Abstracts
Onion is the third most grown vegetable crop in São Paulo state, Brazil. Organic onion farming is expected to increase in the state due to the increasing demand. Pest management in organic onion farming is based on plant extracts with insecticide effects. However, the efficacy of such plant extracts has not been proved yet, and it was observed that they do negatively affect natural enemies. Plants surrounding onion fields, and that are attractive to natural enemies, may be a good option to farmers, since they may lead to increased diversity of arthropod species and, consequently, the natural control of pest populations. This study deals with the effect of marigold plants as a resource plant to natural enemies in onion fields. The experiment was set in a certified organic farm using marigold rows at a center of an onion field. Samples were taken from marigold and the onion plants 5 m (near) and 30 m (far) from the flowering strips. Higher numbers of arthropod pests were observed in onion plants 30 m from the marigold strip, while higher numbers of predators and parasitoids were found at 5 m distance. Species richness and Shannon's diversity index were higher at 5 m from marigold. Therefore, marigold rows next to onion fields resulted in higher number of entomophagous species, potentially enhancing the natural control of onion pests. In the study field, marigold strips may be an alternative to crop sprays for organic control of onion pests.
natural control; conservation; predator; parasitoid
A cebola é a terceira hortaliça mais cultivada em São Paulo, cujo cultivo orgânico tende a crescer devido ao grande mercado consumidor existente. O manejo das pragas na cebolicultura orgânica baseia-se nos extratos de plantas inseticidas que, além de terem eficiência não comprovada, podem afetar negativamente os inimigos naturais. A utilização de plantas atrativas a inimigos naturais no entorno dos cultivos de cebola pode ser uma boa opção para os produtores, pois potencialmente aumentam a diversidade da artropodofauna e podem regular naturalmente as populações de espécies pragas. Verificou-se o efeito de linhas de cravo-de-defunto (Tagetes erecta L.) como cultura atrativa a inimigos naturais em campos de cebola. O experimento foi realizado em propriedade orgânica certificada, instalando-se linhas de cravo-de-defunto nas bordaduras de dois campos de cebola. Os tratamentos foram definidos coletando-se amostras nas plantas de cebola próximas às linhas da atrativa (5m) e distante das linhas (30 m). De maneira geral, observou-se maior quantidade de artrópodes fitófagos nas plantas de cebola longe da faixa atrativa e, inversamente, maior quantidade de inimigos naturais próximos à faixa. A riqueza de espécies também foi maior nas coletas próximas, o mesmo valendo para o índice de diversidade de Shannon. Portanto, a manutenção de linhas de cravo-de-defunto próximas ao cultivo de cebola promoveu maior riqueza e diversidade de artrópodes, bem como maior número de entomófagos, resultando em menor presença de fitófagos nas plantas, auxiliando na regulação natural das pragas da cultura.
controle natural; conservação; predador; parasitóide
ENTOMOLOGY
Marigold (Tagetes erecta L.) as an attractive crop to natural enemies in onion fields
Cravo-de-defunto (Tagetes erecta L.) como cultura atrativa para inimigos naturais em cultivo de cebola
Luís Cláudio Paterno SilveiraI; Evoneo Berti FilhoII, * * Corresponding author < eberti@esalq.usp.br> ; Leonardo Santa Rosa PierreII; Fernanda Salles Cunha PeresIII; Julio Neil Cassa LouzadaIV
IUFLA - Depto. de Entomologia, C.P. 37 - 37200-000 - Lavras, MG - Brasil
IIUSP/ESALQ - Depto. de Entomologia e Acarologia, C.P. 09 - 13418-900 - Piracicaba, SP - Brasil
IIIUNESP/FCAV - Depto. de Fitossanidade - 14884-900 - Jaboticabal, SP - Brasil
IVUFLA - Depto. de Biologia
ABSTRACT
Onion is the third most grown vegetable crop in São Paulo state, Brazil. Organic onion farming is expected to increase in the state due to the increasing demand. Pest management in organic onion farming is based on plant extracts with insecticide effects. However, the efficacy of such plant extracts has not been proved yet, and it was observed that they do negatively affect natural enemies. Plants surrounding onion fields, and that are attractive to natural enemies, may be a good option to farmers, since they may lead to increased diversity of arthropod species and, consequently, the natural control of pest populations. This study deals with the effect of marigold plants as a resource plant to natural enemies in onion fields. The experiment was set in a certified organic farm using marigold rows at a center of an onion field. Samples were taken from marigold and the onion plants 5 m (near) and 30 m (far) from the flowering strips. Higher numbers of arthropod pests were observed in onion plants 30 m from the marigold strip, while higher numbers of predators and parasitoids were found at 5 m distance. Species richness and Shannon's diversity index were higher at 5 m from marigold. Therefore, marigold rows next to onion fields resulted in higher number of entomophagous species, potentially enhancing the natural control of onion pests. In the study field, marigold strips may be an alternative to crop sprays for organic control of onion pests.
