ABSTRACT
A shift towards declining azoxystrobin sensitivity has been previously documented in a collection of Brazilian Monilinia fructicola isolates, but information on the stability of this trait after in vitro sub-culturing is not available. In this study, one highly-sensitive isolate and two isolates with reduced sensitivity to azoxystrobin, not exhibiting point mutations at codons 129, 137 and 143 in the target gene of cytochrome b were used. Two independent experiments consisted of 10 weekly transfers of each strain in potato dextrose agar (PDA). Conidial production and germination were quantified in the initial culture and, again, after the third, sixth and tenth transfer. Measures of mycelial growth were obtained in colonies growing on PDA amended with azoxystrobin at 1 µg mL−1 and salicylhydroxamic acid at 100 µg mL−1. Data were normalized to mycelial growth rate after each of the 10 transfers. The colony diameter in the less sensitive isolates was stable across the transfers in the PDA amended with the fungicide. The conidial production and mycelial growth rate did not decrease in non-amended media either. Our results suggest that the trait of reduced sensitivity to azoxystrobin is stable under no selection pressure for azoxystrobin.
Prunus; Quinone-outside inhibitor
Introduction
Monilinia fructicola is the main causal agent of brown rot of peach in Brazil (Martins et al., 2005Martins, M.C.; Betti, J.A.; Leite, R.M.V.B.C.; Leite, R.P.; Amorim, L. 2005. Stone fruit diseases = Doenças das rosáceas de caroço. p. 545-557. In: Kimati, H.; Amorim, L.; Rezende, J.A.M.; Bergamin Filho, A.; Camargo, L.E.A., eds. Plant Disease Compendium: Diseases in crops = Manual de fitopatologia: doenças das plantas cultivadas. Agronômica Ceres, São Paulo, Brazil (in Portuguese).; May De Mio et al., 2014May De Mio, L.L.; Garrido, L.R.; Ueno, B.; Fajardo, T.V.M. 2014. Peach diseases and control methods = Doenças da cultura do pessegueiro e métodos de controle. p. 355-432. In: Raseira, M.C.B.; Pereira, J.F.M.; Carvalho, L.C., eds. Peach Tree = Pessegueiro. Embrapa, Brasília, DF, Brazil (in Portuguese).). To manage the disease, São Paulo State growers spray triazoles alternated with strobilurin fungicides (Amiri et al., 2010Amiri, A.; Brannen, P.M.; Schnabel G. 2010. Reduced sensitivity in Monilinia fructicola field isolates from South Carolina and Georgia to respiration inhibitor fungicides. Plant Disease 94: 737-743.; May De Mio et al., 2011May De Mio, L.L.; Luo, Y.; Michailides, T.J. 2011. Sensitivity of Monilinia fructicola from Brazil to tebuconazole, azoxystrobin, and thiophanate-methyl and implications for disease management. Plant Disease 95: 821-827.). The intensive use of these fungicides selects for resistance to triazoles and reduced-sensitivity to azoxystrobin in M. fructicola isolates (May De Mio et al., 2011May De Mio, L.L.; Luo, Y.; Michailides, T.J. 2011. Sensitivity of Monilinia fructicola from Brazil to tebuconazole, azoxystrobin, and thiophanate-methyl and implications for disease management. Plant Disease 95: 821-827.). Resistance instability has been reported for M. fructicola isolates with reduced sensitivity to propiconazole without a corresponding point mutation in the cytochrome P450 sterol 14α-demythylase gene (CYP51) (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.). The levels of sensitivity to propiconazole increased in M. fructicola isolates after successive transfers in vitro and under different storage procedures, such as glycerol, filter paper, mineral oil and silica gel at low or ultralow temperatures (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.; Zhu et al., 2012Zhu, F.; Bryson, P.K.; Schnabel, G. 2012. Influence of storage approaches on instability of propiconazole resistance in Monilinia fructicola. Pest Management Science 68: 1003-1009.). Two azoxystrobin-resistant M. fructicola isolates, without point mutation in the target gene cytochrome b (cyt b), were detected in China and exhibited similar behavior with increased sensitivity to the fungicide after five transfers. The EC50 values of the resistant isolates significantly decreased from 19.6 and 9.1 µg mL−1, respectively, to 0.56 µg mL−1 for both isolates (Chen et al., 2014Chen, S.N.; Shang, Y.; Wang, Y.; Schnabel, G.; Lin, Y.; Yin, L.F.; Luo, C.X. 2014. Sensitivity of Monilinia fructicola from peach farms in China to four fungicides and characterization of isolates resistant to carbendazim and azoxystrobin. Plant Disease 98: 1555-1560.).
