ABSTRACT:
Studies that have investigated soil carbon dynamics under Mediterranean conditions are scarce and fragmented and contrasting results have often been reported. This study aimed to fill some gaps in our knowledge by: (i) determining annual dynamics of total (RS) and heterotrophic (RH) soil respiration; (ii) estimating annual cumulative RS and RH; and (iii) investigating the relationships between RS and RH and soil temperature and water content. The study was carried out in central Italy, for a plain and a hilly site, with the focus on two main cropping systems: an alfalfa-based forage system and a wheat-based rotation system. RS and RH showed different dynamics, with spatial and temporal variability across these sites. Estimated annual cumulative RS fluxes were 8.97 and 7.43 t C ha–1 yr–1 for the plain and hilly alfalfa-based sites, respectively, and 4.67 and 5.22 t C ha–1 yr–1 for the plain and hilly wheat-based sites, respectively. The RH components of RS were 4.26 and 3.52 t C ha–1 yr–1 for the plain and hilly alfalfa-based sites, respectively, and 3.89 and 2.45 t C ha–1 yr–1 for the plain and hilly wheat-based sites, respectively. A model with a combination of soil temperature and soil water content explained 43 % to 49 % and 33 % to 67 % of the annual variation of RS and RH, respectively. These findings help to extend our knowledge of Mediterranean cropping systems, although further studies are needed to clarify the effects of management practices on the modelling of soil respiration efflux.
Keywords:
Mediterranean; alfalfa; greenhouse gasses; soil carbon stock; wheat
Introduction
Loss of soil carbon (C) from agricultural practices is likely to have significant effects on atmospheric CO2 concentrations (Smith, 2008Smith, P. 2008. Land use change and soil organic carbon dynamics. Nutrient Cycling in Agroecosystems 81: 169-178.) and land degradation (Ryan et al., 2008Ryan, J.; Masri, S.; Ibrikçi, H.; Singh, M.; Pala, M.; Harris, H.C. 2008. Implications of cereal-based crop rotations, nitrogen fertilization, and stubble grazing on soil organic matter in a Mediterranean-type environment. Turkish Journal of Agriculture and Forestry 32: 289-297.). Many studies have shown that conversion of permanent vegetation to croplands can lead to decreased soil C stock (Lal, 2004Lal, R.D.A. 2004. Soil carbon sequestration impacts on global climate change and food security. Science 304: 1623-1627.; Monaci et al., 2017Monaci, E.; Polverigiani, S.; Neri, D.; Bianchelli, M.; Santilocchi, R.; Toderi, M.; D'Ottavio, P.; Vischetti, C. 2017. Effect of contrasting crop rotation systems on soil chemical and biochemical properties and plant root growth in organic farming: first results. Italian Journal of Agronomy 12: 364-374.), while conversion of cropland to permanent vegetation can increase this stock (Guo and Gifford, 2002Guo, L.B.; Gifford, R.M. 2002. Soil carbon stocks and land use change: a meta-analysis. Global Change Biology 8: 345-360.; Smith, 2008Smith, P. 2008. Land use change and soil organic carbon dynamics. Nutrient Cycling in Agroecosystems 81: 169-178.). Management practices have key roles in soil greenhouse gas emissions (Paustian et al., 2000Paustian, K.; Six, J.; Elliott, E.T.; Hunt, H.W. 2000. Management options for reducing CO2 emissions from agricultural soils. Biogeochemistry 48: 147-163.), as they directly affect the C dynamics of terrestrial ecosystems (Robertson et al., 2015Robertson, F.; Armstrong, R.; Partington, D.; Perris, R.; Oliver, I.; Aumann, C.; Crawford, D.; Rees, D. 2015. Effect of cropping practices on soil organic carbon: evidence from long-term field experiments in Victoria, Australia. Soil Research 53: 636-646.). Uncertainties have been reported in terms of soil respiration dynamics and their interactions with soil temperature (ST) and soil water content (SWC) (e.g., Feiziene et al., 2015Feiziene, D.; Janusauskaite, D.; Feiza, V.; Putramentaite, A.; Sinkeviciene, A.; Suproniene, A.; Seibutis, V.; Kadziene, G.; Deveikyte, I.; Lazauskas, S.; Janusauskaite, D. 2015. After-effect of long-term soil management on soil respiration and other qualitative parameters under prolonged dry soil conditions. Turkish Journal of Agriculture and Forestry 39: 633-651.), and with other factors, such as the soil microbial population (Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.) and soil oxygenation (Ryan and Law, 2005Ryan, M.G.; Law, B.E. 2005. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 73: 3-27.). Moreover, studies on the impact of anthropic activities on soil respiration (RS) are poorly documented (Maestre and Cortina, 2003Maestre, F.T.; Cortina, J. 2003. Small-scale spatial variation in soil CO2 efflux in a Mediterranean semiarid steppe. Applied Soil Ecology 23: 199-209.; Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.) and the available data mainly address ecosystems, where anthropogenic actions are not significant or frequent (e.g., Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.).
The Mediterranean region is considered one of the most vulnerable sites to global climate change, due to the expected prolonged drought period during summer and increasing rainfall in winter and autumn (Giannakopoulos et al., 2009Giannakopoulos, C.; Le Sager, P.; Bindi, M.; Moriondo, M.; Kostopoulou, E.; Goodess, C.M. 2009. Climatic changes and associated impacts in the Mediterranean resulting from 2 °C global warming. Global and Planetary Change 68: 209-224.). Studies that have investigated soil C dynamics under Mediterranean climate conditions are still relatively scarce (Oertel et al., 2015Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.). The lack of information on such systems represents an important challenge for the scientific community (Munoz-Rojas et al., 2012Munoz-Rojas, M.; Jordan, A.; Zavala, L.M.; De La Rosa, D.; Abd-Elmabod, S.K.; Anaya-Romero, M. 2012. Organic carbon stocks in Mediterranean soil types under different land uses (southern Spain). Solid Earth 3: 375-386.).