Key words: natural control, conservation, predator, parasitoid
RESUMO
A cebola é a terceira hortaliça mais cultivada em São Paulo, cujo cultivo orgânico tende a crescer devido ao grande mercado consumidor existente. O manejo das pragas na cebolicultura orgânica baseia-se nos extratos de plantas inseticidas que, além de terem eficiência não comprovada, podem afetar negativamente os inimigos naturais. A utilização de plantas atrativas a inimigos naturais no entorno dos cultivos de cebola pode ser uma boa opção para os produtores, pois potencialmente aumentam a diversidade da artropodofauna e podem regular naturalmente as populações de espécies pragas. Verificou-se o efeito de linhas de cravo-de-defunto (Tagetes erecta L.) como cultura atrativa a inimigos naturais em campos de cebola. O experimento foi realizado em propriedade orgânica certificada, instalando-se linhas de cravo-de-defunto nas bordaduras de dois campos de cebola. Os tratamentos foram definidos coletando-se amostras nas plantas de cebola próximas às linhas da atrativa (5m) e distante das linhas (30 m). De maneira geral, observou-se maior quantidade de artrópodes fitófagos nas plantas de cebola longe da faixa atrativa e, inversamente, maior quantidade de inimigos naturais próximos à faixa. A riqueza de espécies também foi maior nas coletas próximas, o mesmo valendo para o índice de diversidade de Shannon. Portanto, a manutenção de linhas de cravo-de-defunto próximas ao cultivo de cebola promoveu maior riqueza e diversidade de artrópodes, bem como maior número de entomófagos, resultando em menor presença de fitófagos nas plantas, auxiliando na regulação natural das pragas da cultura.
Palavras-chave: controle natural, conservação, predador, parasitóide
INTRODUCTION
Onion (Allium cepa L.) is the third most grown vegetable crop and the sixteenth most important organic vegetable crop in São Paulo state, occupying an area of 10,000 hectares (Camargo et al., 2006). It is largely used for food and medicinal purposes and chemical free bulbs therefore have greater value (Alonso, 1998). Because of the high market value of these products, the Brazilian growers are increasing the area of onion crops (Martins et al., 2006).
Among the insect pests causing yield losses in onion production in Brazil, the onion thrips, Thrips tabaci Lind. (Thysanoptera: Thripidae) is responsible for 45% of onion yield losses in the country (Boiça Júnior & Domingues, 1987; Sato, 1989; Gonçalves, 1996). One of the main recommendations for organic farming is the recovery of functional biodiversity, which is lost or very reduced in conventionally grown monocultures. Through polycultivation, maintenance of volunteer plant species and use of attractive plants the diversity of beneficial insects can be recovered in agroecosystems (Gliessman, 2001; Altieri et al., 2003). This diversity may lead to increased pest control in onion, reducing the use of plant extracts that have been shown to be detrimental to natural enemy populations.
