Óxido Nítrico; Síntases; Metabolismo; Óxido Nítrico mensageiro
Nitric Oxide; Nitric Oxide Synthases; Nitric Oxide messenger; Long Term Potentiation; Nitric Oxide Synthesis and Metabolism
Artigo de Revisão
Óxido nítrico: o simples mensageiro percorrendo a complexidade. Metabolismo, síntese e funções.
R. Flora Filho, B. Zilberstein
Departamento de Gastroenterologia da Faculdade de Medicina da Universidade de São Paulo, São Paulo, SP.
UNITERMOS: Óxido Nítrico. Síntases. Metabolismo. Óxido Nítrico mensageiro.
KEY WORDS: Nitric Oxide. Nitric Oxide Synthases. Nitric Oxide messenger. Long Term Potentiation. Nitric Oxide Synthesis and Metabolism.
INTRODUÇÃO
Vamos supor que após ler este artigo, você guarde na sua memória tardia uma parte do que leu. Provavelmente você estará utilizando para isto uma das menores e mais versáteis moléculas do seu organismo: o óxido nítrico (NO - Nitric Oxide). Esta pequena e simples molécula, talvez a menor produzida pelos mamíferos, tem efeitos fascinantes desde a manutenção inicial da vida, através do controle da circulação placentária, ou a indução do início da vida através da regulagem das contrações uterinas no trabalho de parto, como também efeitos letais demonstráveis, por exemplo, no choque séptico. O óxido nítrico é um importante neurotransmissor com capacidade potencializadora, atuando na memória e no aprendizado, podendo também ter ações endócrinas, autócrinas e parácrinas. A sua ação na imuno-regulação está presente na inflamação e nos mecanismos de autoimunidade. Esta molécula tem revolucionado e obrigado revisão de paradigmas da medicina, principalmente em neurologia, cardiologia, nefrologia e gastroenterologia.
Este artigo procura mostrar os mecanismos de síntese e formação do óxido nítrico apresentando a enzima envolvida - síntase do óxido nítrico (nitric oxide synthase - NOS) e os efeitos do NO como mensageiro nos principais sistemas do organismo.
O início
A evidência inicial de óxidos de nitrogênio no metabolismo vieram de experimentos que demonstraram produção de nitratos em camundongos germ-free no início da década de 801. Em 1985 demonstrou-se que macrófagos ativados por lipopossacárides bacterianos eram capazes de levar à produção de nitritos e nitratos2. Na seqüência evidenciou-se que a L-arginina era o substrato e a L-citrulina era formada como co-produto3. Em 1988, Marletta identificou o óxido nítrico como o produto da reação de oxiredução da L-arginina4.
Quase que simultaneamente, Furchgott investigava um fator vasodilatador associado ao endotélio vascular (endothelium-derived relaxing factor - EDRF5 e poucos anos mais tarde concluiu-se ser o NO responsável pela atividade biológica do EDRF6.
Durante o final da década de 80 e início de 90, a comunidade científica aprofundou pesquisas nesta direção, adicionando importantes conhecimentos sobre o NO como mensageiro (ou sinalizador inter e intracelular) e como toxina, atuando em inúmeros processos patológicos.
Características, síntese e metabolismo do NO
O óxido nítrico é uma molécula gasosa simples, habitualmente encontrada no ar atmosférico em pequenas quantidades, altamente tóxica devido à presença de radical livre (elétron extra) que a torna um agente químico altamente reativo. Quando diluído, o NO tem uma meia vida de menos de 10 segundos devido à sua rápida oxidação a nitrito e nitrato. O NO liga-se à hemoglobina e outras proteínas que contém o núcleo heme levando ao término de sua atividade biológica7.
A fig. 1 indica a clássica reação química de formação do NO, em que a L-arginina é transformada em um intermediário, a NG-hidroxi-L-arginina com a presença de nicotinamida-adenina-dinucleotídeo-fostato-hidrogênio (NADPH) e Ca2+ sendo necessário mais NADPH e O2 para a formação de L-citrulina e NO.