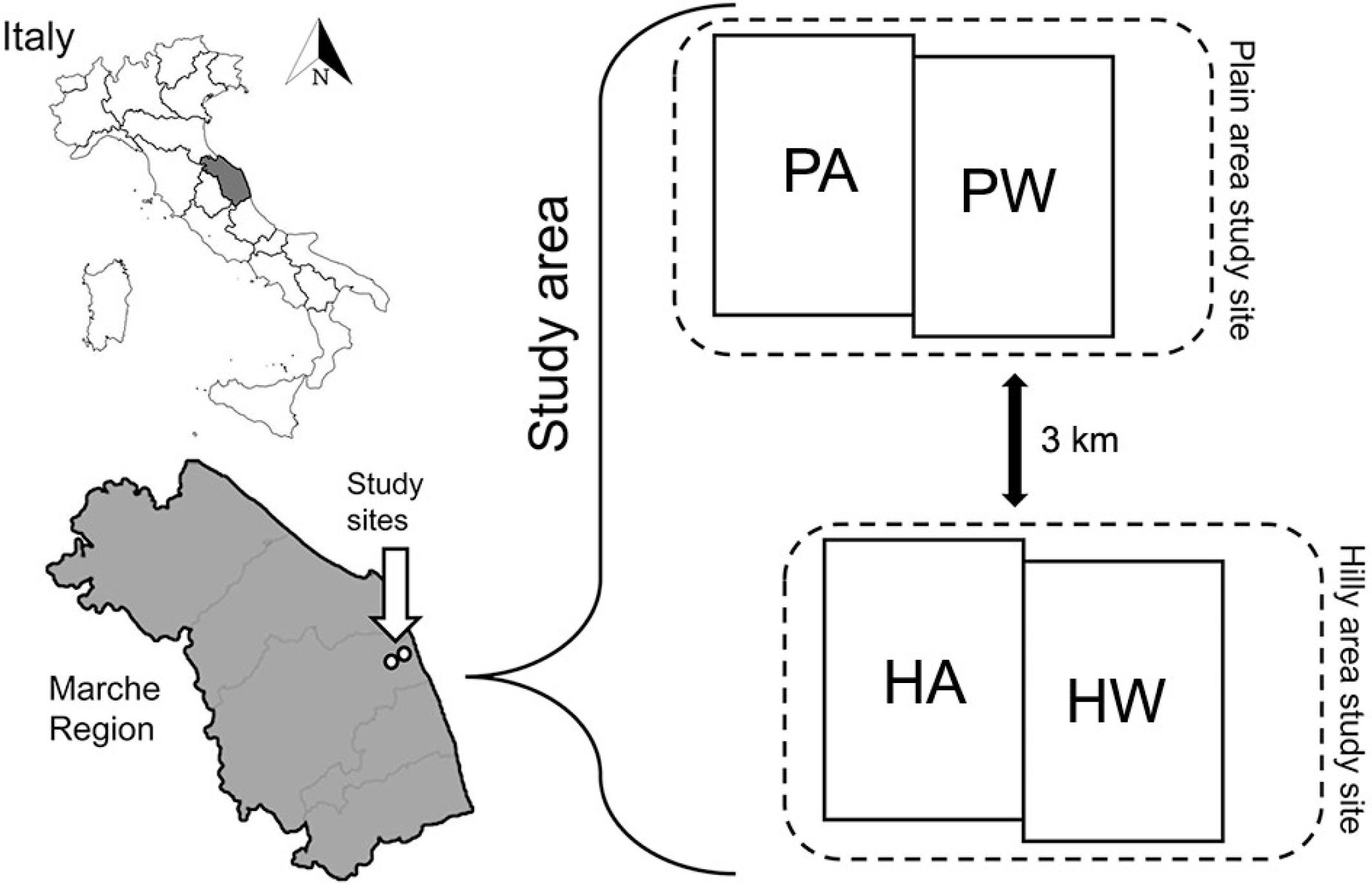
This study aims to contribute to filling gaps of knowledge on the Mediterranean climate conditions, by: (i) determining annual dynamics of RS and heterotrophic soil respiration (RH); (ii) estimating annual cumulative RS and RH; and (iii) investigating the relationships between RS, RH and ST, SWC. Two study sites were selected in central Italy in plain and hilly areas used for two main cropping systems: an alfalfa-based forage system and a wheat-based rotation system. Both sites are hereafter referred to as ‘plain alfalfa’ (PA), ‘plain wheat’ (PW), ‘hilly alfalfa’ (HA) and ‘hilly wheat’ (HW) systems.
Materials and Methods
Study sites
The study sites were located in plain and hilly areas of the Marche region (Macerata Province, central Italy) where two main cropping systems were selected, as follows:
-
An alfalfa-based forage system, where the main crop was usually kept for 3-5 years, with interruption for 1 or 2 years when winter cereals were usually cultivated. This system uses hay meadows that are sometimes grazed during the autumn-winter period by transhumant flocks (Caballero et al., 2009Caballero, R.; Fernández-González, F.; Pérez-Badia, R.; Molle, G.; Roggero, P.P.; Bagella, S.; D'Ottavio, P.; Papanastasis, V.P.; Fotiadis, G.; Sidiropoulou, A.; Ispikoudis, I. 2009. Grazing systems and biodiversity in Mediterranean areas: Spain, Italy and Greece. Pastos 39: 9-152.).
-
A typical cereal-based rotation system with winter cereals (e.g., wheat, barley), summer crops (e.g., mainly maize or sunflower), and alfalfa as a forage crop (Di Bene et al., 2016Di Bene, C.; Maretti, A.; Francaviglia, R.; Farina, R. 2016. Soil organic carbon dynamics in typical durum wheat-based crop rotations of southern Italy. Italian Journal of Agronomy 11: 209-216.). All of these crops were rain fed, except for maize, which is usually irrigated in the plain sites.
In early Nov 2014, both study sites were identified in a representative area of these cropping systems:
-
In an alluvial plain area (43°22'20.2″ N, 13°35'26.9″ E; 28 m a.s.l.), for two adjoining fields of alfalfa-based (PA) and wheat-based (PW) systems.
-
In a hilly area (43°20'40.9″ N, 13°36'19.5″ E; 120 m a.s.l.), for two adjoining fields of alfalfa-based (HA) and wheat-based (HW) systems.
The spatial distance between both study sites (i.e., from PA/PW to HA/HW) was approximately 3 km, as shown in Figure 1.
Study area and study sites. PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat.
Cropping system description and field operation
For PA and HA fields, the soil was plowed to a depth of 0.3 m before planting alfalfa, in 2010 (PA) and 2011 (HA). In central Italy, alfalfa crops do not require any particular treatments, such as fertilizers or irrigation. The mowing of alfalfa was performed two or three times a year, during late spring and summer, which depended on weather climate conditions and crop production. During the observation period, from Jan to Dec 2015, alfalfa was mowed twice, once in the first week of May 2015, and second on the first day of June 2015.
For PW and HW fields, the soil was plowed to a depth of 0.3 m in the third week of July 2014. Secondary tillage was performed in the last week of Sept for PW, and about 3 weeks earlier for HW. Wheat was sown by the end of Oct 2014 for PW, and about 1 week later for HW. The first treatments performed during the monitoring period, from Jan to Dec 2015, was fertilizer application, with 180 kg N ha–1 for PW, and 150 kg N ha–1 for HW. The fertilizer was distributed in two sessions between the second week of Feb 2015 and the end of Mar 2015. The wheat crop was harvested for both PW and HW sites in the second week of July 2015. The soil was plowed again in the first week of Aug 2015 for both PW and HW.
The previous crop rotation used for these fields was determined through interviews with farmers, shown in Figure 2.
Crop succession for experimental fields from 2006 to 2015, as derived from interviews with farmers. ‘Other crops’ include barley, pulses (e.g., pea) and vegetables (e.g., chicory). The study period was from Jan to Dec 2015. PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat.
Study site climate
The climate of the study site is Mediterranean, and during the study period (Jan to Dec 2015), the mean annual precipitation was 908.6 mm, and the mean annual temperature was 15.9 °C. The month with most rain was Oct 2015 (190.4 mm) and with least rain was July 2015 (2.4 mm). The mean air temperatures ranged from 7.3 °C (Feb 2015) to 28.0 °C (July 2015) (Figure 3).
Mean precipitation and air temperatures for the study area during the experimental period from Jan to Dec 2015. Data provided by the Agrometeorological Service of the Marche Region.
Study site soil
According to the United States Department of Agriculture (USDA) soil taxonomy (Soil Survey Staff, 2014Soil Survey Staff. 2014. Keys to Soil Taxonomy. 12ed. USDA-Natural Resources Conservation Service, Washington, DC, USA.), the soils of the study sites were classified as Inceptisol. Their basic physicochemical characteristics are shown in Table 1. Soil bulk density was determined using the cylinder method, total organic C (TOC) was determined using the Springer-Klee method, and the soil organic matter (SOM) was estimated by multiplying TOC values by the Van Bemmelen factor of 1.72 (Soil Survey Staff, 2014Soil Survey Staff. 2014. Keys to Soil Taxonomy. 12ed. USDA-Natural Resources Conservation Service, Washington, DC, USA.).
Source and basic physicochemical characteristics of soils in different fields for both study sites. Data were obtained from the analysis of five subsamples per field collected in the first 0.3 m layer. PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat.
Soil respiration analysis
Soil respiration efflux was measured in situ using a portable, closed chamber, soil respiration system that comprised an environmental gas monitor (EGM-4) with a soil respiration chamber (SRC-1). The measurement time was 120 s, according to Lai et al. (2012)Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201..