A plant that is potentially useful to maintain arthropod biodiversity, including certain species of predator thrips, is the marigold (Tagetes erecta L.). According to Sampaio et al. (2008) this plant hostages species of Orius (Hemiptera: Anthocoridae), which is the main thrips predator around the world, including Brazil (Silveira et al., 2003a,b, 2004). Additionally, marigold plants in-between rows of onion crop have been shown to promote the reduction of aphid, nematode and whitefly populations and virus diseased plants (Martowo & Rohama, 1987; Abid & Magbool, 1990; Zavaleta-Mejia & Gomes, 1995). Moreover marigold plants host other phytophagous species that are alternative prey for entomophagous species. Some organic growers seed marigold for its pollen and nectar, which increase natural enemy fecundity and survival (Baggen, 1999). Therefore it is important to know the richness, abundance and diversity of arthropod species found in this plant, and on the surroundings of a strip, for example, once it may lead to important differences in species composition of a culture crop near or far from the marigold strips.
The use of this plant may contribute to management of onion pests, mainly the thrips, because the increase of the number of predators would promote lowering costs in pest control, thus, turning the organic onion crop more sustainable. Moreover, this study contributes to the identification of insect species occurring on both plants, providing basic information for further researches.
The aim of this study was to determine the effect of marigold plants near or far from organic onion cultivation for maintenance of natural enemies, and study the arthropod composition and its abundance on onion and marigold fields.
MATERIAL AND METHODS
The experiment was set in an organic onion field of 5,000 square meters, in a certified organic farm (certified by 'Mokiti Okada Foundation', FMO), in Fernando Prestes, state of São Paulo, Brazil (21º14'0.29"S; 48º40'15.61"W), from August to October 2004. Seedlings, compost and biofertilizer were produced at the farm, and an irrigation system was used. Marigold seed was obtained directly from field collection on the organic farm selected and because the grower has cultivated it for years, there is no way to determine the cultivar.
The onion field has 50 × 100 m and was divided in two similar areas by a 4 m wide path. Along the borders of this path, on both sides, there was a 50 m long and 0.5 m wide row of marigold plants. Such arrangement allowed the collection of onion samples at varying distances from marigold plants. We sampled onion plants at 5 m (near) 30 m (far) and in the marigold plant strip. Sampling began two weeks after transplanting when seedlings were 30 day-old, and continued for the following seven weeks. Ten random samples on onion according to each distance were taken weekly for seven weeks (n = 140), each sample corresponding to 1 m2 of the onion field.
Onion plants were shaken against a white plastic tray to dislodge arthropods, which were immediately transferred to plastic containers and kept in 60% ethanol for later identification. Arthropods were identified to the lowest possible level and grouped as phytophagous or entomophagous. We performed an ANOVA with the average calculated over seven weeks of sampling, showing patterns of weekly abundance but performed statistical analysis for the whole period of the experiment. We used the software Statistica® for the analysis and SigmaPlot® for graphics.
The marigold strips were also sampled during the period of onion cultivation, and each sample corresponding to one square meter of crop coverage, using the same methodology as for onion plants. These marigold samples were taken randomly and at three developmental stages during the experiment: a) onset flowering, when up to 10% of plants were flowering (25 August and 7 September), plants were in full vegetative growth with few flowers (N = 8); b) peak flowering stage (N = 8), when more than 80% of the plants were flowering (8 and 15 September); c) late flowering, when flowers were senescent and losing their attractiveness (29 September and 6 October), and the number of plants were lower, since they were dying (N = 6).
The following ecological parameters were determined for onion and marigold using the software Biodiversity Pro® (McAleece, 1997): (i) Richness of species (the total number of species and morph species collected); abundance index (Lambshead et al., 1983), calculated from the mean of the entire 7-week sampling season of each species; (ii) Diversity index H' (Shannon & Weaver, 1949), which considers the quantitative uniformity of each species in relation to the others; (iii) Similarity index, calculated through cluster analysis (Pielou, 1984), which indicates how similar two substrates can be in relation to the species found. In this case the data were separated in two groups, phytophagous or entomophagous, and the whole sampling periods were considered for each substrate; and (iv) Correlation index (R2) (Sokal & Rohlf, 1969), which shows the dependence between two groups of data obtained from different plants.