A L-arginina é um aminoácido semi-essencial produzido no organismo, porém em quantidade insuficiente para todas necessidades. Além do ciclo da uréia, a arginina é utilizada na síntese de creatinina e fornece ornitina para a síntese de poliaminas9. Como há uma solicitação metabólica continuada da L-arginina, existe uma neo-síntese da L-arginina nos túbulos proximais renais a partir da citrulina. Proteínas ingeridas são degradadas até arginina, que podem ser diretamente absorvidas e utilizadas no ciclo da uréia no tecido hepático, ou transformadas no epitélio intestinal em ornitina que, juntamente com a glutamina secretada como glutamato, são convertidas em citrulina. A citrulina absorvida se transforma em arginina no ciclo renal (fig. 2)9. A citrulina também pode ser convertida diretamente em L-arginina no citoplasma das células endoteliais e dos macrófagos.
A síntese enzimática de citrulina (Fig. 1), pode ser inibida por análogos da L-arginina tais como NG-monometil-L-arginina (L-NMMA), NG-nitro-L-arginina (L-NNA) e NG-nitro-L-arginina-metil-éster (L-NAME)7,8. Estes inibidores têm grande importância na pesquisa dos prováveis efeitos do NO nos tecidos, uma vez que a substituição do substrato habitual (L-arginina) pelos análogos irá inibir a produção de NO e seus efeitos conseqüentes. Vale salientar que a D-arginina não substitui a L-arginina nesta reação para formação do NO.
A demonstração da produção de NO é ainda difícil, sendo sempre feita de maneira indireta. Aliás, todas pesquisas pioneiras não demonstraram o NO propriamente dito devido sua evanescência, considerando-se a concentração de nitrito e nitrato como prova de sua produção. Outro método de demonstrar o NO, como já citado, é a substituição do substrato por um análogo (L-NNA ou L-NMMA ou L-NAME) sendo a ausência do efeito pesquisado imputado pela não formação de NO devido ao bloqueio da reação da L-arginina à L-citrulina.
As isoenzimas da sintase do NO
Muitas células são capazes de sintetizar o NO através de hemeproteínas da família citocromo P450-like, chamadas de NO síntases (NOS). As NOS são dependentes de O2, NADPH, flavinas e biopterinas para exercer sua atividade. Até o momento, foram isoladas e clonadas três isoenzimas, sendo duas constitutivas em determinadas células e uma induzível, recebendo as siglas respectivas de cNOS e iNOS. Todas as três isoenzimas são semelhantes estruturalmente, porém reguladas de modo diverso e induzidas a partir de genes localizados nos cromossomos 7 (isoforma I), 12 (isoforma II) e 17 (isoforma III)31.
A cNOS ou Isoforma I está presente no cérebro e foi purificada inicialmente no cerebelo do camundongo e do porco10. Na literatura esta enzima pode ser chamada também de bNOS (brain NO synthase) ou nNOS (neuronal NO synthase). Esta proteína, atualmente clonada de cérebros humanos, mantém uma seqüência de aminoácidos altamente conservada entre as espécies, ocorrendo 93% de identidade entre a cNOS de humanos e camundongos9.
A iNOS ou Isoforma II não é expressa constitutivamente, ou seja, não está presente de modo habitual, sendo induzida nos macrófagos e outras células por lipopolisacárides bacterianos e/ou citoquinas11. Esta isoenzima também pode ser chamada de macNOS (macrophage NO synthase). Vários grupos clonaram a iNOS também em músculo liso45, hepatócitos de camundongo11 e em hepatócitos humanos12. Vários autores consideram que qualquer célula do organismo tem a capacidade de produzir iNOS sob estímulos apropriados. Uma vez induzida, a iNOS é capaz de produzir NO por longo tempo, e isso vem a caracterizar seu envolvimento em vários processos patológicos. Assim, o alto nível de NO produzido por macrófagos ou por neutrófilos ou outras células ativadas, que deveria ser tóxico para micróbios, parasitas ou células tumorais, pode também lesar células saudáveis vizinhas, sendo este mecanismo responsável pela maioria de processos inflamatórios e autoimunes.