In early Nov 2014, a homogeneous area composed of six nonadjacent replicated subplots (3 m × 3 m) was identified for each of the four fields. For each subplot, a PVC collar (inner diameter, 0.10 m; length, 0.10 m; with perforated walls for the first 0.05 m) was inserted into the soil at a depth of 0.09 m. Three of the six subplots were used to measure RH, removing crop roots, and using a PVC cylinder (diameter, 0.4 m; height, 0.4 m) that was open at both ends, following the method described by Alberti et al. (2010)Alberti, G.; Delle Vedove, G.; Zuliani, M.; Peressotti, A.; Castaldi, S.; Zerbi, G. 2010. Changes in CO2 emissions after crop conversion from continuous maize to alfalfa. Agriculture, Ecosystems and Environment 136: 139-147.. Thus, for each field, three replicates were used to measure both RS and RH. RH was measured according to Hanson et al. (2000)Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. 2000. Separating root and soil microbial contribution to soil respiration: a review of methods and observations. Biogeochemistry 48: 115-146. and the measurements are considered an indicator of the soil microbial activity.
During each CO2 efflux measurement, the soil respiration chamber was fitted to a PVC collar. The measurements started in Jan 2015 and ended in Dec 2015, with a frequency of two to three times per month, depending on weather variability and management practices, totaling 25 measurements of CO2 per field. Soil respiration was always measured between 08h30 and 12h00 to avoid efflux fluctuations (Xu and Qi, 2001Xu, M.; Qi, Y. 2001. Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology 7: 667-677.; Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.; Fan et al., 2015Fan, L.C.; Yang, M.Z.; Han, W.Y. 2015. Soil respiration under different land uses in eastern China. Plos One 10: e0124198.).
Soil temperature and water content analysis
For each plot, ST and SWC were measured simultaneously with CO2 efflux measurements. The measurements used the built-in temperature probe of environmental gas monitor for ST at a depth of 0.10 m. SWC was determined on soil samples collected from the top 0.2 m layer, and oven dried at 105 °C for constant weight.
Data handling
The one-way analysis of variance (ANOVA) with repeated measures (PROC GLM, SAS) and least significant difference (LSD) tests for pairwise comparisons were used to test differences for RS, RH, ST, and SWC for each of the fields studied. To obtain the best-fit models, the regression analysis was used to examine the relationships between soil respiration (i.e., RS, RH), ST and SWC (Davidson et al., 1998Davidson, E.A.; Belk, E.; Boone, R.D. 1998. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology 4: 217-227.), and a multiple regression model was adopted considering the combined effects of ST and SWC (Fan et al., 2015Fan, L.C.; Yang, M.Z.; Han, W.Y. 2015. Soil respiration under different land uses in eastern China. Plos One 10: e0124198.). The equations used are:
For RS or RH and ST:
For RS or RH and SWC:
For the RS or RH and ST-SWC combinations:
where y is measured RS or RH (μmol CO2 m–2 s–1) and a, b and c are equation coefficients.
The annual cumulative RS and RH were calculated by linear interpolation of the CO2 fluxes between measurement days from Jan 5, 2015, to Dec 29, 2015 (Rong et al., 2015Rong, Y.; Ma, L.; Johnson, D.A.; Yuan, F. 2015. Soil respiration patterns for four major land-use types of the agro-pastoral region of northern China. Agriculture, Ecosystems and Environment 213: 142-150.). The one-way ANOVA was performed to test differences for cumulative RS and RH within the fields of the study sites (i.e., PA vs. PW and HA vs. HW).
Results
ST, SWC, RS, and RH showed different spatial and temporal dynamics, which varied between the study sites, fields, and dates of determination (Figures 4A-H).
Seasonal dynamics of SWC (A, B), ST (C, D), RS (E, F) and RH (G, H) in different fields (PA, PW, HA, HW) of both study sites (plain, hilly) during the study period from Jan to Dec 2015. Vertical dotted lines represent management practices adopted for different crops in the study fields. *p < 0.05; **p < 0.01 (LSD). Vertical bars represent standard errors. PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat; F = fertilizer applied; M = mowing; HR = harvesting; P = plowing; He = herbicide; S = sowing.
Soil temperature and water content dynamics
From Jan to Dec 2015, SWC at 0.2 m of depth ranged from 11 % to 43 % for PA, from 10 % to 40 % for PW, from 10 % to 42 % for HA, and from 14 % to 41 % for HA. Both study sites (i.e., plain and hilly) showed very similar SWC dynamics, with the lowest values observed from July to Oct. Within this period, differences in SWC were less pronounced in the plain study sites (PA vs. PW) compared to the variation for the hilly study sites (HA vs. HW) (Figures 4A and B). The annual mean ST at 0.1 m of depth was 16.32 °C for PA, 15.63 °C for PW, 17.07 °C for HA, and 16.54 °C for HW. In both study sites, the highest ST were recorded during the summer period, which was the same period for the lowest SWC values. Differences for PA versus PW sites occurred mostly during winter and autumn, while for HA versus HW, differences were more scattered throughout the monitoring period (Figures 4C and D).
Soil respiration rates and dynamics
For the plain study site, PA and PW showed annual mean RS of 2.48 μmol CO2 m–2 s–1 and 1.33 μmol CO2 m–2 s–1, respectively. A large reduction in RS was observed for PA in May, followed by a rapid increase from the end of May into June. This reduction in RS occurred concomitantly to the particularly high rainfall volume in May (Figure 3). Subsequently, RS decreased until mid-Sept. Within this observation period, RS for PA showed two peaks to 5.3 μmol CO2 m–2 s–1 that occurred immediately after alfalfa mowing, in the first week of May and the second week of June. RS for PW showed an initial increasing trend until the second week of April, when it decreased to 0.8 μmol CO2 m–2 s–1 in mid-May. Later, RS for PW gradually increased again until July, when the grain was harvested and the plowing was performed about one month later. In the second half of April, (i.e., approximately one month after the second application of fertilizer), RS for PW showed a peak of 2.88 μmol CO2 m–2 s–1. Differences in RS were generally seen between PA and PW from the beginning of Apr to the end of June, when PA showed higher RS than PW did. Starting from mid-Oct, RS was again higher for PA than for PW, which lasted until the end of the monitoring period (Figure 4E).
Conversely, the hilly sites showed similar mean RS between HA and HW (1.33 vs. 1.49 μmol CO2 m–2 s–1, respectively). The peaks in RS were also less pronounced, but more frequent for the hilly sites, where HA showed three RS peaks (3.85, 3.83, 4.04 μmol CO2 m–2 s–1) in the first fortnights of Apr, May and June. Similar to the plain sites, HA showed RS peaks immediately after the alfalfa mowing, except for the first RS peak at the beginning of Apr. In general, HA and HW showed increasing trends for RS from Jan to the first day of Apr. Subsequently, two RS decreases were recorded for HA, in Apr and May, when ST also decreased slightly. RS for HW showed two peaks (3.35, 3.28 μmol CO2 m–2 s–1) for two near measurement dates in Apr, after the second application of fertilizer (Figure 4F). Overall, RS trends were very similar for HA and HW until the first week of Aug, when differences tended to increase, which coincided with both HW plowing and ST decrease (Figure 4D). Indeed, from Jan to mid-June, RS for HW was not different from RS for HA, with the exception of the first half of Apr (Figure 4F).