Besides these indices, collection curves were calculated for onion samples (Magurran, 1988). These curves may indicate whether sampling was regular and enough to potentially collect all species that may be present in the crop (for the whole experimental period, independently of the sampling week).
For the determination of these parameters, arthropods presenting very low occurrence (one or two individuals) during the entire sampling period were not included in analysis.
RESULTS AND DISCUSSION
First of all, the calculated collection curve indicated that the 140 samples taken from onion crop were enough to collect the majority of species present in the crop during the experiment. After the sampling 134 the collection curve reaches the asymptotic position, indicating that no new species were collected from this point on.
Data from onion samples were taken at 5 m and 30 m from the marigold strip (Figure 1). Arthropods were grouped in two main ecological groups: phytophagous and entomophagous, considering all specimens found on samples, regardless of their higher or lower abundance. There was a higher number of phytophagous arthopods m-2 in onion plants far from marigold (30 m). Despite the higher variation, the number of entomophagous arthopods m-2 may explain this result, as the number of natural enemies was lower in plants localized far from marigold row, therefore resulting in higher amounts of herbivores. In this field there is a positive effect of marigold rows on natural enemy abundance and negative effect on pest insect abundance. Similar results were reported by Driutti (1998) using several plant species in surrounding rows and within organic onion fields, who found that the number of phytophagous insects, especially thrips, which account for more than 86% of the herbivores, was significantly lower near the attractive plants.
Another possible explanation for this result is that marigold plants could produce repellent volatiles to thrips, including T. tabaci. So, there would be less thrips near marigold strips because of its repellence, not by natural enemy predation. As stated by Visser (1986), phytophagous insects respond to a complex mixture of plant odors to recognize their hosts. Four plant essential oils to repeal T. tabaci were tested by Tol et. al. (2007), who conclude that Origanum majorana L. (Labiatae) is a promising thrips repellent which could be used for further testing in a pushpull system. Similar tests should be performed to identify this possibility in relation to T. erecta.
The analysis of ecological indices on grouped data from onion (5 m and 30 m from the marigold strip, N = 70 for each) and marigold rows (N = 8 for onset flowering stage and peak flowering stage and N = 6 for late flowering stage) (Table 1) showed higher richness in samples near the marigold strip, with 37 taxa or morphospecies, and 24 species in samples far from the marigold strip. The diversity index (H') was higher at 5 m from the marigold strip, indicating that dominance of species is slightly lower than at 30 m from the marigold strip. This is due to a higher number of T. tabaci individuals observed at 30 m than at 5 m from marigold.
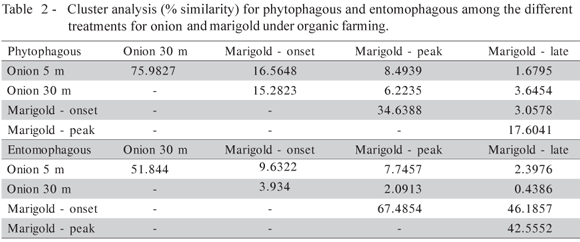
In general, there was high similarity for phythophagous (75.98 through cluster analysis, Table 2) and correlation (R2 = 0.9985) between species collected in onions at 5 m and at 30 m from the marigold strip, thus indicating high relation between the amount of each species in different distances. For natural enemies these values are low (51.84% similarity, Table 2, and R2 = 0.9063). In spite of these similarities, the main phytophagous species collected, T. tabaci, was 57% more abundant far from marigold than near it. A similar result was found for the second most abundant species, Therioaphis trifolii (Hemiptera: Aphidae), which was 15% more abundant far from the marigold strip. The opposite was observed with predators, especially Stomatothrips angustipennis, Stomatothrips rotundus and Franklinothrips vespiformis (Thysanoptera: Aeolothripidae), which were twice as abundant in onion plants near the marigold strip. Parasitoid abundance, in general, was very low in onion plants, but much lower far from the marigold strip (Table 1). Regarding marigold plants (Table 1) we found more richness and abundance of parasitoids compared to onion plants, since those plants produce pollen and nectar that potentially can sustain parasitoid populations (Baggen, 1999).