A eNOS ou Isoforma III é expressa constitutivamente nas células endoteliais podendo também ser chamada cNOS ou EC-NOS (endothelial constitutive NO synthase). O código genético responsável pela isoforma III foi clonado a partir de células endoteliais de bovinos e humanos13, confirmando o local preferencial de produção e ação.
As isoenzimas NOS podem ser, sob ponto de vista prático, caracterizadas como de baixo ou alto débito conforme a duração da atividade da NOS. As isoformas I e III (cNOS e eNOS) são de baixo débito, estando envolvidas em processos homeostáticos como neurotransmissão, peristaltismo, controle imediato da pressão arterial14. Considera-se a eNOS de menor débito do que cNOS. Já a isoenzima II ou iNOS, quando estimulada permanece em atividade por horas com mecanismo de sinergismo de indução inclusive do próprio NO produzido12. Esta ação lhe confere uma importante característica podendo levar à morte da célula15.
Outra classificação das NOS é determinada pela sua dependência de cálcio para ativação. O Ca2+ é um importante sinalizador citoplasmático, atuando a partir de ligações com proteínas intracelulares receptoras específicas. Uma destas proteínas citoplasmáticas (Ca2+-receptoras) é a calmodulina, encontrada em praticamente todas células. Quando o Ca2+ se liga à calmodulina forma-se o complexo Ca2+/calmodulina, este, um elemento regulatório de algumas atividades enzimáticas intracelulares. Quando a concentração intracelular de Ca2+ cai, desfaz-se o complexo Ca2+/calmodulina e a atividade enzimática é desativada. Este mecanismo Ca2+/calmodulina é responsável pela ativação da NOS.
No caso da cNOS, segue-se o mecanismo descrito, ou seja, é necessária determinada concentração de Ca2+ intracelular para atividade enzimática, ocorrendo inativação da cNOS com queda do Ca2+ citoplasmático abaixo de determinado nível. Para a iNOS, o mecanismo de ação depende da concentração intracelular de Ca2+ somente para ativação, sendo que a queda do Ca2+ intracelular não inibe a atividade da iNOS (Fig. 3). Assim, classificam-se as NOS como cálcio-dependentes (que são as isoformas I e III) e cálcio-independentes (isoforma II)20.
A Tabela 1 sumariza as abreviaturas, sinonímia e diferenças quanto ao débito e dependência de Ca2+ das síntases do NO.
Estes conhecimentos quanto à classificação e modo de ação das três NOS são importantes para entender os demais mecanismos envolvidos na ação do NO.
Aspectos fisiológicos e tóxicos do NO
As funções do NO até hoje descobertas são complexas e antagônicas. Um aspecto marcante desta molécula é a sua capacidade de ser benéfica ou potencialmente tóxica conforme a concentração ou depuração tecidual. Alguns autores, como Schmidt16, denominam muito apropriadamente o NO como uma "faca de dois gumes" (double-edged sword).
O NO é um importante mensageiro intercelular nos mamíferos superiores. O mecanismo de sinalização intercelular é, em geral, realizado através de receptores de membrana celular na célula alvo; estes receptores são, habitualmente, transmembranosos tendo contato com citoplasma e desencadeando uma "cascata" de sinais intracelulares que finalizarão em uma mudança na célula. Pelas suas características químicas de alta difusibilidade, a sinalização do NO é exercida diretamente em nível intracelular, sem receptores transmembranosos. Devido à sua penetração intracelular sem intermediários membranosos, o organismo utiliza o NO em funções fisiológicas em que é necessária uma resposta rápida.
