Abstract
We present a documentation of the morphological details of two larval stages of mantis shrimps. Documentation was done using the autofluorescence capacities of the cuticle. This is the first time that morphological details of late mantis shrimp larvae are documented in great detail via photography, including all parts of the body up to the proximal elements of the appendages, and not presented as line drawings; it is the second time for mantis shrimp larvae in general. The description is presented as a standardized descriptive matrix. Documentation and description style are adjusted to facilitate comparison with fossil representatives of mantis shrimps, but also their extant counterparts, as well as specimens in the wider framework of Malacostraca and Eucrustacea. Through an exemplary comparison with fossil mantis shrimps, we provide indications about the early evolutionary history of the group. Through an out-group comparison, we identify several possible evolutionary changes of developmental timing, i.e., heterochrony, which could explain some morphological specialisations of mantis shrimps.
Keywords:
Verunipeltata; larva; erichthus; heterochrony; palaeo-evo-devo
INTRODUCTION
Mantis shrimps (Stomatopoda) are exceptional among malacostracans in regard to their morphology. Adults are equipped with a highly developed optical system allowing complex colour vision (e.g., Marshall et al., 1991Marshall, N.J.; Land, M.F.; King, C.A. and Cronin, T.W. 1991. The compound eyes of mantis shrimps (Crustacea, Hoplocarida, Stomatopoda). II. Colour pigments in the eyes of stomatopod crustaceans: polychromatic vision by serial and lateral filtering. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 334: 57-84. ) and specialized, highly efficient raptorial claws (e.g., Caldwell and Dingle, 1975Caldwell, R.L. and Dingle, H. 1975. Ecology and evolution of agonistic behavior in stomatopods. Naturwissenschaften, 62: 214-222. ).
Larval forms of mantis shrimps are also highly specialised, and possess an aberrant set of morphological traits among crustacean larvae. This includes their large, fully functional raptorial maxilliped 2, their often large overall body size (up to 50 mm in length), the elongated head region in some species, and the hypertrophied shield (e.g., Giesbrecht, 1910Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p. ).
Specialisations of larvae and adults may result from various evolutionary processes (Haug, in pressHaug, J. T. In press. Chapter 9. Metamorphosis in Crustaceans: Towards a Synthesis. p. X-X. In: K. Anger, S. Harzsch and M. Thiel (eds), Developmental Biology and Larval Ecology. The Natural History of the Crustacea, Vol. 7. Oxford, Oxford University Press. ), in many cases this process involves heterochrony. Heterochrony has relatively rarely been discussed as an explanation for the appearance of specialized structures of non-vertebrate species (Jirikowski et al., 2015Jirikowski, G.J.; Wolff, C. and Richter, S. 2015. Evolution of eumalacostracan development - new insights into loss and reacquisition of larval stages revealed by heterochrony analysis. EvoDevo, 6: 4.; examples in Webster and Zelditch, 2005Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.; Haug et al., 2010aHaug, J.T.; Maas, A. and Waloszek, D. 2010a. †Henningsmoenicaris scutula, †Sandtorpia vestrogothiensis gen. et sp. nov. and heterochronic events in early crustacean evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 100: 311-350.). So far, no study has focused on identifying heterochronic events in mantis shrimps. Studying heterochronic events in Stomatopoda is challenging, if based only on literature data for two main reasons:
-
Complete ontogenetic sequences are scarce (Hamano and Matsuura, 1987Hamano, T. and Matsuura, S. 1987. Egg size, duration of incubation, and larval development of the Japanese mantis shrimp in the laboratory. Nippon Suisan Gakkaishi, 53: 23-29.; Morgan and Goy, 1987Morgan, S.G. and Goy, J.W. 1987. Reproduction and larval development of the mantis shrimp Gonodactylus bredini (Crustacea: Stomatopoda) maintained in the laboratory. Journal of Crustacean Biology, 7: 595-618.; Ahyong et al., 2014Ahyong, S.T.; Haug, J.T. and Haug, C. 2014. Stomatopoda. p. 185-189. In: J.W. Martin; J. Olesen and J.T. Høeg (eds), Atlas of Crustacean Larvae. Baltimore, The Johns Hopkins University Press. ).