Information about the stability of reduced sensitivity to azoxystrobin in M. fructicola is important for both methodological and practical issues (Koller et al., 1991Koller, W.; Smith, F.D.; Reynolds, K.L. 1991. Phenotypic instability of flusilazole sensitivity in Venturia inaequalis. Plant Pathology 40: 608-611.). While a decreasing sensitivity in the population could lead to increasing doses of fungicides, potential loss of resistance upon continuous fungus transfer in non-amended media should be considered in resistance and fitness studies (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.).
This study aimed to investigate whether consecutive transfers affect selected fitness-related traits and the stability of M. fructicola isolates that exhibit a reduced-sensitivity to azoxystrobin.
Materials and Methods
Two isolates (SP09-839 and SP08-345) with reduced-sensitivity to azoxystrobin and one highly sensitive isolate (PR09-638) were stored for one year on sterile filter paper at 4 °C and cultured on PDA plates. The EC50 for azoxystrobin, determined by the spore germination in water agar with increasing concentrations of azoxystrobin (from 0.05 to 3.32 μg mL−1) plus salicylhydroxamic acid (SHAM) were 1.38 and 1.54 μg mL−1, respectively for SP09-839 and SP08-345 isolates. Isolates considered sensitive to azoxystrobin were not able to grow at doses equal or greater than 0.3 μg mL−1 (May De Mio et al., 2011May De Mio, L.L.; Luo, Y.; Michailides, T.J. 2011. Sensitivity of Monilinia fructicola from Brazil to tebuconazole, azoxystrobin, and thiophanate-methyl and implications for disease management. Plant Disease 95: 821-827.) or 1 μg mL−1 (Amiri et al., 2009Amiri, A.; Brannen, P.M.; Schnabel, G. 2009. Validation of the lipbalm tube assay for evaluation of fungicide sensitivity in field isolates of Monilinia fructicola. Plant Health Progress. DOI:10.1094/PHP-2009-1118-01-RS.). The reduced sensitivity isolates exhibited conidial germination rates higher than 50 % at the azoxystrobin-discriminatory dose of 1 µg mL−1 and presented a group I intron 1166 bp downstream of codon 143 without a point mutation at codons 129, 137 and 143 in previous experiments. Five-millimeter plugs of SP09-839 and SP08-345 isolates were transferred to the center of 9 cm diameter PDA plates amended with 1 μg mL−1 of azoxystrobin and 100 µg mL−1 of SHAM (Sigma-Aldrich, St. Louis). SHAM was dissolved in 100 % methanol (Schnabel et al., 2003Schnabel, G.; Dai, Q.; Paradkar, M.R. 2003. Cloning and expression analysis of the ATP-binding cassette transporter gene MFABC1 and the alternative oxidase gene MFAOX1 from Monilinia fructicola. Pest Management Science 59: 1143-1151.). After 15 days, the reduced-sensitive isolates were transferred to azoxystrobin-free PDA media. Seven-day-old cultures of all isolates were transferred 10 consecutive times at weekly intervals to PDA and the mycelial diameter growth was measured daily. Conidial production, conidial germination and fungicide sensitivity (mycelial diameter in azoxystrobin-amended PDA) were assessed four times, at the initial culture (T0), third (T3), sixth (T6) and tenth (T10-last) transfers. Petri dishes were incubated at 22 °C in the dark. To assess conidial production, a 5-mm mycelial plug taken 1 cm away from the center of each dish was collected and vortexed with 5 mL of distilled water and 1 % of tween for 30 s. The number of conidia was counted three times for each tube using a hemocytometer. For conidial germination, three 40 µL droplets from each conidial suspension replicate were placed in a polystyrene Petri dish. After incubating for 12 h at 22 °C in the dark, conidial germination was stopped using lactoglycerol, and the conidia were examined. The percentage of conidial germination was evaluated by counting 100 conidia per droplet, and the mean of three drops was considered a single replicate. A conidium was considered germinated when the germ tube was twice or more the length of the conidium. To assess fungicide sensitivity, a 5-mm mycelial plug from the colony periphery was placed at the center of a Petri dish with PDA, 1 μg mL−1 of azoxystrobin, and 100 µg mL−1 of SHAM, and the mycelial diameters were measured after incubating for 4 days in the dark at 22 °C. The experiment was performed twice using a completely randomized design with ten plates per isolate.