O NO também faz parte do arsenal de primeira defesa do organismo com poder microbicida. Assim, está demonstrado sua ação antibactericida, antiparasítica e antiviral40- 44. Nestes casos, o NO atua em concentrações maiores do que as de mensageiro, sendo tóxico aos microorganismos invasores. Existe um tênue limite de concentração tissular entre a não-toxicidade às células do hospedeiro e a toxicidade necessária para ação antimicrobicida. No caso de doenças autoimunes e situações de sobrecarga exageradas do organismo, o NO encontra-se em concentrações tóxicas para as células do organismo. Portanto, o NO atua como toxina conforme a concentração e o tecido em questão, devendo ainda ser considerada a capacidade de depuração tecidual. A determinação destas concentrações tissulares relativas permanece um segredo da natureza.
A Tabela 2 resume algumas ações teciduais do NO como mensageiro ou toxina.
Neste artigo, nos restringiremos mais a alguns efeitos do NO como mensageiro nos mais importantes sistemas do organismo, devido à amplitude e complexidade do tema.
NO-mensageiro no sistema cardiovascular
Devido aos estudos pioneiros do EDRF por Furchgott, a ação vasoprotetora do NO é bem conhecida. O NO antagoniza as contrações da musculatura lisa vascular e inibe a ativação plaquetária. Atuando nas integrinas, o NO modifica a adesividade leucocitária e a diapedese dos neutrófilos5.
No endotélio vascular ocorre liberação continuada de NO, responsável pela manutenção do fluxo sangüíneo tecidual e controle do extravasamento tecidual. O NO-mensageiro produzido no endotélio tem função vasodilatadora fisiológica. Assim, durante exercício físico ocorre aumento do débito cardíaco e redistribuição do fluxo sangüíneo para musculatura esquelética e circulação coronariana. Este mecanismo é mediado pela cNOS (eNOS ou isoforma III) cuja expressão genética pode ser potencializada com exercícios aeróbios regulares24. Entretanto, a função principal do NO é a exercida no controle de adesão dos elementos sangüíneos (leucócitos e plaquetas) ao endotélio18. Por este mesmo mecanismo, o NO diminui a permeabilidade vascular. Em estudos com inibição da síntese de NO vascular utilizando-se o análogo L-NAME em intestino de gatos, observa-se aumento da permeabilidade microvascular às proteínas, mediadas por leucócitos e outras células inflamatórias (macrófagos, mastócitos, plaquetas, etc.)19.
No miocárdio existe também expressão da isoforma III ou eNOS, determinada por estudos de imunohistoquímica e técnicas de biologia molecular. A eNOS é encontrada nos miócitos, células endoteliais e músculo liso vascular sendo a nNOS achada nos neurônios. O NO-mensageiro atua como inotrópico negativo levando a aumento da dilatação diastólica, sem influenciar os índices de contractilidade miocárdica25.
Já a isoforma II ou iNOS pode ser produzida pelas mesmas células sob estímulos como, por exemplo, sepse, miocardite, rejeição de transplante. Quando ocorre aumento do NO em níveis elevados, o NO-toxina induz à disfunção cardíaca característica da síndrome de resposta inflamatória sistêmica (Systemic Inflamatory Response Syndrome - SIRS)26.
NO-mensageiro no sistema bronco-pulmonar
Em culturas de células alveolares pulmonares epithelium-like, Asano et al. demonstraram atividade basal de NOS constatada como isoforma I ou nNOS, não se constatando eNOS ou isoforma III. Sob estímulo de interferon-gama (IFN-g) isolado ou juntamente com interleucina-1b (IL-1b) mais fator de necrose tumoral- (TNF-a), observou-se produção de RNA-mensageiro da isoforma II ou iNOS21.
Considera-se o NO-mensageiro produzido a partir da cNOS como responsável pela homeostase das vias aéreas. O NO mantém o calibre brônquico e regula a freqüência dos movimentos ciliares; na circulação pulmonar, o NO equilibra a relação ventilação-perfusão.