Very low percentages of similarity were observed between phytophagous and entomophagous on onion compared to marigold (Table 2). For herbivores the values are similar in relation to the distance from marigold, but become lower as the flowering stage advance. That was expected since the main herbivore species, T. tabaci, was not observed on marigold at peak and late flowering stage. For natural enemies the similarity was even lower compared to herbivores, but the distance to marigold strip affect the values, since they were higher if compared onion at 5 m with onion at 30 m from the attractive plants. So these low natural enemies' similarities between marigold and onion indicate that not the same but different species were found on onion near attractive plants. Therefore, the main phytophagous species were more abundant in onion plants 30 m from marigold, while the main predators were more abundant in plants close to the attractive species. However, Gonçalves & Souza-Silva (2003), studying only T. tabaci and Toxomerus spp. (Diptera: Syrphidae) populations in relation to different plant species in rows surrounding the onion field, observed no population decrease of the pest species. This study, however, only included abundance of thrips nymphs and syrphid larvae, and there may have been differences in the abundance of other phytophagous and predatory species.
Higher herbivore species richness but lower abundance were detected in plots near marigold than in plots far from marigold. Nineteen taxa 5 m from marigold and 15 taxa 30 m from onion plants were found. Phytophagous insect abundance, however, was greater far from than near to marigold strips, 41.99 and 27.57 herbivores / sample, respectively. Both greater abundance and higher species richness of entomophagous species (predators and parasitoids) were found in plots near marigold than in plots far from marigold. Eighteen taxa 5 m from marigold and only ten 30 m from marigold plants were found, as well as nearly twice as many natural enemies near to as far from marigold strips, 0.91 and 0.46 insects / sample, respectively. Hence, we found higher diversity of both phytophagous and entomophagous species on onion plants close to marigold rows, but increased herbivore abundance and decreased natural enemy abundance on onion plants at greater distances from marigold. For the samples on marigold strips, species dynamics varied depending on the developmental stage, abundance and similarity (Tables 1 and 2), but not for richness: the same number of taxa (40) was collected considering all arthropod categories. While no numerical difference was observed, species composition varied greatly between flowering stages. The analysis of similarity indices between marigold developmental stages (Table 2) indicated that there is greater similarity between the onset and peak flowering periods for phytophagous (34.6%) than between the late flowering and the other stages (3.05 to 17.6%). For the entomophagous the similarities is bigger (Table 2) ranging between 42 and 67.5% depending on the flowering stage. This indicates that a major number of natural enemy species uses the marigold strip during more time than the herbivores do, witch is interesting for biological control.
The higher diversity index was observed in marigold during full bloom (H', Table 1), when the predator Orius insidiosus was most abundant, as well as herbivore Aphis fabae Scopoli (Hemiptera: Aphididae). This is in accordance with previous observations. There was a slight dominance of phytophagous species at the beginning and at the end of flowering periods (the aphid A. fabae and the Lygaeidae Nysius sp., respectively), resulting in lower diversity indices at these stages. The marigold at the onset of flowering was more attractive to sucking phytophagous species, e.g. Hemiptera from the families Aphidae, Psyllidae and Membracidae, which combined represented over 60% of the collected individuals. This was expected because plants were still undergoing vegetative growth, i.e. sprouting and producing flowers, which were attractive to these species. At full flowering period, Aphidae represented only 29% of phytophagous species, when thrips (Thysanoptera) became dominant, with over 52% of the phytophagous and 34% of the entire arthropod abundance at this stage. This tendency was confirmed at the end of the flowering period, when thrips abundance increased to 55% of the total arthropods. Besides these phytophagous species, the sucking hemipteran Nysius sp. became very abundant in marigold flowers at this stage.