Homogeneity of variances was checked for each variable with F-test for both experiments. After confirming there was no significance (p > 0.05) in the variance ratios, all further analysis was performed with pooled data. Arcsine-square root and square root transformations were applied to conidia germination and conidia production, respectively, for stabilizing variances. ANOVA was conducted to test the significance of transfers, isolates and their interaction for each variable. A t-test tested the hypothesis that mycelial diameter in fungicide-amended media did not differ between the initial culturing and after the last transfer. A linear regression model was fitted to data in the original scale. All statistical analyses were conducted using R software (version 3.2.1).
Results and Discussion
The isolate × transfer interaction was not significant (p > 0.26) for all variables, meaning that differences between isolates did not depend on the transfers. The mycelial growth rate in PDA was not affected by transfers, as the angular coefficients of the regressions were not significant (p > 0.05) (Figure 1A-C). However, the sensitive isolate (PR09-638, Figure 1A) exhibited a higher mycelial growth rate than reduced sensitivity isolates (p < 0.001, Figure 1B and C).
− Mycelial daily growth rate on non-amended media of Monilinia fructicola sensitive (PR09-638, A) and reduced sensitivity isolates (SP08-345, B, and SP09-839, C) determined in all transfers. Bars represent the mean standard errors.
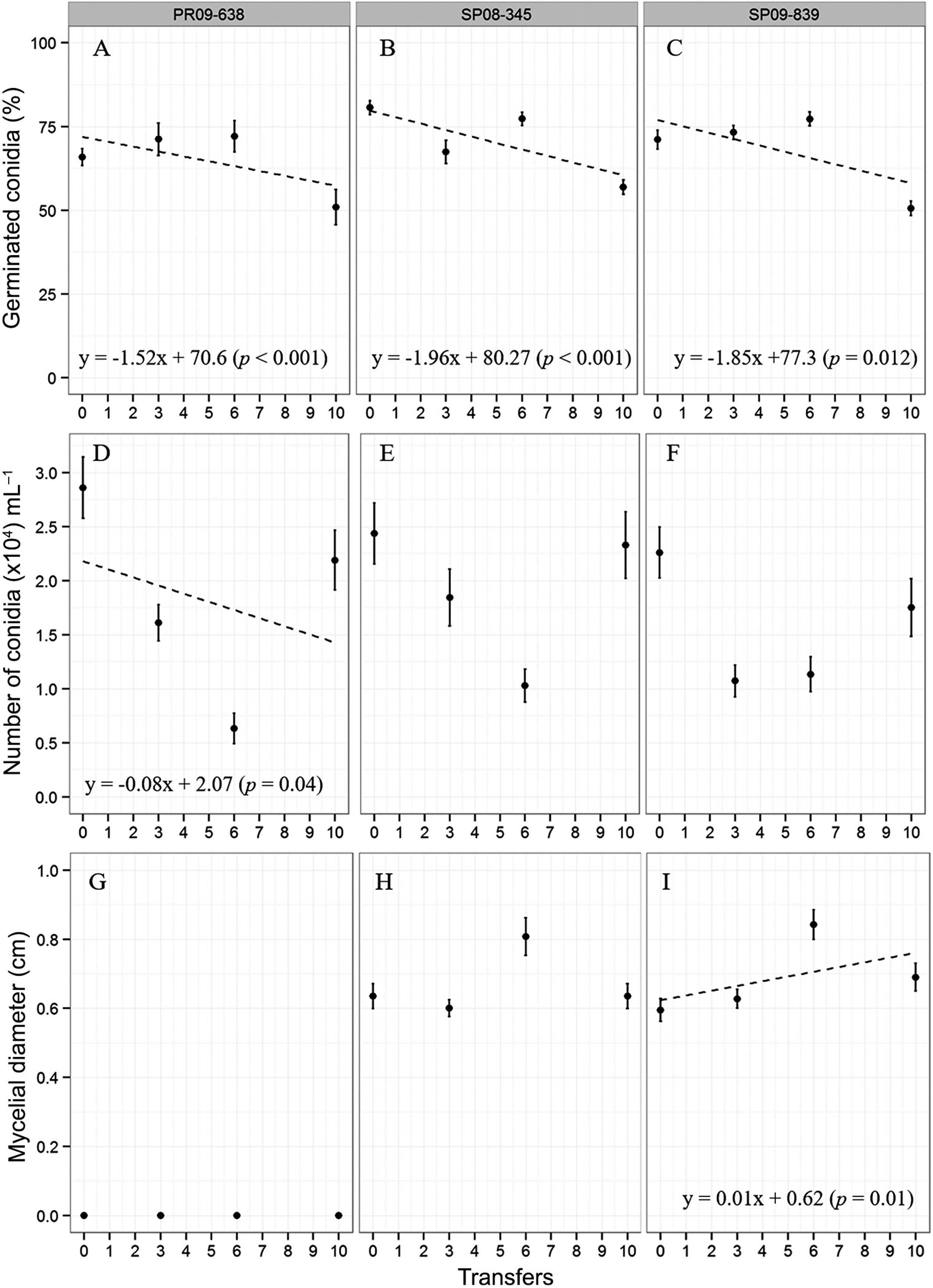
Conidial germination did not differ between the isolates (p > 0.05) but was affected by transfer (p < 0.01, Figure 2A-C). On average, conidial germination declined at a rate of 2 % during each transfer for the reduced sensitivity and sensitive isolates. Although significant (p < 0.01), this decline in conidial germination was not of major impact as the minimum germination remained above 50 % for the isolates, including the sensitive isolate (Figure 2A-C).
– Conidial germination (A, B and C) and conidial production (D, E and F) on non-amended media, and mycelial diameter on amended media with azoxystrobin at 1 µg mL−1 and salicylhydroxamic acid at 100 µg mL−1 (G, H and I) in four evaluations (the initial culture - 0 -, the third - 3 -, the sixth - 6 - and the last transfer - 10) of Monilinia fructicola PR09-638 (A, D, and G), SP08-345 (B, E, and H) and SP09-839 (C, F, and I) isolates. Bars represent the mean standard errors.
High variability in conidia production was observed across transfers for all isolates. However, considering all transfers, similar average values for conidia production were observed for all isolates (p = 0.17). Conidial production did not differ between the two reduced sensitivity isolates and was not affected by transfer (Figure 2D-F).