Heterotrophic soil respiration rates and dynamics
The annual means of RH were 1.19 μmol CO2 m–2 s–1 for PA and 1.03 μmol CO2 m–2 s–1 for PW. RH for PA showed a slowly increasing trend from Jan to the first week of May, when it decreased sharply, concomitant to a sharp SWC decline (Figure 4A). After a peak of RH of 2.65 μmol CO2 m–2 s–1 in the first week of May, RH remained relatively constant from June to the first half of Aug (1.68 to 1.89 μmol CO2 m–2 s–1). A second RH peak for PA was seen in the second half of Aug (2.08 μmol CO2 m–2 s–1), which occurred together with SWC increase (Figures 4A and G). From Sept to Dec, RH for PA remained low and relatively constant (0.11 to 1.07 μmol CO2 m–2 s–1). Similar dynamics for RH for PW were relatively constant through to the beginning of May, when ST was increasing and SWC ranged between 34 % and 42 %. After plowing in the second half of Aug, differences for RH between PA and PW were observed, from Oct to Dec. However, no differences were seen for RH between the two plain fields (PA, PW) for the dates of the two RH peaks for PA.
The annual means RH for the hilly sites were 1.03 μmol CO2 m–2 s–1 for HA and 0.80 μmol CO2 m–2 s–1 for HW. Here, HA showed a slowly increasing RH trend from Jan to Aug, when it then decreased to its minimum of 0.19 μmol CO2 m–2 s–1 at the end of Sept. Very similar RH dynamics were observed for HW, with the exception of a decrease in mid-May. Regarding HA, RH then decreased from Aug to Oct, when SWC was also decreasing. RH for HW increased again until Nov, again similar to SWC. From Jan to the first half of Dec, RH for HA was never higher than RH for HW, except for the second week of May, when HW followed the drop in SWC (Figures 4B and G).
Overall, compared to RS, the variations in RH were less pronounced, with differences concentrated in spring and winter, for both PA vs PW and HA vs HW (Figures 4G and H).
Annual cumulative RS and RH
The annual cumulative RS were 8.97 ± 0.52 t C ha–1 yr–1 for PA and 4.67 ± 0.54 t C ha–1 yr–1 for PW, and 7.43 ± 0.72 t C ha–1 yr–1 for HA and 5.22 ± 0.68 t C ha–1 yr–1 for HW (Figure 5, total bars). The annual cumulative RH were 4.26 ± 0.03 t C ha–1 yr–1 for PA and 3.89 ± 0.20 t C ha–1 yr–1 for PW, and 3.52 ± 0.17 t C ha–1 yr–1 for HA and 2.45 ± 0.18 t C ha–1 yr–1 for HW (Figure 5, shaded bars). For the plain site, PA showed significantly higher annual cumulative RS and RH compared to PW (p < 0.01). However, no differences were seen for annual cumulative RS for the hilly sites (p > 0.05), although RH was significantly higher for HA (p < 0.05).
Cumulative soil respiration in the different fields (PA, PW, HA, HW) of both study sites (plain, hilly) during the study period from Jan to Dec 2015. Bars represent standard errors. Different letters denote significant differences for p < 0.01 (capital letters) or p < 0.05 (small letters). PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat.
Relationships between soil respiration and soil temperature and water content
Over the year, no significant relationships were identified between ST (Eq. (1)) or SWC (Eq. (2)) and RS or RH. Instead, significant linear relationships were defined for each of the fields between the seasonal variations of RS and RH regarding the combination of ST and SWC (Table 2). For the plain sites, the combined model explained 49 % and 48 % of the seasonal variation of RS, and 67 % and 36 % of the seasonal variation of RH, for PA and PW, respectively. For the hilly sites, the model explained 49 % and 43 % of the seasonal variation of RS, and 33 % and 58 % of the seasonal variation of RH, for HA and HW, respectively.
Seasonal variations for RS and RH in combination with ST (depth, 0.1 m) and SWC (depth, 0.2 m) in different fields of both study sites. PA = plain field with alfalfa; PW = plain field with wheat; HA = hilly field with alfalfa; HW = hilly field with wheat; RS = soil respiration; RH = heterotrophic soil respiration.
Discussion
This study measured RS and RH in a Mediterranean environment, for which little information is available compared to other environments (Oertel et al., 2015Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.). The soil CO2 fluxes, mainly RH, is correlated with mineralization of soil organic matter and thus with depletion of soil C stock (Jarvis et al., 2007Jarvis, P.; Rey, A.; Petsikos, C.; Wingate, L.; Rayment, M.; Pereira, J.; Banza, J.; David, J.; Miglietta, F.; Borghetti, M.; Manca, G. 2007. Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the ‘Birch effect.’ Tree Physiology 27: 929-940.; Lai et al., 2017Lai, R.; Arca, A.; Lagomarsino, A.; Cappai, C.; Seddaiu, G.; Demurtas, C.E.; Roggero, P.P. 2017. Manure fertilization increases soil respiration and creates a negative carbon budget in a Mediterranean maize (Zea mays L.)-based cropping system. Catena 151: 202-212.). Peaks in soil CO2 emissions indicate when the C flow to the atmosphere reaches its highest values, and hence when SOM mineralization occurs at the highest rates (Hanson et al., 2000Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. 2000. Separating root and soil microbial contribution to soil respiration: a review of methods and observations. Biogeochemistry 48: 115-146.).
Many previous studies have reported single peaks for soil respiration in areas with different climates. For example, in sub-tropical climates, Fan et al. (2015)Fan, L.C.; Yang, M.Z.; Han, W.Y. 2015. Soil respiration under different land uses in eastern China. Plos One 10: e0124198. reported RS peaks for different cropping systems (including tea gardens with different management intensities, forests, and vegetable fields) concentrated between July and Aug. Fenn et al. (2010)Fenn, K.M.; Malhi, Y.; Morecroft, M.D. 2010. Soil CO2 efflux in a temperate deciduous forest: environmental drivers and component contributions. Soil Biology and Biochemistry 42: 1685-1693. reported a single peak for RS in temperate woodlands from June to July over three monitoring seasons. Tufekcioglu et al. (2001)Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. 2001. Soil respiration within riparian buffers and adjacent crop fields. Plant and Soil 229: 117-124. reported RS peaks for different crop fields (e.g., corn, soybean) and adjacent riparian buffers in a temperate climate concentrated between June and Aug. In all of these cases, the seasonal RS patterns were associated to variations in ST, which reached peaks simultaneously with the soil CO2 effluxes.
In line with other studies carried out in areas under a Mediterranean climate, where the highest ST values correspond to the lowest SWC values in the summer period (e.g., Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.; Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.; Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.), this study shows strong seasonal variability of ST and SWC (Figures 4A-D). Moreover, in contrast to the non-Mediterranean studies mentioned above, this study shows RS dynamics with multiple peaks (Figures 4E and F). These peaks can be explained as a combination of different management practices with variations in ST and SWC.
