibited more than 500 generative structures, being superior (p < 0.05) to those observed in the Northwest (NW) and Southwest (SW) positions, with less than 450 structures. Accordingly, Araújo et al. (1999) also found the highest number of flowers in the NE and SE positions for 'Pera' sweet orange plants.
The spatial variability of the canopy for 'Valencia' sweet orange grafted on 'Rangpur' lime was higher than on 'Cleopatra' mandarin (Figure 2). Although differences between canopy positions have been observed in some evaluations (Figure 2G,H,J), a clear pattern was not observed. Around 15 days prior to the flowering peak (Figure 2G), the NW and SE canopy positions presented higher numbers of generative structures (p < 0.05), whereas the NW and SE positions exhibited respectively the highest and lowest amount of generative structures (p < 0.05) around 30 days after the flowering peak (Figure 2J).
As observed for Valencia/Cleopatra, the NE and SE positions of Valencia/Rangpur had more generative structures than the others at the flowering peak (Figure 3H). In fact, environmental conditions regulate flower distribution on the tree (Rocha et al., 1990; Davies & Albrigo, 1994; Araújo et al., 1999). Due to the row orientation of the field under study, the NE and SE canopy positions are located at the side of plant row that receives direct solar radiation during the morning, when there is low air temperature and high relative humidity (RH). During afternoon, the NW and SW canopy positions receive higher solar radiation than the other positions, which is accompanied by higher air temperature and lower RH (Ribeiro et al., 2005). These environmental conditions are probably determining these differences between canopy positions in relation to flowering intensity. Higher temperature and low RH induce decreases in leaf water potential, stomatal conductance and leaf CO2 assimilation of exposed leaves (Syvertsen & Albrigo, 1980; Ribeiro, 2006). They may reduce carbohydrate synthesis and the supply to reproductive sinks, being a cause of intense drop of flowers and fruitlets (Garcia-Luis et al., 1988; Goldschmidt, 1999; Iglesias et al., 2003).
When comparing the relation between canopy positions and full flowering, Tonietto & Tonietto (2005) observed that the South position had less open flowers than the others. This spatial pattern was related to distribution and incidence of solar radiation in the canopy, which was lower in the South position. Here the South positions of Valencia/Rangpur showed the highest (SE) and lowest (SW) amount of generative structures at the flowering peak, being in discordance with Tonietto & Tonietto (2005). The lack of relation at full flowering between the present study and Tonietto & Tonietto (2005), may be partially explained by varietal differences, i.e. 'Valencia' vs. 'Tobias' and by the time in which flowering was evaluated. In addition, different years and sites may also affect citrus flowering.
After the flowering peak, canopy positions did not exhibit differences for 'Cleopatra' mandarin considering the amount of generative structures (Figure 2D,E). However, 'Rangpur' lime induced lower (p < 0.05) amount of generative structures in both NE and SE positions around 30 days after the flowering peak (Figure 2J). Since the highest amounts of generative structures were verified in the NE and SE positions at the flowering peak (Figure 2C,H), it is suggested that the drop of generative structures was more intense in the NE and SE for 'Valencia' sweet orange grafted on both rootstocks. Rocha et al. (1990) working with 'Valencia' and other citrus varieties grafted on 'Rangpur' lime also observed that the East position exhibited the lowest fruit set compared to other canopy positions.
Air temperature is one environmental element that regulates flowering intensity and duration (Davies, 1997), causing drop of floral structures or impairment of flower bud development for temperatures higher than 30ºC (Reuther, 1973; Davies & Albrigo, 1994). In fact, some differences have been observed when comparing leaf temperatures in the East and West canopy positions. In an orchard oriented in the South-North direction, maximum temperatures can reach around 40ºC at the West position while at the East they are around 36ºC (Ribeiro et al., 2005). An association between high temperature and drop of generative structures was not evident in this study. However, this issue should be addressed in further studies, considering differential hydration of the plant canopy.
Since the differences observed between canopy positions were not consistent throughout the flowering period for both scion/rootstock combinations, one should consider at least one canopy position on each side of the planting row to get reliable data that does not over or under-estimate citrus flowering. This recommendation may vary according to the row orientation, which is North-South in the evaluated orchard.
Sampling volume
The relation between generative structures (buds, flowers and fruitlets) should be constant for a reliable flowering evaluation, regardless of canopy volume sampled. Three evaluation dates were chosen to assess possible differences in the relative number of generative structures induced by canopy sampling volume (Figure 3). The proportion (%) between generative structures sampled in 0.5 and 1.0 m3 was compared along evaluation dates with higher number of buds (Figure 3A,D), flowers (Figure 3B,E) or fruitlets (Figure 3C,F).
The number of each generative structures was approximately half when comparing 0.5 m3 and 1.0 m3 samplings (Figure 3). In all evaluation dates the proportion between buds, flowers and fruitlets was maintained, regardless scion/rootstock combination. This suggests that the evaluation of 0.5 m3 is reliable for assessing citrus flowering in field-grown plants. Such a recommendation will save time, however, the researcher/worker should have in mind that irregularities in plant canopy must be considered with caution.