The level of sensitivity to azoxystrobin did not change between the initial culture and after the 10th transfer of M. fructicola isolates SP09-839 (p = 0.10) and SP08-345 (p = 0.073, Figure 2G-I). The angular coefficient of the regression was not significant for SP08-345; however, for the SP09-839 isolate a positive slope between transfers and mycelial diameter was observed (p = 0.013, Figure 2I).
An approach to estimating the stability of resistance and fitness is the comparison of two data points: the current situation (we call here “the initial culture”) with the last transfer (Bardas et al., 2008Bardas, G.A.; Myresiotis, C.K.; Karaoglanidis, G.S. 2008. Stability and fitness of anilinopyrimidine-resistant strains of Botrytis cinerea. Phytopathology 98: 443-450.; Chen et al., 2014Chen, S.N.; Shang, Y.; Wang, Y.; Schnabel, G.; Lin, Y.; Yin, L.F.; Luo, C.X. 2014. Sensitivity of Monilinia fructicola from peach farms in China to four fungicides and characterization of isolates resistant to carbendazim and azoxystrobin. Plant Disease 98: 1555-1560.; Karaoglanidis and Thanassoulopoulos, 2002Karaoglanidis, G.S.; Thanassoulopoulos, C.C. 2002. Phenotypic instability of Cercospora beticola Sacc. strains expressing resistance to the sterol demethylation-inhibiting (DMI) fungicide flutriafol after cold exposure. Journal of Phytopathology 150: 692-696.; Kim and Xiao, 2011Kim, Y.K.; Xiao, C.L. 2011. Stability and fitness of pyraclostrobin- and boscalid-resistant phenotypes in field isolates of Botrytis cinerea from apple. Phytopathology 101: 1385-1391.; Vega and Dewdney, 2014Vega, B.; Dewdney, M.M. 2014. QoI-resistance stability in relation to pathogenic and saprophytic fitness components of Alternaria alternata from citrus. Plant Disease 98: 1371-1378.), but additional data in different time point of transfers will provide a better understanding of the process (Zhan and McDonald, 2013Zhan, J.; McDonald, B.A. 2013. Experimental measures of pathogen competition and relative fitness. Annual Review of Phytopathology 51: 131-153.). There is a high degree of variability in conidia production and usually the number of conidia shows linear and gradual changes between transfers (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.; Fan et al., 2015Fan, Z.; Yang, J.H.; Fan, F.; Luo, C.X.; Schnabel, G. 2015. Fitness and competitive ability of Alternaria alternata field isolates with resistance to SDHI, Qol, and MBC fungicides. Plant Disease 99: 1744-1750.; Laleve et al., 2014Laleve, A.; Fillinger, S.; Walker, A.S. 2014. Fitness measurement reveals contrasting costs in homologous recombinant mutants of Botrytis cinerea resistant to succinate dehydrogenase inhibitors. Fungal Genetics and Biology 67: 24-36.; Vega and Dewdney, 2014Vega, B.; Dewdney, M.M. 2014. QoI-resistance stability in relation to pathogenic and saprophytic fitness components of Alternaria alternata from citrus. Plant Disease 98: 1371-1378.). This high variability has also been observed in M. fructicola isolates with reduced sensitivity to propiconazole (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.).