This dominance of phytophagous taxa in the beginning and the end of marigold flowering periods, however, did not produce a great impact on the onion field, because insects using marigold as a host plant (e.g. aphids, thrips or other phytophagous bugs) are not the same as those that attack onion plants, especially T. tabaci, the primary onion pest (Table 1). This is confirmed by the similarity index between onion and marigold collections (Table 2), which reached 16.56% between onion near marigold and the initial marigold development stage. This is an indication that few phytophagous species were found at both plant species, except for A. fabae and T. trifolli, which were collected in marigold and onion.
This study was set in a particular situation and therefore the data would probably not fit the expectations of other people working on organic agriculture in other conditions. Thus, further tests could be necessary to clarify the role of marigold plants near the onion crop in other areas. Anyway the data of the association of marigold plants and onion crops will certainly help organic onion growers mainly in reducing the production costs.
We found evidence that marigold strip is supporting alternative prey and hosts, mainly from full blossom to the end of the flowering period, which are key components of conservation biological control. This fact is very important to maintain local populations of generalist predators and parasitoids (Landis et al., 2000). For example, O. insidiosus abundance was higher during late flowering, possibly in response to the increase in thrips number at that time. Since the phytophagous species involved are not potential pests to onion plants, their population maintenance in the system is not likely to have the associated negative impact of increasing crop herbivore.
Therefore, keeping marigold rows close to the onion field results in higher diversity and abundance of natural enemies than when the attractive species is kept far from onion plants. This indicates a beneficial effect of marigold on the onion field, especially on the dynamics of the arthropod species, which varies as onion plants are collected from more distant marigold plants. Additional studies are necessary to determine the least distance between onion crop and the marigold rows and what is the best width of these rows.
ACKNOWLEDGEMENTS
To Mr. Ulisses Murakami for his collaboration, to FAPESP for the scholarship granted to the first author (Process no. 2003/01497-4) and financial support to this study (Process no. 2003/09819-0), as well as to Valmir A. Costa (Instituto Biológico de Campinas) for parasitoid identification.
Received December 17, 2007
Accepted June 10, 2009
- ABID, M.; MAGBOOL, M.A. Effects of inter-cropping of (Tagetes erecta) on root-knot disease and growth of tomato. International Nematology Network Newsletter, v.7, p.41-42, 1990.
- ALONSO, J.R. Tratado de fitomedicina: bases clínicas e farmacológicas. Buenos Aires: ISIS Ediciones S.R.L., 1998. 1039p.
- ALTIERI, M.A.; SILVA, E.N.; NICHOLLS, C.I. O papel da biodiversidade no manejo de pragas Ribeirão Preto: Editora Holos, 2003. 226p.
- BAGGEN, L.R.; GURR, G.M.; MEATS, A. Flowers in tri-trophic systems: mechanisms allowing selective exploitation by insect natural enemies for conservation biological control. Entomologia Experiementalis et Applicata, v.91, p.155-161, 1999.
- BOIÇA JÚNIOR, A.L.; DOMINGUES, E.P. Efeito de dimetoato, monocrotofós e carbaryl no controle de Thrips tabaci Lindeman, 1888, em duas variedades de cebola. Científica, v.15, p.67-77, 1987.
- CAMARGO, A.M.M.P.; CASER, D.V.; CAMARGO-FILHO, W.P.; CAMARGO, F. P.; COELHO, P.J. Área cultivada com agricultura orgânica no Estado de São Paulo. Informações Econômicas, v.36, p.33-62, 2006.
- DRIUTTI, A.A. Controle biológico de tripes na cultura da cebola pelo cultivo de bordaduras ou entrelinhas. Florianópolis: UFSC, 1998. 61p. Dissertação (Mestrado).
- GLIESSMAN, S.R. Agroecologia: processos ecológicos em agricultura sustentável. 2 ed. Porto Alegre: Editora Universidade/UFRGS, 2001. 653p.
- GONÇALVES, P.A.S. Determinação de danos de Thrips tabaci Lind. em cultivares de cebola. Pesquisa Agropecuária Brasileira, v.31, p.173-179, 1996.