Flowering and fruit set
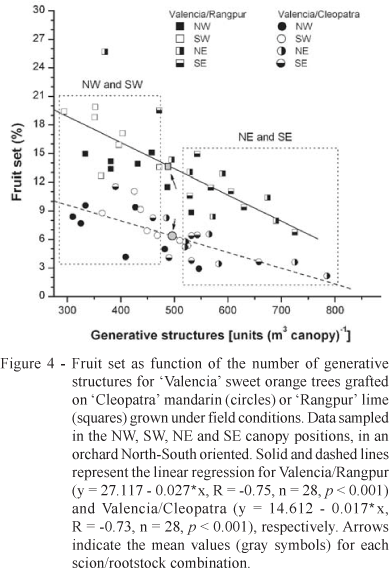
Flowering has a direct relationship with fruit set and crop production. Therefore, an early evaluation of the reproductive status of citrus trees may reveal changes in final crop production, which is economically interesting for growers and industry. High fruit set occurs where there is low amount of flowers, while low fruit set is found where there is high amount of flowers as a consequence of the source-sink relationship (Davies & Albrigo, 1994; Goldschmidt, 1999). This trend was observed for both scion/rootstock combinations, but with Valencia/Cleopatra showing lower fruit set than Valencia/Rangpur (Figure 4).
Hólguin et al. (1992), Davies & Albrigo (1994), Tonietto & Tonietto (2005) reported the influence of rootstock on scion flowering and yield. However, the mean fruit set for both Valencia/Cleopatra (6.4 ± 0.5 %) and Valencia/Rangpur (13.8 ± 0.8 %) combinations was higher than the values observed by Rocha et al. (1990), around 4.4%. At first glance, these differences in fruit set may be induced by climate, crop loading and/or previous crop yield (Reuther, 1973; Goldschmidt, 1999; Albrigo & Saúco, 2004). In general, those fruit set values observed herein (Figure 4) are in the range of the values reported for citrus trees (Davies & Albrigo, 1994; Spiegel-Roy & Goldschmidt, 1996).
Curiously, the NW and SW canopy positions showed higher fruit set and lower amount of generative structures than the NE and SE positions (Figure 4) around 5.5 months after the flowering peak. High values of fruit set were observed, between 2.9 and 11.5% for Valencia/Cleopatra and 6.8 and 25.7% for Valencia/Rangpur, depending on the canopy position (Figure 4). The high fruit set in the West canopy positions was probably caused by the low amount of generative structures rather than to microclimate, which presents small differences between East and West positions (Ribeiro et al., 2005). An essential aspect is that one may overestimate fruit set and underestimate the amount of generative structures when only NW and/or SW canopy positions are sampled. The contrary is also true, an underestimate of fruit set and over-estimate of generative structures may happen when only NE and/or SE positions are considered.
Taking into account practical aspects of crop production, such as spray of chemical substances to control diseases and insects and/or nutrients to maintain healthy plants, a rapid, simple and representative method for evaluating flowering is necessary, especially for growers that often have to make strategic decisions about the management of citrus orchards. As a conclusion it can be said that the flowering of field-grown 'Valencia' sweet orange trees can rapidly be assessed by considering at least five plants using sampling canopy volumes of 0.5 m3 in at least one position on each plant row side.
Received October 11, 2006
Accepted November 29, 2007
- AGUSTÍ, M.; ZARAGOZA, S.; BLEIHOLDER, H.; BUHR, L.; HACK, H.; KLOSE, R.; STAUB, R. The BBCH scale for describing phenological stages in citrus development. In: INTERNATIONAL CONGRESS OF CITRICULTURE, 9., Orlando, 2000. Proceedings Orlando: International Society of Citriculture, 2000. p.445-446.
- ALBRIGO, L.G.; SAÚCO, V.G. Flower bud induction, flowering and fruit-set of some tropical and subtropical fruit tree crops with special reference to citrus. Acta Horticulturae, n.632, p.81-90, 2004.
- ARAÚJO, P.S.R.; MOURÃO FILHO, F.A.A.; SPOSITO, M.B. Pegamento de frutos de laranjeira 'Pera' em diferentes alturas na copa relacionado aos quadrantes geográficos. Scientia Agricola, v.56, p.157-162, 1999.
- DAVIES, F.S. An overview of climatic effects on citrus flowering and fruit quality in various parts of the world. In: CITRUS FLOWERING & FRUITING SHORT COURSE, 1., Lake Alfred, 1997. Proceedings Lake Alfred: CREC/IFAS, 1997.
- DAVIES, F.S.; ALBRIGO, L.G. Citrus Wallingford: CABI, 1994. 254p.
- DAVIES, F.S.; ZALMAN, G.R. Fruit quality sampling of 'Valencia' orange trees. Proceedings of the Florida State Horticultural Society, v.117, p.86-88, 2004.