In most studies on QoI-resistant fungi, pathogens with a mutation at codon 143 showed, qualitatively, stable resistance following successive transfers in non-amended media, such as in azoxystrobin-resistant Magnaporthe grisea (Avila-Adame and Koller, 2003Avila-Adame, C.; Koller, W. 2003. Characterization of spontaneous mutants of Magnaporthe grisea expressing stable resistance to the Qo-inhibiting fungicide azoxystrobin. Current Genetics 42: 332-338.), Didymella bryoniae isolates (Finger et al., 2013Finger, M.J.; Stevenson, K.L.; Ji, P. 2013. Stability of azoxystrobin resistance and fitness of fungicide-sensitive and -resistant field isolates of Didymella bryoniae. Phytopathology 103: 43-44.) and pyraclostrobin-resistant Venturia inaequalis isolates (Frederick et al., 2014Frederick, Z.A.; Villani, S.M.; Cooley, D.R.; Biggs, A.R.; Raes, J.J.; Cox, K.D. 2014. Prevalence and stability of qualitative qol resistance in populations of Venturia inaequalis in the northeastern United States. Plant Disease 98: 1122-1130.). Botrytis cinerea isolates resistant to boscalid and pyraclostrobin exhibited stable resistance after as many as 20 transfers (Kim and Xiao, 2011Kim, Y.K.; Xiao, C.L. 2011. Stability and fitness of pyraclostrobin- and boscalid-resistant phenotypes in field isolates of Botrytis cinerea from apple. Phytopathology 101: 1385-1391.). However, this behavior contrasts with a study with two Chinese M. fructicola isolates with low azoxystrobin sensitivity, which showed unstable resistance after five transfers (Chen et al., 2014Chen, S.N.; Shang, Y.; Wang, Y.; Schnabel, G.; Lin, Y.; Yin, L.F.; Luo, C.X. 2014. Sensitivity of Monilinia fructicola from peach farms in China to four fungicides and characterization of isolates resistant to carbendazim and azoxystrobin. Plant Disease 98: 1555-1560.). These isolates also showed no cytochrome b mutation associated with QoI resistance. These isolates were also evaluated for stability through mycelial growth in a culture medium with fungicide; however, a lower SHAM concentration was added to the media (Chen et al., 2014Chen, S.N.; Shang, Y.; Wang, Y.; Schnabel, G.; Lin, Y.; Yin, L.F.; Luo, C.X. 2014. Sensitivity of Monilinia fructicola from peach farms in China to four fungicides and characterization of isolates resistant to carbendazim and azoxystrobin. Plant Disease 98: 1555-1560.). Although M. fructicola has low risk of resistance to QoI fungicide due to the absence of G143A point mutation (Luo et al., 2010Luo, C.X.; Hu, M.J.; Jin, X.; Yin, L.F.; Bryson, P.K.; Schnabel, G. 2010. An intron in the cytochrome b gene of Monilinia fructicola mitigates the risk of resistance development to QoI fungicides. Pest Management Science 66: 1308-1315.), the shift toward declining azoxystrobin sensitivity should be considered in further studies.
