Abstracts
Incomplete panicle exsertion is one of the symptoms of cold injury at the reproductive stage of the rice plant (Oryza sativa L.), which damages grain yield and raises disease incidence. For this reason, panicle exsertion is a better indicator of cold tolerance under field conditions than spikelet sterility, which may also be affected by other climatic factors. This work studies the variability of degree of panicle exsertion in rice, under the Southern Brazilian environmental conditions and determines the inheritance and heritability of this trait. Four rice genotypes of different cool temperature reactions at the reproductive stage were crossed and field evaluated, with the F2 generation, in relation to the degree of panicle exsertion and spikelet fertility. There was variability among the genotypes for panicle exsertion. The F2 generation of the crosses presented continuous distribution and transgressive segregation towards incomplete exsertion, indicating that genes controlling this trait may be complementary distributed between the parents. Heritability was moderate, so selection for complete panicle exsertion should be applied in advanced generations.
Oryza sativa L.; chilling temperature; cold tolerance; complete panicle exsertion; heritability
A exerção incompleta da panícula é um dos sintomas de dano por frio no estádio reprodutivo da planta de arroz (Oryza sativa L.), o qual prejudica o rendimento de grãos e aumenta a incidência de doenças. Devido a isso, a exerção da panícula é um bom indicador da tolerância ao frio sob condições de campo ao invés da esterilidade de espiguetas, que também pode ser afetada por outros fatores climáticos. Este trabalho objetivou estudar a variabilidade do grau de exerção da panícula de arroz sob as condições ambientais do sul do Brasil e determinar a herança e herdabilidade desta característica. Quatro genótipos de arroz com diferentes reações à temperatura baixa no período reprodutivo foram cruzados e avaliados a campo, juntamente com a geração F2, quanto ao grau de exerção da panícula e fertilidade de espiguetas. Houve variabilidade entre os genótipos para exerção da panícula. A geração F2 apresentou distribuição contínua e segregação transgressiva na direção da exerção incompleta, indicando que os genes para exerção da panícula devem estar distribuídos complementarmente entre os genitores. A herdabilidade foi moderada, portanto seleção para exerção completa da panícula é recomendada em gerações avançadas.
Oryza sativa L.; temperatura baixa; tolerância ao frio; exerção completa da panícula; herdabilidade
GENETICS AND PLANT BREEDING
Inheritance of pinacle exsertion in rice
Herança da exerção da panícula em arroz
Renata Pereira da CruzI,* * Corresponding Author < rpdacruz@hotmail.com> ; Sandra Cristina Kothe MilachII; Luiz Carlos FederizziIII
IIRGA - Divisão de Pesquisa, Equipe de Melhoramento Genético, C.P. 29 - 94930-030 - Cachoeirinha, RS - Brasil
IIPioneer Sementes - Paissandú, 582 - 99010-080 - Passo Fundo, RS - Brasil
IIIUFRGS/FA - Depto. de Plantas de Lavoura, C.P. 776 - 91501-970 - Porto Alegre, RS - Brasil
ABSTRACT
Incomplete panicle exsertion is one of the symptoms of cold injury at the reproductive stage of the rice plant (Oryza sativa L.), which damages grain yield and raises disease incidence. For this reason, panicle exsertion is a better indicator of cold tolerance under field conditions than spikelet sterility, which may also be affected by other climatic factors. This work studies the variability of degree of panicle exsertion in rice, under the Southern Brazilian environmental conditions and determines the inheritance and heritability of this trait. Four rice genotypes of different cool temperature reactions at the reproductive stage were crossed and field evaluated, with the F2 generation, in relation to the degree of panicle exsertion and spikelet fertility. There was variability among the genotypes for panicle exsertion. The F2 generation of the crosses presented continuous distribution and transgressive segregation towards incomplete exsertion, indicating that genes controlling this trait may be complementary distributed between the parents. Heritability was moderate, so selection for complete panicle exsertion should be applied in advanced generations.