A iNOS epitelial bronquial é responsável pela defesa imunológica a agentes externos inalados, principalmente pelo recrutamento de macrófagos epiteliais e subepiteliais22. Isto é configurado pela queda da resistência às infecções broncopulmonares com administração de análogos da L-arginina in vitro e in vivo3.
NO-mensageiro no sistema renal
O NO-mensageiro é sintetizado nos rins fisiologicamente, exercendo importantes funções de homeostase no fluxo sangüíneo e excreção renal. O bloqueio experimental da produção do NO-mensageiro leva à diminuição da irrigação renal e queda da eliminação de sódio27.
Nos rins foram isoladas as três isoformas da NOS. Na mácula densa constatou-se forte presença da isoforma I ou nNOS e nas arteríolas glomerulares foram encontradas as isoformas I e III (nNOS e eNOS) nos ramos aferentes e eferentes, com maior concentração nas arteríolas eferentes.
A presença da isoforma I é importante nos vasos aferentes pré-glomerulares pela estreita correlação com liberação de renina28. A ação do NO-mensageiro sobre a secreção do sistema renina-angiotensina tem óbvias implicações terapêuticas, porém os achados ainda são discrepantes.
Em um estudo com infusão de análogo da L-arginina (L-NMMA) em voluntários sadios, observou-se aumento da pressão arterial média (+7%) com aumento da resistência periférica (+36%) sem aumento do fluxo sangüíneo renal, porém com aumento da resistência vascular renal (+21%) seguido de queda da filtração glomerular (-19%), queda do fluxo urinário (-18%) e da eliminação de sódio (-25%), sem aumento da atividade plasmática da renina29. Outro estudo semelhante, porém em cães, em que foi utilizado a L-NAME juntamente com bloqueio da mácula densa pelo modelo de rim não-filtrante, mostrou que o bloqueio intrarenal de NO aumenta a secreção de renina em nível renal, ocorrendo elevação plasmática somente no rim com bloqueio simultâneo da mácula densa30. Portanto, à medida que as pesquisas avançam, surgem fortes indícios que o NO está envolvido no mecanismo da hipertensão arterial.
Na glomerulonefrite observa-se produção exagerada do NO que é devido à indução da iNOS, expressa a partir de macrófagos nos glomérulos27 com infiltração leucocitária e aumento da permeabilidade vascular com conseqüente proteinúria.
NO-mensageiro no sistema nervoso
O NO é um poderoso vasodilatador cerebral mantendo o fluxo cerebral basal e levando a aumento do fluxo sangüíneo sob atividade neuronal intensa32. A isoforma I ou nNOS foi historicamente purificada em cérebros de camundongo e porco10, como já citado. Porém, também a eNOS e a iNOS estão presentes e são importantes nas funções fisiológicas e estão envolvidas em várias patologias do SNC.
A nNOS e eNOS podem intercambiar funções, como demonstrado em estudos de camundongos modificados geneticamente com ausência da nNOS, em que se observou manutenção da função neurológica às custas da eNOS33.
O NO-mensageiro é considerado um neurotransmissor. Devido as suas características de alta difusibilidade, como já citado, o NO é capaz de penetrar no citoplasma celular diretamente, sem necessidade de receptores de membrana, levando a respostas rápidas e precisas .
O hipocampo é uma das áreas de pesquisa mais promissoras da ação do NO no SNC. Sabe-se que, quando o hipocampo é destruído em ambos hemisférios cerebrais, em mamíferos, os mesmos perdem a capacidade de reter memórias novas. Entre os múltiplos neurotransmissores excitatórios do SNC, destaca-se o glutamato. Entre os receptores de glutamato existem os receptores de N-metil-D-aspartato (NMDA) - pertencentes à subclasse de glutamato. Estes receptores recebem o nome de NMDA devido ao fato de serem estimulados seletivamente pelo N-metil-D-aspartato, que é um análogo do glutamato. Além dos receptores NMDA estarem preferencialmente situados no hipocampo, animais com bloqueio específico dos receptores NMDA perdem a capacidade de aprendizado.