Soil respiration and management practices
Most studies that have investigated soil C dynamics were performed in ecosystems where human disturbance is relatively rare, such as forest ecosystems (Casals et al., 2000Casals, P.; Romanyà, J.; Cortina, J.; Bottner, P.; Coûteaux, M.M.; Vallejo, V.R. 2000. CO2 efflux from a Mediterranean semi-arid forest soil: seasonality and effects of stoniness. Biogeochemistry 48: 261-281.; Cotrufo et al., 2011Cotrufo, M.F.; Alberti, G.; Inglima, I.; Marjanović, H.; Lecain, D.; Zaldei, A.; Peressotti, A.; Miglietta, F. 2011. Decreased summer drought affects plant productivity and soil carbon dynamics in a Mediterranean woodland. Biogeosciences 8: 2729-2739.; Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.) or other land uses (e.g., Maestre and Cortina, 2003Maestre, F.T.; Cortina, J. 2003. Small-scale spatial variation in soil CO2 efflux in a Mediterranean semiarid steppe. Applied Soil Ecology 23: 199-209.; Munoz-Rojas et al., 2012Munoz-Rojas, M.; Jordan, A.; Zavala, L.M.; De La Rosa, D.; Abd-Elmabod, S.K.; Anaya-Romero, M. 2012. Organic carbon stocks in Mediterranean soil types under different land uses (southern Spain). Solid Earth 3: 375-386.). This study investigated soil C dynamics in ‘nonequilibrium systems’ and thus where management practices had major roles in soil CO2 trends and rates, which affected both RS and RH. In semi-arid grasslands, mowing can suppress RS by ceasing the substrate supply from photosynthesis to the root and rhizosphere microbes (Wan and Luo, 2003Wan, S.; Luo, Y. 2003. Substrate regulation of soil respiration in a tallgrass prairie: results of a clipping and shading experiment. Global Biogeochemical Cycles 17: 1-12.). In addition, mowing can affect ST, as the vegetation removal exposes the soil to more solar radiation and ST increment might positively stimulate the activities of microbes and plant roots (Wei et al., 2016Wei, L.; Liu, J.; Su, J.; Jing, G.; Zhao, J.; Cheng, J.; Jin, J. 2016. Effect of clipping on soil respiration components in temperate grassland of Loess plateau. European Journal of Soil Biology 75: 157-167.). In line with reports by Wan and Luo (2003)Wan, S.; Luo, Y. 2003. Substrate regulation of soil respiration in a tallgrass prairie: results of a clipping and shading experiment. Global Biogeochemical Cycles 17: 1-12., RS decreases were observed after mowing for PA and HA (Figures 4E and F). As there were no significant ST changes immediately after PA and HA mowing (Figures 4C and D), it appears that RS decreases were related only to the effects of the mowing on the root and microbial respiration. The maximum values of RS for PA and HA observed in late spring might have been associated with autotrophic respiration, as the removal of aboveground biomass might increase root metabolic activity (Wei et al., 2016Wei, L.; Liu, J.; Su, J.; Jing, G.; Zhao, J.; Cheng, J.; Jin, J. 2016. Effect of clipping on soil respiration components in temperate grassland of Loess plateau. European Journal of Soil Biology 75: 157-167.).
In cereal-based cropping systems, soil plowing and fertilization might have major roles in soil C dynamics (Oertel et al., 2015Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.). Nitrogen fertilization increase RS rates in wheat (Liu et al., 2016Liu, Q.; Wang, R.; Li, R.; Hu, Y.; Guo, S. 2016. Temperature sensitivity of soil respiration to nitrogen fertilization: varying effects between growing and non-growing seasons. Plos One 11: e0168599.). For HW and PW, no RS peaks were detected immediately after application of N fertilizer. This might be explained by the low soil organic C (Table 1), as an increase in soil N content leads to an increase in RS when there is no limitation for the soil organic C (Micks et al., 2004Micks, P.; Aber, J.D.; Boone, R.D.; Davidson, E.A. 2004. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology and Management 196: 57-70.; Oertel et al., 2015Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.). RS increases observed for the cereal-based systems here (i.e., PW, H W), which occurred roughly one month after application of N fertilizer, might be attributed to an increase in plant biomass production and stimulation of soil biological activity (Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.; Wang et al., 2016Wang, R.; Wang, Z.; Sun, Q.; Zhao, M.; Du, L.; Wu, D.; Li, R.; Gao, X.; Guo, S. 2016. Effects of crop types and nitrogen fertilization on temperature sensitivity of soil respiration in the semi-arid Loess plateau. Soil and Tillage Research 163: 1-9.).
Although soil plowing alters soil porosity and bulk density, with effects on soil CO2 efflux (Carlisle et al., 2006Carlisle, E.A.; Steenwerth, K.L.; Smart, D.R. 2006. Effects of land use on soil respiration. Journal of Environment Quality 35: 1396-1404.), for PW and HW, the soil plowing did not have any immediate effects on RH, probably due to low SWC, which may have inhibited soil microbial activity. RS and RH differences observed between PA and PW and between HA and HW in autumn might be attributed to increase of microbial activities due to soil plowing that altered soil porosity (Oertel et al., 2015Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.).
RS and RH dynamics observed in this study associated to the Mediterranean climate and the effects of the soil management suggest that it is possible to identify agronomic practices that limit soil C losses. For example, it is conceivable to remove the alfalfa aerial biomass produced in summer, which is usually never mowed due to adverse climate conditions that prevent hay from drying in the field. However, biomass removal might be achieved through sheep grazing alfalfa from late summer to autumn, a practice that is still followed in large-scale grazing systems of Mediterranean climate areas (Budimir et al., 2018Budimir, K.; Trombetta, M.F.; Francioni, M.; Toderi, M.; D'Ottavio, P. 2018. Slaughter performance and carcass and meat quality of Bergamasca light lambs according to slaughter age. Small Ruminant Research 164: 1-7.).
Several factors affected CO2 emissions of the cereal-based cropping system. Indeed, reduction in soil plowing depth or no tillage have different effects on soil C dynamics because they in turn affect ST, SWC and soil porosity (Bilandžija et al., 2016Bilandžija, D.; Zgorelec, Z.; Kisić, I. 2016. Influence of tillage practices and crop type on soil CO2 emissions. Sustainability 8: 1-10.; López-Garrido et al., 2014López-Garrido, R.; Madejó, N.E.; Moreno, F.; Murillo, J.M. 2014. Conservation tillage influence on carbon dynamics under Mediterranean conditions. Pedosphere 24: 65-75.). The Mediterranean climate conditions are characterized by low SWC during the summer. Thus, as well as shallower soil plowing depth (López-Garrido et al., 2014López-Garrido, R.; Madejó, N.E.; Moreno, F.; Murillo, J.M. 2014. Conservation tillage influence on carbon dynamics under Mediterranean conditions. Pedosphere 24: 65-75.), it is conceivable that soil tillage performed in early summer might help to reduce soil CO2 emissions, because of the limiting effect of SWC. Further soil tillage should then be performed as soon as possible to reduce soil porosity before SWC increase, which usually occurs in autumn in the Mediterranean climate areas.
Finally, to maximize the impact of specific management practices to reduce soil GHG emissions, practices should be included into agro-environmental measures at the landscape scale to involve more farmers (e.g., Toderi et al., 2017Toderi, M.; Francioni, M.; Seddaiu, G.; Roggero, P.P.; Trozzo, L.; D'Ottavio, P. 2017. Bottom-up design process of agri-environmental measures at a landscape scale: Evidence from case studies on biodiversity conservation and water protection. Land Use Policy 68: 295-305.).
Annual cumulative CO2 efflux
In general, the annual cumulative RS emissions in this study are within the ranges reported by Raich and Schlesinger (1992)Raich, J.; Schlesinger, W.W.H. 1992. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 44: 81-99. for cropland and grassland ecosystems. The cumulative RS emission of the forage-based fields here (PA, HA) were much higher than those reported by Paustian et al. (1990)Paustian, K.; Andrén, O.; Clarholm, M.; Hansson, A.C.; Johansson, G.; Lagerlof, J.; Lindberg, T.; Pettersson, R.; Sohlenius, B. 1990. Carbon and nitrogen budgets of four ecosystems with annual and perennial crops, with and without N fertilization. Journal of Applied Ecology 27: 60-84. and by Gong et al. (2015)Gong, J.R.R.; Xu, S.; Wang, Y.; Luo, Q.; Liu, M.; Zhang, W. 2015. Effect of irrigation on the soil respiration of constructed grasslands in inner Mongolia, China. Plant and Soil 395: 159-172. under other climate conditions (i.e., Sweden, inner Mongolia, respectively), where CO2 rates are expected to be lower due to lower SOM mineralization rates. The annual cumulative RS emissions of the cereal-based fields here (i.e., PW, HW) are in line with those reported by Lai et al. (2017)Lai, R.; Arca, A.; Lagomarsino, A.; Cappai, C.; Seddaiu, G.; Demurtas, C.E.; Roggero, P.P. 2017. Manure fertilization increases soil respiration and creates a negative carbon budget in a Mediterranean maize (Zea mays L.)-based cropping system. Catena 151: 202-212. under similar climate conditions.