Propiconazole-resistant M. fructicola isolates (triazoles chemical group) showed instability in resistance after successive transfer in vitro, but no differences were observed in germination, mycelial growth and sporulation (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.; Zhu et al., 2012Zhu, F.; Bryson, P.K.; Schnabel, G. 2012. Influence of storage approaches on instability of propiconazole resistance in Monilinia fructicola. Pest Management Science 68: 1003-1009.). For the M. fructicola isolates, propiconazole resistance was not related to mutations in the target gene but rather with overexpression of the gene due to the ‘Mona’ element near this gene (Cox et al., 2007Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola. Phytopathology 97: 448-453.; Luo and Schnabel, 2008Luo, C.X.; Schnabel, G. 2008. The cytochrome p450 lanosterol 14 alpha-demethylase gene is a demethylation inhibitor fungicide resistance determinant in Monilinia fructicola field isolates from Georgia. Applied and Environmental Microbiology 74: 359-366.). Apparently, this overexpression produces an adaptive cost to the pathogen if the fungus is only resistant to propiconazole. However, it has been reported that M. fructicola isolates resistant to both thiophanate-methyl and propiconazole were not affected by fitness penalties (Chen et al., 2013Chen, F.; Liu, X.; Schnabel, G. 2013. Field strains of Monilinia fructicola resistant to both MBC and DMI fungicides isolated from stone fruit orchards in the eastern United States. Plant Disease 97: 1063-1068.).
In conclusion, this study showed that transferring M. fructicola isolates in culture media without azoxystrobin did not affect their sensitivity nor cultural characteristics. These experiments are critical to the maintainance of isolates with reduced sensitivity to azoxystrobin in the laboratory. It may also have important implications for the management of brown rot in Brazil over the long term as the decreasing sensitivity of isolates could lead to increasing doses of fungicide to control this disease.
Acknowledgements
This research was funded by the São Paulo State Foundation for Research Support (FAPESP - grant number 2013/20285-0). The opinions, assumptions, conclusions and recommendations expressed in this article are those of the authors and do not necessarily reflect the views of FAPESP.
References
- Amiri, A.; Brannen, P.M.; Schnabel, G. 2009. Validation of the lipbalm tube assay for evaluation of fungicide sensitivity in field isolates of Monilinia fructicola Plant Health Progress. DOI:10.1094/PHP-2009-1118-01-RS.
- Amiri, A.; Brannen, P.M.; Schnabel G. 2010. Reduced sensitivity in Monilinia fructicola field isolates from South Carolina and Georgia to respiration inhibitor fungicides. Plant Disease 94: 737-743.
- Avila-Adame, C.; Koller, W. 2003. Characterization of spontaneous mutants of Magnaporthe grisea expressing stable resistance to the Qo-inhibiting fungicide azoxystrobin. Current Genetics 42: 332-338.
- Bardas, G.A.; Myresiotis, C.K.; Karaoglanidis, G.S. 2008. Stability and fitness of anilinopyrimidine-resistant strains of Botrytis cinerea Phytopathology 98: 443-450.
- Chen, F.; Liu, X.; Schnabel, G. 2013. Field strains of Monilinia fructicola resistant to both MBC and DMI fungicides isolated from stone fruit orchards in the eastern United States. Plant Disease 97: 1063-1068.
- Chen, S.N.; Shang, Y.; Wang, Y.; Schnabel, G.; Lin, Y.; Yin, L.F.; Luo, C.X. 2014. Sensitivity of Monilinia fructicola from peach farms in China to four fungicides and characterization of isolates resistant to carbendazim and azoxystrobin. Plant Disease 98: 1555-1560.
- Cox, K.D.; Bryson, P.K.; Schnabel, G. 2007. Instability of propiconazole resistance and fitness in Monilinia fructicola Phytopathology 97: 448-453.
- Fan, Z.; Yang, J.H.; Fan, F.; Luo, C.X.; Schnabel, G. 2015. Fitness and competitive ability of Alternaria alternata field isolates with resistance to SDHI, Qol, and MBC fungicides. Plant Disease 99: 1744-1750.