Key words:Oryza sativa L., chilling temperature, cold tolerance, complete panicle exsertion, heritability
RESUMO
A exerção incompleta da panícula é um dos sintomas de dano por frio no estádio reprodutivo da planta de arroz (Oryza sativa L.), o qual prejudica o rendimento de grãos e aumenta a incidência de doenças. Devido a isso, a exerção da panícula é um bom indicador da tolerância ao frio sob condições de campo ao invés da esterilidade de espiguetas, que também pode ser afetada por outros fatores climáticos. Este trabalho objetivou estudar a variabilidade do grau de exerção da panícula de arroz sob as condições ambientais do sul do Brasil e determinar a herança e herdabilidade desta característica. Quatro genótipos de arroz com diferentes reações à temperatura baixa no período reprodutivo foram cruzados e avaliados a campo, juntamente com a geração F2, quanto ao grau de exerção da panícula e fertilidade de espiguetas. Houve variabilidade entre os genótipos para exerção da panícula. A geração F2 apresentou distribuição contínua e segregação transgressiva na direção da exerção incompleta, indicando que os genes para exerção da panícula devem estar distribuídos complementarmente entre os genitores. A herdabilidade foi moderada, portanto seleção para exerção completa da panícula é recomendada em gerações avançadas.
Palavras-chave:Oryza sativa L., temperatura baixa, tolerância ao frio, exerção completa da panícula, herdabilidade
INTRODUCTION
Rice is very sensitive to cool temperatures during the reproductive period (Yoshida, 1981). Although the main symptom of damage from cold is the high spikelet sterility (Jacobs & Pearson, 1994), incomplete panicle exsertion has been cited as a symptom of cold injury in many countries (Chung, 1979; Hamdani, 1979; Alvarado & Grau, 1991).
Cold tolerance at the reproductive period has been associated with the degree of panicle exsertion, which could be used as a selection criterion (Nanda & Seshu, 1979). In fact, this trait was suggested as an indicative of genotype adaptability to cool temperatures (Takahashi, 1984). The inheritance of the degree of panicle exsertion under cool temperatures in field conditions was pointed out as having a simple mendelian pattern, with one dominant gene responsible for the complete panicle exsertion (Mahishi et al., 1991; Pandey & Gupta, 1993). More recently, 44 QTLs located on 11 of the 12 rice chromosomes were associated with increased panicle exsertion and described as major genes with overdominance (Han et al., 2006). In a recombinant inbred line population derived from an Indica × Japonica cross, however, a lower number of QTLs clustered in a few chromosomal blocks Inheritance of panicle exsertion in rice explained a small portion of the variation on this trait (Oh et al., 2004), discarding the involvement of major genes in panicle exsertion. In a diallel study, the general combining ability effects were more important than the specific combining ability effects for panicle exsertion (Han et al., 2004), showing that inheritance studies are highly dependent on the genetic material.
In Southern Brazil, in spite of reports on the occurrence of incomplete panicle exsertion due to cool temperatures during the reproductive period (Souza, 1990), there are no data concerning the genetics of this trait under field conditions. So, this work aimed at studying the variability for the degree of panicle exsertion in four rice genotypes and determining the inheritance and heritability of the trait.
MATERIAL AND METHODS
The rice genotypes studied were Quilla 64117, Diamante, IRGA 417 and BRS Taim. The former two are from Chile, belong to the Japonica subspecies and are cold tolerant, with complete panicle exsertion under cool temperatures. The latter two are Brazilian rice cultivars belonging to the Indica subspecies and are cold sensitive, with incomplete panicle exsertion under cool temperatures.
In the 1997/98 growing season, the four genotypes were crossed in a partial diallel and the F1 generation was planted in the next growing season for F2 seed production. In 1999/2000, a field experiment with the four parents and the F2 populations was conducted at Santa Vitória do Palmar, State of Rio Grande do Sul, Brazil, (33º31' S and 53º22' W). This location was chosen because it has the highest incidence of cool temperatures during the rice growing season in Southern Brazil.