The annual cumulative RS for the plain study site and RH rates for both study sites were higher for alfalfa. Higher RS and/or RH rates are probably related to soil SOM (Table 1) and its light fraction in these fields. Previous studies have shown that the SOM light fraction increases under crop rotation, including for alfalfa, and with the number of alfalfa growing years (Wang et al., 2009Wang, X.L.; Jia, Y.; Li, X.G.; Long, R.J.; Ma, Q.; Li, F.M.; Song, Y.J. 2009. Effects of land use on soil total and light fraction organic, and microbial biomass C and N in a semi-arid ecosystem of northwest China. Geoderma 153: 285-290.; Zhang et al., 2009Zhang, T.; Wang, Y.; Wang, X.; Wang, Q.; Han, J. 2009. Organic carbon and nitrogen stocks in reed meadow soils converted to alfalfa field. Soil and Tillage Research 105: 143-148.), while cereal-based cropping systems have detrimental effects on all organic C pools (Bongiovanni and Lobartini, 2006Bongiovanni, M.D.; Lobartini, J.C. 2006. Particulate organic matter, carbohydrate, humic acid contents in soil macro- and microaggregates as affected by cultivation. Geoderma 136: 660-665.). Higher SOM stimulates microbial activity (García-Orenes et al., 2010García-Orenes, F.; Guerrero, C.; Roldán, A.; Mataix-Solera, J.; Cerdà, A.; Campoy, M.; Zornoza, R.; Bárcenas, G.; Caravaca, F. 2010. Soil microbial biomass and activity under different agricultural management systems in a semiarid Mediterranean agroecosystem. Soil and Tillage Research 109: 110-115.; Lai et al., 2014Lai, R.; Lagomarsino, A.; Ledda, L.; Roggero, P.P. 2014. Variation in soil C and microbial functions across tree canopy projection and open grassland microenvironments. Turkish Journal of Agriculture and Forestry 38: 62-69.); therefore, alfalfa fields are expected to have higher CO2 emission rates compared to tilled cropping systems (Frank et al., 2006Frank, A.B.; Liebig, M.A.; Tanaka, D.L. 2006. Management effects on soil CO2 efflux in northern semiarid grassland and cropland. Soil and Tillage Research 89: 78-85.). In general, the continuous presence of legumes in the forage-based cropping system (Figure 2) favored a continuous supply of organic C, concurrent with a high level of available N, which leads microorganisms to degrade the SOM light fraction thus enhancing RH (Leitner et al., 2012Leitner, S.; Wanek, W.; Wild, B.; Haemmerle, I.; Kohl, L.; Keiblinger, K.M.; Zechmeister-Boltenstern, S.; Richter, A. 2012. Influence of litter chemistry and stoichiometry on glucan depolymerization during decomposition of beech (Fagus sylvatica L.) litter. Soil Biology and Biochemistry 50: 174-187.).
Conversely, the absence of differences in the annual cumulative RS in the hilly sites here (i.e., HA vs. HW) might be attributed to the herbicide treatment in the second half of Oct for HA suppression before the sod seeding of wheat in early Nov, which might have affected both the plant root and soil microbial activities (Nguyen et al., 2016Nguyen, D.B.; Rose, M.T.; Rose, T.J.; Morris, S.G.; van Zwieten, L. 2016. Impact of glyphosate on soil microbial biomass and respiration: a meta-analysis. Soil Biology and Biochemistry 92: 50-57.).
The soil C improvements for the forage-based cropping system, along with higher CO2 emissions, are a considerable trade-off between the different ecosystem services (D'Ottavio et al., 2018D'Ottavio, P.; Francioni, M.; Trozzo, L.; Sedić, E.; Budimir, K.; Avanzolini, P.; Trombetta, M.F.; Porqueddu, C.; Santilocchi, R.; Toderi, M. 2018. Trends and approaches in the analysis of ecosystem services provided by grazing systems: a review. Grass and Forage Science 73: 15-25.), which require further investigations.
Effects of soil temperature and water content on soil respiration
Unlike some studies carried out under similar climate conditions (Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.; Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.), no relationships were found here between soil CO2 efflux and ST or SWC during the year using either exponential or linear models. While some studies have reported very high correlation coefficients between soil respiration and ST through the definition of empirical SWC thresholds (e.g., Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.; Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.; Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.), all attempts to use empirical threshold values for SWC did not result in better correlation coefficients for RS or RH for any of these study fields (data not shown). If, on the one hand, there is no clear relationship between soil respiration drivers and CO2 emissions during the entire year; on the other hand, there emerges a strong relationship between ST and CO2 only for the first part of the year (i.e., from Jan to June for PA-HA, from Jan to May for PW-HW), as reported by other studies under Mediterranean climate conditions (e.g., Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.). In this case, ST was the main driver of soil CO2 emissions, as a simple exponential model (Eq. (1)) explained 82 % (p < 0.01), 82 % (p < 0.05), 80 % (p < 0.01), and 68 % (p < 0.05) of the seasonal variations of RS in PA, PW, HA, and HW, respectively. During the same period, the exponential model explained 78 % (p < 0.01) and 84 % (p < 0.01) of the seasonal variation of RH in PA ad HA, respectively, while no significant relationships were observed for PW (R2 = 0.38; p > 0.05) and HW (R2 = 0.55; p > 0.05). For the rest of the year, RS and/or RH did not only depend on ST and SWC, as other factors had major roles, which may have been either biotic factors, such as microbial or enzyme activities (Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.; Fan et al., 2015Fan, L.C.; Yang, M.Z.; Han, W.Y. 2015. Soil respiration under different land uses in eastern China. Plos One 10: e0124198.; Lai et al., 2014Lai, R.; Lagomarsino, A.; Ledda, L.; Roggero, P.P. 2014. Variation in soil C and microbial functions across tree canopy projection and open grassland microenvironments. Turkish Journal of Agriculture and Forestry 38: 62-69.; Wang et al., 2016Wang, R.; Wang, Z.; Sun, Q.; Zhao, M.; Du, L.; Wu, D.; Li, R.; Gao, X.; Guo, S. 2016. Effects of crop types and nitrogen fertilization on temperature sensitivity of soil respiration in the semi-arid Loess plateau. Soil and Tillage Research 163: 1-9.), and abiotic factors, such as soil disturbance (e.g., soil tillage) or changes to vegetation (e.g., mowing) (Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.; Liang et al., 2015Liang, G.; Houssou, A.A.; Wu, H.; Cai, D.; Wu, X.; Gao, L.; Li, J.; Wang, B.; Li, S. 2015. Seasonal patterns of soil respiration and related soil biochemical properties under nitrogen addition in winter wheat field. Plos One 10: e0144115.; Ryan and Law, 2005Ryan, M.G.; Law, B.E. 2005. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 73: 3-27.), as previously discussed. This implies that modeling soil respiration under these Mediterranean climate conditions tends to be difficult, especially if only ST and SWC are the factors considered (Cotrufo et al., 2011Cotrufo, M.F.; Alberti, G.; Inglima, I.; Marjanović, H.; Lecain, D.; Zaldei, A.; Peressotti, A.; Miglietta, F. 2011. Decreased summer drought affects plant productivity and soil carbon dynamics in a Mediterranean woodland. Biogeosciences 8: 2729-2739.; Lai et al., 2012Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.; Oyonarte et al., 2012Oyonarte, C.; Rey, A.; Raimundo, J.; Miralles, I.; Escribano, P. 2012. The use of soil respiration as an ecological indicator in arid ecosystems of the SE of Spain: spatial variability and controlling factors. Ecological Indicators 14: 40-49.), as during the summer, low SWC has a major role in inhibiting both RS and RH (Davidson et al., 1998Davidson, E.A.; Belk, E.; Boone, R.D. 1998. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology 4: 217-227.; Xu and Qi, 2001Xu, M.; Qi, Y. 2001. Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology 7: 667-677.). In addition, ST and SWC often vary together and this co-variation prevents the emergence of clear relationships with RS and/or RH (Davidson et al., 1998Davidson, E.A.; Belk, E.; Boone, R.D. 1998. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology 4: 217-227.; Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.). Nevertheless, in this study, the two-variable model that combined ST and SWC did explain 43 % to 49 % and 33 % to 67 % of the annual variations for RS and RH, respectively. These values appear to be slightly below those reported in other studies carried out under similar climate conditions (Rey et al., 2002Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.; Almagro et al., 2009Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.). For example, for PA, the ST-SWC combined model was the best RH predictor due to the absence of soil disturbance (R2 = 0.67; p < 0.01). Conversely, for the same study site, the same model explained only 36 % of the annual variation of PW RH. This was mainly due to management practices usually carried out in these cropping systems (i.e., fertilizer, soil plowing, mowing, herbicides), which affect RS and RH (Carlisle et al., 2006Carlisle, E.A.; Steenwerth, K.L.; Smart, D.R. 2006. Effects of land use on soil respiration. Journal of Environment Quality 35: 1396-1404.; Wan and Luo, 2003Wan, S.; Luo, Y. 2003. Substrate regulation of soil respiration in a tallgrass prairie: results of a clipping and shading experiment. Global Biogeochemical Cycles 17: 1-12.; Wei et al., 2016Wei, L.; Liu, J.; Su, J.; Jing, G.; Zhao, J.; Cheng, J.; Jin, J. 2016. Effect of clipping on soil respiration components in temperate grassland of Loess plateau. European Journal of Soil Biology 75: 157-167.), and consequently introduced bias into the model.