- GONÇALVES, P.A.S.; SOUZA-SILVA, C.R. Efeito de espécies vegetais em bordadura em cebola sobre a densidade populacional de tripes e sirfídeos predadores. Horticultura Brasileira, v.21, p.731-734, 2003.
- LANDIS, D.A.; WRATTEN, S.D.; GURR, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology, v.45, p.175-201, 2000.
- LAMBSHEAD, P.J.D.; PLATT, H.M.; SHAW K.M. Detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. Journal of Natural History, v.17, p.859-874, 1983.
- McALEECE, N.J.; LAMBSHEAD; G. PATERSON; J. GAGE. Biodiversity professional: Beta Version. London: The Natural History Museum and the Scottish Association for Marine Science, 1997.
- MAGURRAN, A.E. Ecological diversity and its measurement London: Chapman & Hall, 1988. 179p.
- MARTINS, V.A.; CAMARGO FILHO, W.P.; BUENO, C.R.F. Preços de frutas e hortaliças da agricultura orgânica no mercado varejista da cidade de São Paulo. Informações Econômicas, v.36, p.42-52, 2006.
- MARTOWO, B.; ROHANA, D. The effect of intercropping of pepper (Capsicum annum L.) with some vegetable crops on pepper yield and disease incidence caused by Meloidogyne spp. Buletin Penelitian Hortikultura, v.15, p.55-59, 1987.
- PIELOU, E.C. The Interpretation of ecological data: a primer on classification and ordination. New York: John Wiley, 1984. 263p.
- SAMPAIO, M.V.; BUENO, V.H.P. ; SILVEIRA, L.C.P. ; AUAD, A.M.. Biological control of insect pests in the Tropics. In: DEL CLARO, K. (Ed.) Encyclopedia of Life Support Systems (EOLSS). Oxford: EOLSS Publishers, 2008.
- SATO, M.E. Avaliação do dano e controle do Thrips tabaci Lindeman, 1888 na cultura da cebola (Allium cepa L.). Piracicaba: USP/ESALQ, 1989. 93p. Tese (Doutorado).
- SHANNON, C.E.; WEAVER, W. The mathematical theory of communication Urbana: University of Illinois Press, 1949. 144p.
- SILVEIRA, L.C.P; BUENO, V.H.P; MENDES, S.M. Record of two species of Orius Wolff (Hemiptera, Anthocoridae) in Brazil. Revista Brasileira de Entomologia, v. 47, p.303-306, 2003a.
- SILVEIRA, L.C.P; BUENO, V.H.P; PIERRE, L.S.R.; MENDES, S.M. Plantas cultivadas e invasoras como habitat para predadores do gênero Orius (Wolff) (Heteroptera: Anthocoridae). Bragantia, v.62, p.261-265, 2003b.
- SILVEIRA, L.C.P; BUENO, V.H.P; LENTEREN, J.C. van. Orius insidiosus as biological control agent of thrips in greenhouse chrysanthemums in the tropics. Bulletin of Insectology, v.57, p.103-109, 2004.
- SOKAL, R.R.; F.J. ROHLF. Biometry: the principles of statistic in biological research. San Francisco: W.H. Freeman, 1969. 776p.
- TOL, R.W.H.M. van; JAMES, D.E.; DE KOGEL, W.J.; TEULON, D.A.J. Plant odours with potential for a push-pull strategy to control the onion thrips, Thrips tabaci Entomologia Experimentalis et Applicata, v.122, p.69-76, 2007.
- ZAVALETA-MEJÍA, E.; GOMEZ, R.O. Effect of Tagetes erecta L.-tomato (Lycopersicon esculentum Mill.) intercropping on some tomato pests. Fitopatologia Brasileira, v.30, p.35-46, 1995.
Publication Dates
-
Publication in this collection
03 Dec 2009 -
Date of issue
Dec 2009
History
-
Accepted
10 June 2009 -
Received
17 Dec 2007