Sowing time was purposely chosen to be late, on the 2nd of December, 1999, allowing the coincidence between the rice flowering period and the lowest minimum temperatures, which occur at the end of Feb., begginning of March. Experimental units were 9 m2 plots with six rows 5 m long and 0.3 m between rows, which were machine sowed using 10 g of seed per plot. Plant emergence occurred 11 days after sowing and N fertilization was applied 35, 50 and 60 days after emergence using 22.5 kg ha-1 of N in each application.
The experiment was conducted using a randomized block design with three replications for each treatment (four genotypes and six F2 segregating populations). Twenty plants were randomly labeled in parental each plot and 70 plants per F2 plot. For each labeled plant, the main tiller was individually identified and observed every week as to its flowering stage. The observations started when the earlier genotypes were at the booting stage and extended until all the labeled plants reached maturity. These weekly observations were used to estimate the flowering period of all the labeled tillers. At maturity, each identified tiller had its panicle harvested and evaluated for degree of exsertion. The panicles of the parents were also evaluated for percentage of spikelet fertility. This trait was not evaluated in the F2 populations due to the high sterility observed in the crosses between Indica and Japonica genotypes, which was attributed to genetic causes, rather than to low temperatures.
Degree of panicle exsertion was measured as the distance from the flag leaf ligule to the panicle node, in cm. Positive and negative values indicated complete and incomplete panicle exsertion, respectively. Spikelet fertility was obtained by counting the number of filled and empty grains and expressed as the percentage of filled grains by the total number of spikelets of the panicle.
Statistical analysis of the traits measured in the parents was performed considering a randomized block design with subsampling in the plots, with three replications and 20 subsamples per replication. Residual and regression analysis were performed to verify data normality and the need for transformation, respectively. Both degree of panicle exsertion and percentage of spikelet fertility were transformed according to √ (x + 10). The traits were submitted to analysis of variance, means comparison by Duncan's Test and calculation of Pearson's correlation coefficients, all performed by the Statistical Analysis System (SAS Institute, 2000).
The inheritance of degree of panicle exsertion was studied through an analysis of the distribution of frequencies at the generations P1, P2 and F2 in each of the crosses, which were obtained by grouping the data into classes, with the class interval corresponding to ¼ of the average standard deviation (Steel & Torrie, 1980).
Variances were estimated as follows (Allard, 1960): VP = VF2; VE = (VP1 + VP2) / 2; VG = VP VE
where: VP1 and VP2 represent the variances of the parents and VF2 represent the variance of the F2 generation. Heritability estimates in the broad sense were calculated according to the formula (Allard, 1960):
hb2 = (VP VE) / VP
RESULTS AND DISCUSSION
Flowering in the parents and in the F2 progenies began in the second half of Feb. and extended until the end of March (data not presented). The cold tolerant genotypes Quilla 64117 and Diamante flowered approximately 20 days before the sensitive ones, BRS Taim and IRGA 417. As a consequence, there was a large segregation for growth duration in the F2 generation derived from the crosses between sensitive and tolerant genotypes. This makes selection for cold tolerance under field conditions very imprecise, once the plants were exposed to different air temperatures at flowering. To avoid this effect, only plants that flowered between Feb. 19th and March 18th were considered for evaluating the parents and for the inheritance study. Average temperature and average minimum temperature did not vary considerably within this period. Average temperature was 22ºC (±1.5) and the average minimum temperature was 17ºC (±2.0), which is below the critical limit of 20ºC for cold damage (Fepagro, 2000).
Differences were observed among the parents for panicle exsertion and percentage of spikelet fertility, indicating variability among them for these traits. The cold tolerant genotypes Quilla 64117 and Diamante had a better performance than the sensitive parents, presenting complete panicle exsertion (positive values) and higher percentage of spikelet fertility (Table 1).