- Finger, M.J.; Stevenson, K.L.; Ji, P. 2013. Stability of azoxystrobin resistance and fitness of fungicide-sensitive and -resistant field isolates of Didymella bryoniae Phytopathology 103: 43-44.
- Frederick, Z.A.; Villani, S.M.; Cooley, D.R.; Biggs, A.R.; Raes, J.J.; Cox, K.D. 2014. Prevalence and stability of qualitative qol resistance in populations of Venturia inaequalis in the northeastern United States. Plant Disease 98: 1122-1130.
- Karaoglanidis, G.S.; Thanassoulopoulos, C.C. 2002. Phenotypic instability of Cercospora beticola Sacc. strains expressing resistance to the sterol demethylation-inhibiting (DMI) fungicide flutriafol after cold exposure. Journal of Phytopathology 150: 692-696.
- Kim, Y.K.; Xiao, C.L. 2011. Stability and fitness of pyraclostrobin- and boscalid-resistant phenotypes in field isolates of Botrytis cinerea from apple. Phytopathology 101: 1385-1391.
- Koller, W.; Smith, F.D.; Reynolds, K.L. 1991. Phenotypic instability of flusilazole sensitivity in Venturia inaequalis Plant Pathology 40: 608-611.
- Laleve, A.; Fillinger, S.; Walker, A.S. 2014. Fitness measurement reveals contrasting costs in homologous recombinant mutants of Botrytis cinerea resistant to succinate dehydrogenase inhibitors. Fungal Genetics and Biology 67: 24-36.
- Luo, C.X.; Hu, M.J.; Jin, X.; Yin, L.F.; Bryson, P.K.; Schnabel, G. 2010. An intron in the cytochrome b gene of Monilinia fructicola mitigates the risk of resistance development to QoI fungicides. Pest Management Science 66: 1308-1315.
- Luo, C.X.; Schnabel, G. 2008. The cytochrome p450 lanosterol 14 alpha-demethylase gene is a demethylation inhibitor fungicide resistance determinant in Monilinia fructicola field isolates from Georgia. Applied and Environmental Microbiology 74: 359-366.
- Martins, M.C.; Betti, J.A.; Leite, R.M.V.B.C.; Leite, R.P.; Amorim, L. 2005. Stone fruit diseases = Doenças das rosáceas de caroço. p. 545-557. In: Kimati, H.; Amorim, L.; Rezende, J.A.M.; Bergamin Filho, A.; Camargo, L.E.A., eds. Plant Disease Compendium: Diseases in crops = Manual de fitopatologia: doenças das plantas cultivadas. Agronômica Ceres, São Paulo, Brazil (in Portuguese).
- May De Mio, L.L.; Garrido, L.R.; Ueno, B.; Fajardo, T.V.M. 2014. Peach diseases and control methods = Doenças da cultura do pessegueiro e métodos de controle. p. 355-432. In: Raseira, M.C.B.; Pereira, J.F.M.; Carvalho, L.C., eds. Peach Tree = Pessegueiro. Embrapa, Brasília, DF, Brazil (in Portuguese).
- May De Mio, L.L.; Luo, Y.; Michailides, T.J. 2011. Sensitivity of Monilinia fructicola from Brazil to tebuconazole, azoxystrobin, and thiophanate-methyl and implications for disease management. Plant Disease 95: 821-827.
- Schnabel, G.; Dai, Q.; Paradkar, M.R. 2003. Cloning and expression analysis of the ATP-binding cassette transporter gene MFABC1 and the alternative oxidase gene MFAOX1 from Monilinia fructicola Pest Management Science 59: 1143-1151.
- Vega, B.; Dewdney, M.M. 2014. QoI-resistance stability in relation to pathogenic and saprophytic fitness components of Alternaria alternata from citrus. Plant Disease 98: 1371-1378.
- Zhan, J.; McDonald, B.A. 2013. Experimental measures of pathogen competition and relative fitness. Annual Review of Phytopathology 51: 131-153.
- Zhu, F.; Bryson, P.K.; Schnabel, G. 2012. Influence of storage approaches on instability of propiconazole resistance in Monilinia fructicola Pest Management Science 68: 1003-1009.
Edited by
Publication Dates
-
Publication in this collection
Mar-Apr 2017
History
-
Received
02 Dec 2015 -
Accepted
19 Apr 2016