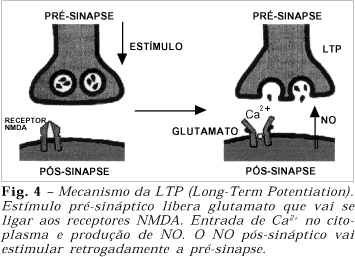
Nos receptores NMDA um estímulo pode levar à própria potencialização. Este mecanismo é conhecido como Long-Term Potentiation (LTP), consistindo de um sistema progressivo e prolongado do estímulo, mesmo com estímulo inicial em condições submáximas. O envolvimento do NO-mensageiro no mecanismo da LTP foi primeiramente demonstrado por Böhme, em um experimento com camundongos, em que o bloqueio de formação de NO levou à ausência de condicionamento e de resposta olfatória34,35.
Em nível de sinapse neuronal, após estímulo do neurônio à montante, ocorre liberação de glutamato que vai se ligar aos receptores NMDA. Enquanto persistir esta união (glutamato/receptor NMDA), o Ca2+ é capaz de entrar no citoplasma do neurônio à jusante levando à produção da nNOS. Assim, o NO é liberado na pós-sinapse após estímulo pré-sináptico submáximo, funcionando como mensageiro retrógrado para a pré-sinapse e reiniciando todo o processo (Fig. 4).
Os receptores NMDA estão envolvidos na memória tardia cuja indução pelo mecanismo da LTP culmina com alterações de proteínas celulares à jusante, resultando em aquisição de experiências e conhecimentos16. Esta linha de pesquisa é intensa devido às implicações na dificuldade de aprendizado em crianças e falhas de memória em adultos. Também existe esperança de melhor conhecimento da fisiopatologia da doença de Alzheimer.
O eqüivalente dos macrófragos no cérebro são as células microgliais. As células gliais em cultura mostram capacidade de indução da iNOS, mais fortemente presente quando há o estímulo tríplice de IFN-g, IL-1b e LPS bacteriano38. Existem fortes evidências que a iNOS pode estar envolvida, além dos processos bacterianos e virais, em processos neurodegenerativos e outras condições que levam à liberação de citoquinas no cérebro tais como traumas e isquemias39.
- 1. Green LC, Tannenbaum SR, Goldmann P. Nitrate synthesis in the germfree and conventional rat. Science 1981, 212: 56-58.
- 2. Stuehr DJ, Marletta MA. Mammalian nitrate biosynthesis: Mouse macrophages produce nitrite in response to Escherichia coli lipopolysaccharide. Proc Natl Acad Sci USA 1985, 82: 7738-7742.
- 3. Hibbs JB, Taintor RR, Vavrin Z. Macrophage cytotoxicity: Role of L-arginine deiminase and imino nitrogen oxidation to nitrite. Science 1987: 235: 473-476.
- 4. Marletta MA, Yoon PS, Iyengar R, Leaf CD, Wishnok JS. Macrophague oxidation of L-arginine to nitrite and nitrate: Nitric oxide is an intermediate. Biochemistry 1988: 27: 8706-8711.
- 5. Furchgott RF, Cherry PD, Zawakzki JV, Jothianandan D. Endothelial cells as mediators of vasodilation of arteries. J Cardiovasc Pharm 1984, 53: 557-573.
- 6. Ignarro LJ, Buga GM, Wood KS, Byrns RE, Chaudhuri G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc Natl Acad Sci USA vol 84, pp.9265-9269, 1987.
- 7. Snyder SH, Bredt DS. Biological role of nitric oxide. Science Am 1992, 266: 68-77.
- 8. Rees DD, Palmer RMJ, Schultz R, Hodson HF, Moncada S. Characterization of three inhibitors of endothelial nitric oxid synthase in vitro and in vivo. Brit J Pharm 1990, 101: 746-752.
- 9. Konturek SK, Konturek PC. Role of nitric oxid in the digestive system. Digestion 1995, 56: 1-13.