The F1 and F2 generations derived from the crosses between sensitive and tolerant genotypes presented a high degree of sterility (data not shown), as they resulted from an Indica × Japonica combination. High sterility has been often observed in this type of cross and has genetic causes (Oka & Morishima, 1997; Kubo & Yoshimura, 1999; Kubo et al., 2000). In fact, in the F2 generation, segregation for spikelet fertility was clearly observed, and due to its genetic origin, evaluation of this trait as a cold tolerance parameter was not performed. So, panicle exsertion was chosen as a cold tolerance indicator because complete panicle exsertion has been associated with cold tolerance (Chung, 1979; Han et al., 2006). A moderate and posi tive phenotypic correlation (r = 0.43**) between panicle exsertion and spikelet fertility was found based on parental data obtained in the field experiment, demonstrating that larger the panicle exsertion, higher the percentage of spikelet fertility. This is in accordance with data that shows a positive correlation between panicle exsertion and seed setting in rice under cold water conditions (Han et al., 2004).
The distributions of frequency for the degree of panicle exsertion in the six crosses are presented in Figure 1. The two cold tolerant genotypes had very similar distributions, which was expected since they did not differ to degree of panicle exsertion (Table 1). In the case of the cold sensitive genotypes, although there was a certain similarity, the genotype IRGA 417 presented lower values of panicle exsertion than BRS Taim, as shown by Duncan's Test (Table 1).
Distinction between the parents could be observed in the four sensitive × tolerant crosses in which the cold sensitive parent presented predominantly negative values (incomplete panicle exsertion) and the cold tolerant parent mostly positive values (complete panicle exsertion). Panicle exsertion data for the F2 generation of these crosses revealed a continuous distribution, but the average was negative, indicating predominance of incomplete panicle exsertion such as the cold sensitive parent (Figure 1). A continuous distribution for panicle exsertion has also been reported (Bardhan Roy et al., 1982), but other studies have related that complete panicle exsertion is controlled by a dominant gene (Mahishi et al., 1991; Pandey & Gupta, 1993). In both of these studies, however, panicle exsertion was treated as a discrete variable with data computed in one of the two classes: complete vs. incomplete exsertion and analyzed by the Chi-Square Test.
There are also two other differences between those studies and the present one. First, in both studies the tolerant × sensitive type of cross was between Indica genotypes, while here they involved an Indica versus a Japonica genotype. Second, despite being also conducted under field conditions, they were performed in India, which presents environmental differences in relation to the Brazilian irrigated rice region. So the different results between this study and the ones by Mahishi et al. (1991) and Pandey & Gupta (1993) may be attributed to different genotypes and environmental conditions.
Besides the continuous distribution, the transgressive segregation observed for panicle exsertion in the F2 generation points to genes complementary distributed between the parents. This transgressive segregation was towards incomplete exsertion, which was clearly dominant over complete exsertion in the crosses between sensitive and tolerant genotypes. A similar result was reported in a diallel analysis, where it was verified that the alleles that increased panicle exsertion were, in their majority, recessives (Acharya, 1987).
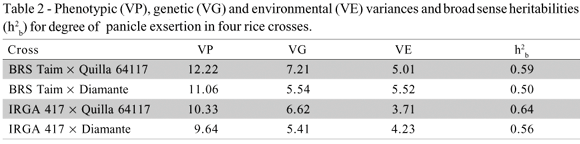
Broad sense heritabilities for the crosses between cold sensitive and cold tolerant genotypes were moderate (Table 2), and the crosses involving Quilla 64117 presented slightly higher heritabilities than the ones in which Diamante was the cold tolerant parent.