- FOOD AND AGRICULTURE ORGANIZATION - FAO. FAOSTAT databases Available at: http://faoestat.fao.org Accessed 24 Feb. 2006.
- GARCIA-LUIS, A.; FORNES, F.; SANZ, A.; GUARDIOLA, J.L. The regulation of flowering and fruit set in Citrus: relationship with carbohydrate levels. Israel Journal of Botany, v.37, p.189-201, 1988.
- GUARDIOLA, J.L.; GARCIA-MARÍ, F.; AGUSTÍ, M. Competition and fruit set in Washington navel orange. Physiologia Plantarum, v.62, p.297-302, 1984.
- GOLDSCHMIDT, E.E. Carbohydrate supply as a critical factor for citrus fruit development and productivity. HortScience, v.34, p.1020-1024, 1999.
- HOLGUÍN, E.C.L.; ANGEL, S.G.A.; BAENA, G.D.; ESCOBAR, T.W. Efecto de cinco porta-enjertos sobre la floracion, fructificacion y maduracion de la naranja "Frost Valencia" Citrus sinensis (L.) Osb. en condiciones del CNI-Palmira, Valle del Cauca. Acta Agronomica, v.42, p.81-97, 1992.
- KOLLER, O.L.; SOPRANO, E.; COSTA, A.C.Z.; KOLLER, O.C.; YAMANISHI, O.K. Indução de floração e produção de frutos em laranjeira 'Shamouti'. Laranja, v.21, p.307-325, 2000.
- IGLESIAS, D.J.; TADEO, F.R.; PRIMO-MILLO, E.; TALON, M. Fruit set dependence on carbohydrate availability in citrus trees. Tree Physiology, v.23, p.199-204, 2003.
- KOSHITA, Y.; TAKAHARA, T.; OGATA, T.; GOTO, A. Involvement of endogenous plant hormones (IAA, ABA, GAs) in leaves and flower bud formation of satsuma mandarin (Citrus unshiu Marc.). Scientia Horticulturae, v.79, p.185-194, 1999.
- KRAJEWSKI, A.J.; RABE, E. Citrus flowering: a critical evaluation. Journal of Horticultural Science, v.70, p.357-374, 1995.
- LOMAS, J.; BURD, P. Prediction of the commencement and duration of the flowering period of citrus. Agricultural Meteorology, v.28, p.387-396, 1983.
- NUÑEZ, R.L.; VIDARTE, G.E.Y.; BAENA, G.D.; ESCOBAR, T.W. Estudio de la floracion, fructificacion y produccion de dos variedades de naranja, Citrus sinensis (L.) Osbeck, y dos de manadarina, Citrus reticulata Blanco, en condiciones climaticas de Palmira, Valle del Cauca. Acta Agronomica, v.42, p.50-63, 1992.
- REUTHER, W. Climate and citrus behavior. In: REUTHER, W. The citrus industry. Riverside: University of California, 1973. v.3., chap.9, p.280-337.
- RIBEIRO, R.V. Variação sazonal da fotossíntese e relações hídricas de laranjeira 'Valencia'. Piracicaba: USP/ESALQ, 2006. 157p. Tese (Doutorado).
- RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G. Leaf temperature in sweet orange plants under field conditions: influence of meteorological elements. Revista Brasileira de Agrometeorologia, v.13, p.393-403, 2005.
- ROCHA, A.C.; TAVARES, E.D.; SANDRINI, M.; PAIVA, R.; CARVALHO, S.A. Época e intensidade de florescimento e pegamento de frutos segundo a distribuição pelos quadrantes em laranjeiras. Pesquisa Agropecuária Brasileira, v.25, p.85-88, 1990.
- SANCHES, F.R.; LEITE, I.C.; CASTRO, P.R.C. Efeito do ácido giberélico (AG3) na floração e produção da lima ácida 'Tahiti' (Citrus latifolia Tan.). Revista Brasileira de Fruticultura, v.23, p.504-509, 2001.
- SPIEGEL-ROY, P.; GOLDSCHMIDT, E.E. Biology of citrus Cambridge: Cambridge University Press. 1996. 230p.
- SYVERTSEN, J.P.; ALBRIGO, L.G. Some effects of grapefruit tree canopy position on microclimate, water relations, fruit yield, and juice quality. Journal of the American Society for Horticultural Science, v.105, p.454-459, 1980.
- TONIETTO, S.M.; TONIETTO, A. Floração da variedade Tobias (Citrus sinensis Osbeck.) sobre três porta-enxertos no Vale do Taquari-RS. Revista Brasileira de Fruticultura, v.27, p.14-16, 2005.
- VOLPE, C.A. Fenologia de citros. In: SIMPÓSIO INTERNACIONAL DE CITROS FISIOLOGIA, 2., Bebedouro, 1992. Anais Bebedouro: Fundação Cargill, 1992. p.107-120.
Publication Dates
-
Publication in this collection
21 July 2008 -
Date of issue
2008
History
-
Accepted
29 Nov 2007 -
Received
11 Oct 2006