- 10. Schmidt HHHW, Pollock JS, Nakana M, Gorsky LD, Förstermann U, Murad F. Purification of a soluble isoform of guanylyl cyclase-activating-factor synthase. Proc Natl Acad Sci USA 1991, 88: 365-369.
- 11. Stuehr DJ, Cho HJ, Kwon NS, Weise MF, Nathan CF. Purification and characterization of the cytokine-induced macrophage nitric oxide synthase: an FAD- and FMN-containing flavoprotein. Proc Natl Acad Sci USA 1991, 88: 7773-7777.
- 12. Geller DA, Lowenstein CJ, Shapiro RA, Nussler AK, Di SM, Wang SC, Nakayama DK, Simmons RL, Snyder SH, Billiar TR. Molecular cloning and expression of inducible nitric oxide synthase from human hepatocytes. Proc Natl Acad Sci USA 1993, 90: 3491-3495.
- 13. Pollock JS, Förstermann U, Mitchell JA, Warner TD, Schmidt HHHW, Nakane M, Murad F. Purification and characterization of particulate endotelium-derived relaxing factor synthase from cultured and natived bovine aortic endothelial cells. Proc Natl Acad Sci USA 1991, 88: 10480-10484.
- 14. Zhang J, Dawson TM, Snyder SH. Nitric oxid activation of poly(ADP-ribose) synthase in Neurotoxicity. Science 1994, 263: 687-689.
- 15. Nathan C, Xie, QW. Regulation of biosynthesis of nitric oxide. J Biol Chem 1994, 269: 13725-13728.
- 16. Scmidt HHHW, Walter U. NO at work. Cell 1994, 78: 919-925.
- 17. Schuman EM, Madosom DV. A requirement for the intercellular messenger nitric oxide in long-term potentiation. Science 1991, 254: 1503-1506.
- 18. Kubes P, Suzuki M, Granger DN. Nitric oxide: an endogenous modulator of leukocyte adhesion. Proc Natl Acad Sci USA 1991, 88: 4651-4655.
- 19. Kubes P, Granger DN. Nitric oxide modulates microvascular permeability. American J Phys 1992, 262: H611-H615.
- 20. Nathan C, Xie Q. Nitric oxide synthases: Roles, tolls, and controls. Cell 1994, 78: 915-918.
- 21. Asano K, Chee CBE, Gaston B, Lilly CM, Gerard C, Drazen JM, Stamler J. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proc Natl Acad Sci USA 1994, 91: 10089-10093.
- 22. Nathan C. Natural resistance and nitric oxide. Cell 1995, 82: 873-876.
- 23. Nicholson S, Almeida MGB, Silva JRL, Nathan C, Xie Q, Mumford R, Weidner JR, Calaycay J, Geng J, Boechat N, Linhares C, Rom W, Ho JL. Inducible nitric oxide synthase in pulmonary alveolar macrophages from patients with tuberculosis. J Exp Med 1996, 183: 2293-2302.
- 24. Shen W, Zhang X, Wolin MS, Sessa W, Hintze TH. Nitric oxide production and NO synthase gene expression contribute to vascular regulation during exercise. Med Sci Sports Exerc 1995, 8: 1125-1134.
- 25. Hare JM, Colucci WS. Role of nitric oxide in the regulation of myocardial function. Prog Cardiovasc Dis 1995, 38: 155-166.
- 26. Smith TW, Balligand JL, Kaye DM, Wiviott SD, Simmons WW, Han X, Michel T, Singh K, Kelly RA. The Role of the NO pathway in the control of cardiac function. J Card Fail 1994, 4-Suppl: S141-S147.
- 27. Bachmann S, Mundel P. Nitric oxide in the kidney: synthesis, localization and function. American J Kidney Dis 1994, 24: 112-129.
- 28. Bachmann S, Bosse HM, Mundel P. Topography of nitric oxide synthesis by localizing constitutive NO synthases in mammalian kidney. American J Phys 1995, 5 Pt2: F885-898.