In the process of breeding for cold tolerant rice genotypes at the reproductive period, under the Southern Brazilian field conditions, selection for complete panicle exsertion is recommended as an indicative of cold tolerance, once it increases spikelet fertil ity (Chung, 1979). However, considering the results here obtained, temperature variation in the field and the fact that plants do not flower at the same time in a segregating population, it is advisable to select for panicle exsertion in more advanced generations of the breeding program. In this case, plants will have already been selected for adequate growth duration and so they will flower mostly at the same period, lowering the environmental variance. An association between plant stature and panicle exsertion has been reported (Bardhan Roy et al., 1982) and should also be taken into account, with selection for taller plants recommended in order to get better panicle exsertion. In the present study, the two genotypes with complete panicle exsertion are indeed taller than the other two genotypes, which was already expected as they are Japonicas.
In fact, panicle exsertion seems to be related to many other traits in rice as demonstrated by the clustering of QTLs for panicle exsertion, plant height, panicle number, panicle length and biomass production at the same marker intervals (Hittalmani et al., 2002). Pleiotropism and/or tight linkage were suggested as the plausible reasons for the congruence of all these QTLs. If these arguments are confirmed, selection for better panicle exsertion will have a direct effect on many other traits or will be affected by selection on these traits. Also, the reported relationship between the last internode and panicle development (Sunohara et al., 1999) indicates that the genes responsible for complete panicle exsertion under low temperature are probably internode elongation genes less sensitive to chilling. The recessive eui1 gene, which controls elongation of the uppermost internode in rice and was produced by mutagenesis (Ma et al., 2004), proves the relationship among internode elongation, plant stature and panicle exsertion in rice. When compared with the wild type, plant height, internode elongation and panicle exsertion were increased significantly in the mutated genotype. QTL for culm length and affecting cold tolerance have been mapped in a Japonica population (Takeuchi et al., 2001). These results make the study of panicle exsertion more complex as an individual trait, but as mapping efforts advance, more about the genetic control of plant stature, internode elongation and panicle exsertion will be revealed, which will contribute to our knowledge of the relationship among them.
Received December 11, 2006
Accepted April 14, 2008
- ACHARYA, S. Genetic parameters and their implication in breeding cold tolerant varieties of rice (Oryza sativa L.). Crop Improvement, v.14, p.100-103, 1987.
- ALLARD, R.W. Principles of plant breeding New York: John Wiley, 1960. 485p.
- ALVARADO, J.R.; GRAU, P.B. Mejoramiento del arroz en Chile por tolerancia a frío. In: Reunión sobre Mejoramiento de Arroz en el Cono Sur, Montevideo, 1991. Proceedings Montevideo: IICA-Procisur, 1991. p.105-114.
- BARDHAN ROY, S.K.; PATEÑA, G.F.; VERGARA, B.S. Feasibility of selection for traits associated with cold tolerance in rice under rapid generation advance method. Euphytica, v.31, p.25-31, 1982.
- CHUNG, G.S. The rice cold tolerance program in Korea. In: RICE COLD TOLERANCE WORKSHOP, Los Baños, 1979. Report Los Baños: International Rice Research Institute, 1979. p.7-19.
- FUNDAÇÃO ESTADUAL DE PESQUISA AGROPECUÁRIA - FEPAGRO. Banco de Dados do Centro de Meteorologia Aplicada Porto Alegre: FEPAGRO, 2000. Disquete.
- HAMDANI, A.R. Low-temperature problems and cold tolerance research activities for rice in India. In: RICE COLD TOLERANCE WORKSHOP, Los Baños, 1979. Report Los Baños: International Rice Research Institute, 1979. p.39-48.
- HAN, L.; YUAN, D.; XUAN, Y.; PIAO, Z.; KOH, H. Genetic analysis of cold water response on several agronomic traits of rice. Chinese Journal of Rice Science, v.18, p.23-28, 2004.
- HAN, L.; QIAO, Y.; ZHANG, S.; CAO, G.; YE, C.; XU, F.; DAI, L.; YE, J.; KOH, H. QTL analysis of some agronomic traits in rice under different growing environments. Agricultural Sciences in China, v.5, p.15-22, 2006.