Conclusions
In this study, RS and RH showed different dynamics, with spatial and temporal variability across these study sites. The forage-based cropping system (alfalfa) showed higher annual cumulative RS only for the plain area site (PA). Conversely, and probably due to herbicide effects on alfalfa RS, there were no differences for the hilly site (HA). The forage-based cropping systems (PA, HA) showed higher annual cumulative RH, as well as higher SOM and total organic C accumulation. The model with ST and SWC combination provided better prediction for RS and RH variation for all study sites, when compared to single ST or SWC modeling over the year, or to models that use empirical threshold values.
Although these findings contribute towards filling some gaps in knowledge of cropping systems under Mediterranean climate conditions, further studies should focus on drives of RS and RH. Thus, further studies on soil C dynamics should increase the effects of common management practices, particularly regarding areas under Mediterranean climate conditions, where the data are relatively fragmented and controversial and where they have mainly addressed systems with limited human disturbance.
References
- Alberti, G.; Delle Vedove, G.; Zuliani, M.; Peressotti, A.; Castaldi, S.; Zerbi, G. 2010. Changes in CO2 emissions after crop conversion from continuous maize to alfalfa. Agriculture, Ecosystems and Environment 136: 139-147.
- Almagro, M.; López, J.; Querejeta, I.; Martínez-Mena, M. 2009. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biology and Biochemistry 41: 594-605.
- Bilandžija, D.; Zgorelec, Z.; Kisić, I. 2016. Influence of tillage practices and crop type on soil CO2 emissions. Sustainability 8: 1-10.
- Bongiovanni, M.D.; Lobartini, J.C. 2006. Particulate organic matter, carbohydrate, humic acid contents in soil macro- and microaggregates as affected by cultivation. Geoderma 136: 660-665.
- Budimir, K.; Trombetta, M.F.; Francioni, M.; Toderi, M.; D'Ottavio, P. 2018. Slaughter performance and carcass and meat quality of Bergamasca light lambs according to slaughter age. Small Ruminant Research 164: 1-7.
- Caballero, R.; Fernández-González, F.; Pérez-Badia, R.; Molle, G.; Roggero, P.P.; Bagella, S.; D'Ottavio, P.; Papanastasis, V.P.; Fotiadis, G.; Sidiropoulou, A.; Ispikoudis, I. 2009. Grazing systems and biodiversity in Mediterranean areas: Spain, Italy and Greece. Pastos 39: 9-152.
- Carlisle, E.A.; Steenwerth, K.L.; Smart, D.R. 2006. Effects of land use on soil respiration. Journal of Environment Quality 35: 1396-1404.
- Casals, P.; Romanyà, J.; Cortina, J.; Bottner, P.; Coûteaux, M.M.; Vallejo, V.R. 2000. CO2 efflux from a Mediterranean semi-arid forest soil: seasonality and effects of stoniness. Biogeochemistry 48: 261-281.
- Cotrufo, M.F.; Alberti, G.; Inglima, I.; Marjanović, H.; Lecain, D.; Zaldei, A.; Peressotti, A.; Miglietta, F. 2011. Decreased summer drought affects plant productivity and soil carbon dynamics in a Mediterranean woodland. Biogeosciences 8: 2729-2739.
- Davidson, E.A.; Belk, E.; Boone, R.D. 1998. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology 4: 217-227.
- Di Bene, C.; Maretti, A.; Francaviglia, R.; Farina, R. 2016. Soil organic carbon dynamics in typical durum wheat-based crop rotations of southern Italy. Italian Journal of Agronomy 11: 209-216.
- D'Ottavio, P.; Francioni, M.; Trozzo, L.; Sedić, E.; Budimir, K.; Avanzolini, P.; Trombetta, M.F.; Porqueddu, C.; Santilocchi, R.; Toderi, M. 2018. Trends and approaches in the analysis of ecosystem services provided by grazing systems: a review. Grass and Forage Science 73: 15-25.
- Fan, L.C.; Yang, M.Z.; Han, W.Y. 2015. Soil respiration under different land uses in eastern China. Plos One 10: e0124198.
- Feiziene, D.; Janusauskaite, D.; Feiza, V.; Putramentaite, A.; Sinkeviciene, A.; Suproniene, A.; Seibutis, V.; Kadziene, G.; Deveikyte, I.; Lazauskas, S.; Janusauskaite, D. 2015. After-effect of long-term soil management on soil respiration and other qualitative parameters under prolonged dry soil conditions. Turkish Journal of Agriculture and Forestry 39: 633-651.
- Fenn, K.M.; Malhi, Y.; Morecroft, M.D. 2010. Soil CO2 efflux in a temperate deciduous forest: environmental drivers and component contributions. Soil Biology and Biochemistry 42: 1685-1693.
- Frank, A.B.; Liebig, M.A.; Tanaka, D.L. 2006. Management effects on soil CO2 efflux in northern semiarid grassland and cropland. Soil and Tillage Research 89: 78-85.
- García-Orenes, F.; Guerrero, C.; Roldán, A.; Mataix-Solera, J.; Cerdà, A.; Campoy, M.; Zornoza, R.; Bárcenas, G.; Caravaca, F. 2010. Soil microbial biomass and activity under different agricultural management systems in a semiarid Mediterranean agroecosystem. Soil and Tillage Research 109: 110-115.
- Giannakopoulos, C.; Le Sager, P.; Bindi, M.; Moriondo, M.; Kostopoulou, E.; Goodess, C.M. 2009. Climatic changes and associated impacts in the Mediterranean resulting from 2 °C global warming. Global and Planetary Change 68: 209-224.
- Gong, J.R.R.; Xu, S.; Wang, Y.; Luo, Q.; Liu, M.; Zhang, W. 2015. Effect of irrigation on the soil respiration of constructed grasslands in inner Mongolia, China. Plant and Soil 395: 159-172.
- Guo, L.B.; Gifford, R.M. 2002. Soil carbon stocks and land use change: a meta-analysis. Global Change Biology 8: 345-360.
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. 2000. Separating root and soil microbial contribution to soil respiration: a review of methods and observations. Biogeochemistry 48: 115-146.