- 29. Haynes WG, Hand MF, Dockrell ME, Eadington DW, Lee MR, Hussein Z, Benjamin N, Webb DJ. Physiological role of nitric oxide in regulation of renal function in humans. American J Phys 1997, 272 Pt2: F364-F371.
- 30. Schnackenberg CG, Tabor BL, Strong MH, Granger JP. Inhibition of intrarenal NO stimulates renal secretion through a macula densa-mediated mechanism. American J Phys 1997, 272 Pt2: R879-R886.
- 31. Wang Y, Mardsen PA. Nitric oxide synthases: Biochemical and molecular regulation. Curr Opinion Nephr Hypert 1995, 4: 12-22.
- 32. Iadecola C. Regulation of the cerebral microcirculation during neural activity: Is nitric oxide the missing link? Trends Neurosci 1993, 16: 206-214.
- 33. Paakkari I, Lindsberg P. Nitric oxide in the central nervous system. Ann Medic 1995, 27: 369-377.
- 34. Böhme GA, Bon C, Lemaire M, Reibaud M, Piot O, Stutsmann JM, Doble A, Blanchard JC. Altered synaptic plasticity and memory formation in nitric oxide synthase inhibitor-treated rats. Proc Natl Acad Scien USA 1993, 90: 9191-9194.
- 35. Böhme GA, Bon C, Stutsmann JM, Doble A, Blanchard JC. Possible involvement of nitric oxide in long-term potentiation. European J Pharm 1991, 199: 379-381.
- 36. Balligand JL, Kelly RA, Marsden PA, Smith TW, Michel T. Control of cardiac muscle cell function by an endogenous nitric oxide signaling system. Proc Natl Acad Sci USA 1993, 90: 347-351.
- 37. Wiedner EB, Bao X, Altschuler SM. Localization of nitric oxide synthase in the brain stem neural circuit controlling esophageal peristalsis in rats. Gastroenterology 1995, 108: 367-375.
- 38. Zielasek J, Hartung HP. Molecular mechanisms of microglial activation. Adv Neuroimmunol 1996, 6: 191-222.
- 39. Sparrow JR. Inducible nitric oxide synthase in the central nervous system. J Mol Neurosci 1995, 5: 219-229.
- 40. Drapier JC, Weizesbin J, Hibbs JB. Interferon gamma and tumor necrosis factor induce the L-arginine cytotoxic effects or mechanism in murine macrophages. European J Immuno 1988, 18: 1587-1592.
- 41. Feng HM, Walker DH. Interpheron-gamma and tumor necrosis factor alpha exert their antirickettsial effect via induction of synthesis of nitric oxide. American J Physio 1993, 143: 1016-1023.
- 42. Oswald IP, Eltoum I, Wynn TA, Scwartz B, Caspar P, Paulin D, Sher A, James SL. Endothelial cells are activated by cytokine treatment to kill an intravascular parasite, Schistosoma mansoni. Proc Nat Acad Sci USA 1994, 91: 999-1003.
- 43. Gazzinelli RT, Oswald IP, Hieny S, James SL, Sher A. The microbicidal activity of interferon-gamma-treated macrophages against Trypanossoma cruzi involves an L-arginine-dependent, nitrogen oxide-mediated mechanism inhibitable by interleukin-10 and transforming growth factor-beta. European J Immuno 1992, 22: 2501-2506.
- 44. Lowenstein C, Allen G, Walker A, Rose N, Snyder S, Herskowitz A. Nitric oxide inhibits viral replication in myocarditis. Circulation 1993, 88: IG-IH (Abstr).
- 45. Teng B, Murthy KS, Kuemmerle JF, Grider JR, Sase K, Michel T, Makhlouf GM: Expression of endothelial nitric oxide synthase in human and rabbit gastrointestinal smooth muscle cells. Am J Physiol 275: G342-351, 1998.
Datas de Publicação
-
Publicação nesta coleção
16 Nov 2000 -
Data do Fascículo
Set 2000