- HITTALMANI, S.; SHASHIDHAR, H.E.; BAGALI, P.; HUANG, N.; SIDHU, J.S.; SINGH, V.P. KHUSH, G.S. Molecular mapping of quantitative trait loci for plant growth, yield and yield related traits across three diverse locations in a doubled haploid rice population. Euphytica, v.125, p.207-214, 2002.
- JACOBS, B.C.; PEARSON, C.J. Cold damage and development of rice: a conceptual model. Australian Journal of Experimental Agriculture, v.34, p.917-919, 1994.
- KUBO, T.; YOSHIMURA, A. Complementary genes causing F2 sterility in Japonica/Indica cross of rice. Rice Genetics Newsletter, v.16, p.68-70, 1999.
- KUBO, T.; EGUCHI, M.; YOSHIMURA, A. A new gene for F1 pollen sterility in Japonica/Indica cross of rice. Rice Genetics Newsletter, v.17, p.63-64, 2000.
- MA, H.; ZHANG, S.; LU, Q., FANG, X.; YANG, S.; YANG, R. Genetic analysis and molecular mapping of eui1 gene conferring elongated uppermost internode in the indica rice mutant Xieqingzao eB1. Chinese Journal of Agricultural Biotechnology, v.1, p.103-107, 2004.
- MAHISHI, D.M.; MAHADEVAPPA, M.; REDDY, P.G. Inheritance of panicle exsertion in Oryza sativa under low temperature. In: International Rice Genetics Symposium, 2., Manila, 1990. Proceedings. Los Baños: International Rice Research Institute, 1991. p.175-177.
- NANDA, J.S.; SESHU, D.V. Breeding strategy for cold-tolerant rices. In: RICE COLD TOLERANCE WORKSHOP, Los Baños, 1979. Report Los Baños: International Rice Research Institute, 1979. p.91-99.
- OH, C.; CHOI, Y.; LEE, S.; YOON, D.; MOON, H.; AHN, S. Mapping of quantitative trait loci for cold tolerance in weedy rice. Breeding Science, v.54, p.373-380, 2004.
- OKA, H.I.; MORISHIMA, H. Origin and differentiation of cultivated rice; wild and cultivated rice. In: MATSUO, T.; citar os outros autores (Ed.) Science of the rice plant. Tokyo: Food and Agriculture Policy Research Center, 1997. v.3, p.88-111.
- PANDEY, D.K.; GUPTA, H.S. Genetics of panicle exsertion in cold tolerance rice (Oryza sativa L.). Plant Breeding, v.111, p.82-85, 1993.
- SAS INSTITUTE. System for information: version 8.0. Cary: SAS Institute, 2000.
- SOUZA, P.R. Alguns aspectos da influência do clima temperado sobre a cultura do arroz irrigado no sul do Brasil. Lavoura Arrozeira, v.43, p.9-11, 1990.
- STEEL, R.D.G.; TORRIE, J.L. Principles and procedures of statistics New York: McGraw-Hill, 1980. 631p.
- SUNOHARA, H.; UOZU, S.; KITANO, H.; citar os outros autores. Interaction of panicle development and internode elongation in rice. Rice Genetics Newsletter, v.16, p.30-32, 1999.
- TAKAHASHI, N. Differentiation of ecotypes in Oryza sativa L. In: TAKAHASHI, N.; TSUNODA, S. (Ed.) Biology of rice Tokyo: Japanese Scientific Society, 1984. p.31-67.
- TAKEUCHI, Y.; HAYASAKA, H.; CHIBA, B.; citar os outros autores. Mapping quantitative trait loci controlling cool-temperature tolerance at booting stage in temperate japonica rice. Breeding Science, v.51, p.191-197, 2001.
- YOSHIDA, S. Fundamentals of rice crop science Los Baños: International Rice Research Institute, 1981. 269p.
Publication Dates
-
Publication in this collection
16 Sept 2008 -
Date of issue
2008
History
-
Accepted
14 Apr 2008 -
Received
11 Dec 2006