- Jarvis, P.; Rey, A.; Petsikos, C.; Wingate, L.; Rayment, M.; Pereira, J.; Banza, J.; David, J.; Miglietta, F.; Borghetti, M.; Manca, G. 2007. Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the ‘Birch effect.’ Tree Physiology 27: 929-940.
- Lai, R.; Arca, A.; Lagomarsino, A.; Cappai, C.; Seddaiu, G.; Demurtas, C.E.; Roggero, P.P. 2017. Manure fertilization increases soil respiration and creates a negative carbon budget in a Mediterranean maize (Zea mays L.)-based cropping system. Catena 151: 202-212.
- Lai, R.; Lagomarsino, A.; Ledda, L.; Roggero, P.P. 2014. Variation in soil C and microbial functions across tree canopy projection and open grassland microenvironments. Turkish Journal of Agriculture and Forestry 38: 62-69.
- Lai, R.; Seddaiu, G.; Gennaro, L.; Roggero, P.P. 2012. Effects of nitrogen fertilizer sources and temperature on soil CO2 efflux in Italian ryegrass crop under Mediterranean conditions. Italian Journal of Agronomy 7: 196-201.
- Lal, R.D.A. 2004. Soil carbon sequestration impacts on global climate change and food security. Science 304: 1623-1627.
- Leitner, S.; Wanek, W.; Wild, B.; Haemmerle, I.; Kohl, L.; Keiblinger, K.M.; Zechmeister-Boltenstern, S.; Richter, A. 2012. Influence of litter chemistry and stoichiometry on glucan depolymerization during decomposition of beech (Fagus sylvatica L.) litter. Soil Biology and Biochemistry 50: 174-187.
- Liang, G.; Houssou, A.A.; Wu, H.; Cai, D.; Wu, X.; Gao, L.; Li, J.; Wang, B.; Li, S. 2015. Seasonal patterns of soil respiration and related soil biochemical properties under nitrogen addition in winter wheat field. Plos One 10: e0144115.
- Liu, Q.; Wang, R.; Li, R.; Hu, Y.; Guo, S. 2016. Temperature sensitivity of soil respiration to nitrogen fertilization: varying effects between growing and non-growing seasons. Plos One 11: e0168599.
- López-Garrido, R.; Madejó, N.E.; Moreno, F.; Murillo, J.M. 2014. Conservation tillage influence on carbon dynamics under Mediterranean conditions. Pedosphere 24: 65-75.
- Maestre, F.T.; Cortina, J. 2003. Small-scale spatial variation in soil CO2 efflux in a Mediterranean semiarid steppe. Applied Soil Ecology 23: 199-209.
- Micks, P.; Aber, J.D.; Boone, R.D.; Davidson, E.A. 2004. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology and Management 196: 57-70.
- Monaci, E.; Polverigiani, S.; Neri, D.; Bianchelli, M.; Santilocchi, R.; Toderi, M.; D'Ottavio, P.; Vischetti, C. 2017. Effect of contrasting crop rotation systems on soil chemical and biochemical properties and plant root growth in organic farming: first results. Italian Journal of Agronomy 12: 364-374.
- Munoz-Rojas, M.; Jordan, A.; Zavala, L.M.; De La Rosa, D.; Abd-Elmabod, S.K.; Anaya-Romero, M. 2012. Organic carbon stocks in Mediterranean soil types under different land uses (southern Spain). Solid Earth 3: 375-386.
- Nguyen, D.B.; Rose, M.T.; Rose, T.J.; Morris, S.G.; van Zwieten, L. 2016. Impact of glyphosate on soil microbial biomass and respiration: a meta-analysis. Soil Biology and Biochemistry 92: 50-57.
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. 2015. Greenhouse gas emissions from soils: a review. Geochemistry 76: 327-352.
- Oyonarte, C.; Rey, A.; Raimundo, J.; Miralles, I.; Escribano, P. 2012. The use of soil respiration as an ecological indicator in arid ecosystems of the SE of Spain: spatial variability and controlling factors. Ecological Indicators 14: 40-49.
- Paustian, K.; Andrén, O.; Clarholm, M.; Hansson, A.C.; Johansson, G.; Lagerlof, J.; Lindberg, T.; Pettersson, R.; Sohlenius, B. 1990. Carbon and nitrogen budgets of four ecosystems with annual and perennial crops, with and without N fertilization. Journal of Applied Ecology 27: 60-84.
- Paustian, K.; Six, J.; Elliott, E.T.; Hunt, H.W. 2000. Management options for reducing CO2 emissions from agricultural soils. Biogeochemistry 48: 147-163.
- Raich, J.; Schlesinger, W.W.H. 1992. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 44: 81-99.
- Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G; Valentini, R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology 8: 851-866.
- Robertson, F.; Armstrong, R.; Partington, D.; Perris, R.; Oliver, I.; Aumann, C.; Crawford, D.; Rees, D. 2015. Effect of cropping practices on soil organic carbon: evidence from long-term field experiments in Victoria, Australia. Soil Research 53: 636-646.
- Rong, Y.; Ma, L.; Johnson, D.A.; Yuan, F. 2015. Soil respiration patterns for four major land-use types of the agro-pastoral region of northern China. Agriculture, Ecosystems and Environment 213: 142-150.
- Ryan, J.; Masri, S.; Ibrikçi, H.; Singh, M.; Pala, M.; Harris, H.C. 2008. Implications of cereal-based crop rotations, nitrogen fertilization, and stubble grazing on soil organic matter in a Mediterranean-type environment. Turkish Journal of Agriculture and Forestry 32: 289-297.
- Ryan, M.G.; Law, B.E. 2005. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 73: 3-27.
- Smith, P. 2008. Land use change and soil organic carbon dynamics. Nutrient Cycling in Agroecosystems 81: 169-178.
- Soil Survey Staff. 2014. Keys to Soil Taxonomy. 12ed. USDA-Natural Resources Conservation Service, Washington, DC, USA.
- Toderi, M.; Francioni, M.; Seddaiu, G.; Roggero, P.P.; Trozzo, L.; D'Ottavio, P. 2017. Bottom-up design process of agri-environmental measures at a landscape scale: Evidence from case studies on biodiversity conservation and water protection. Land Use Policy 68: 295-305.
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. 2001. Soil respiration within riparian buffers and adjacent crop fields. Plant and Soil 229: 117-124.
- Wan, S.; Luo, Y. 2003. Substrate regulation of soil respiration in a tallgrass prairie: results of a clipping and shading experiment. Global Biogeochemical Cycles 17: 1-12.
- Wang, R.; Wang, Z.; Sun, Q.; Zhao, M.; Du, L.; Wu, D.; Li, R.; Gao, X.; Guo, S. 2016. Effects of crop types and nitrogen fertilization on temperature sensitivity of soil respiration in the semi-arid Loess plateau. Soil and Tillage Research 163: 1-9.
- Wang, X.L.; Jia, Y.; Li, X.G.; Long, R.J.; Ma, Q.; Li, F.M.; Song, Y.J. 2009. Effects of land use on soil total and light fraction organic, and microbial biomass C and N in a semi-arid ecosystem of northwest China. Geoderma 153: 285-290.
- Wei, L.; Liu, J.; Su, J.; Jing, G.; Zhao, J.; Cheng, J.; Jin, J. 2016. Effect of clipping on soil respiration components in temperate grassland of Loess plateau. European Journal of Soil Biology 75: 157-167.
- Xu, M.; Qi, Y. 2001. Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology 7: 667-677.
- Zhang, T.; Wang, Y.; Wang, X.; Wang, Q.; Han, J. 2009. Organic carbon and nitrogen stocks in reed meadow soils converted to alfalfa field. Soil and Tillage Research 105: 143-148.
Edited by
Publication Dates
-
Publication in this collection
05 Sept 2019 -
Date of issue
2020
History
-
Received
04 Apr 2018 -
Accepted
06 Nov 2018