-
Existing illustrations are usually restricted to diagnostic features and line drawings. This restriction makes comparison in a wider phylogenetic frame, as well as in-group comparisons with extant or fossil specimens, difficult.
It seems therefore necessary to make use of a documentation technique that provides more details for morphological comparisons. Haug et al. (2011Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.) proposed the use of auto-fluorescence in combination with digital processing for documenting the external morphology of representatives of Euarthropoda. This technique proved suitable for morphological comparisons, since it effectively documents fossil (e.g., Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.; 2009aHaug, C.; Haug, J.T.; Waloszek, D.; Maas, A.; Frattigiani, R. and Liebau, S. 2009a. New methods to document fossils from lithographic limestones of southern Germany and Lebanon. Palaeontologia Electronica, 12: 12p. , bHaug, C.; Haug, J.T. and Waloszek, D. 2009b. Morphology and ontogeny of the Upper Jurassic mantis shrimp Spinosculda ehrlichi n. gen. n. sp. from southern Germany. Palaeodiversity, 2: 111-118.; 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.) as well as extant specimens (e.g., Haug et al., 2011Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.; 2012aHaug, C.; Sallam, W.S.; Maas, A.; Waloszek, D.; Kutschera, V. and Haug, J. T. 2012a. Tagmatization in Stomatopoda - reconsidering functional units of modern-day mantis shrimps (Verunipeltata, Hoplocarida) and implications for the interpretation of fossils. Frontiers in Zoology, 9: 31, 14p. ; Rötzer and Haug, 2015Rötzer, M.A.I.N. and Haug, J.T. 2015. Larval development of the European lobster and how small heterochronic shifts lead to a more pronounced metamorphosis. International Journal of Zoology, 2015: 345172, 17p. ; Eiler et al., 2016Eiler, S.M.; Haug, C. and Haug, J.T. 2016. Detailed description of a giant polychelidan Eryoneicus-type larva with modern imaging techniques. Spixiana, 39: 39-60.) in great detail.
In the present paper, we describe the morphology of larval stages of extant mantis shrimps, documented with the methodology outlined in Haug et al. (2011Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.), and compare them with fossil counterparts documented and described with the same methodology. Based on this, we provide a first attempt at identifying heterochronic events in mantis shrimp evolution.
MATERIAL AND METHODS
Material
Material was provided by Björn von Reumont, now Gießen. Originally it was collected by Nils Brenke, Wilhelmshaven, at the Great Meteor Seamount, Northeast Atlantic (collection site: 30°N 28.5°W), during the Meteor Expedition M42/3 in September 1998, with a Bongo plankton net in 20 meter depth. This study is based on 11 larval specimens of mantis shrimps from this collection. To differentiate developmental stages, we calculated a size index based on shield length scaled by pleon length. Specimens were then dissected using fine pincers and needles within the original storage liquid (70% ethanol); one of each pair of appendages was cut off and documented. Specimens are part of the collection of the Zoomorphology work group, LMU Munich. The specimens were originally identified as Pseudosquillopsis ceresii (Roux, 1828), but are more likely representatives of Lysiosquillidae, possibly of Lysiosquilla.
Documentation methods
Overviews of the entire animals were photographed in 70% ethanol on a Keyence BZ-9000 fluorescence microscope with a 2× objective (resulting in about 20× magnification). UV light (377 nm) was used for illumination, using the autofluorescence capacities of the specimens (see e.g., Haug et al., 2011Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.). The dissected appendages were documented using a Zeiss Axio Scope 2 microscope with reflective UV-light (wavelength: 358nm) and an AxioCam digital camera (Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.). An overview image of a stage 1 larva was recorded with a Canon EOS 450D and Canon EF-S 18-55mm 3.5-5.6 lens.
For high-resolution images, composite imaging was applied (Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.; 2009aHaug, C.; Haug, J.T.; Waloszek, D.; Maas, A.; Frattigiani, R. and Liebau, S. 2009a. New methods to document fossils from lithographic limestones of southern Germany and Lebanon. Palaeontologia Electronica, 12: 12p. ; 2011Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.). To increase the depth of field and the field of view, several stacks from adjacent areas were recorded, fused with the freely available software CombineZM/ZP. The fused images were then stitched using Adobe Photoshop CS3 or Microsoft Image Composite Editor.
Images were optimized in brightness and contrast in Adobe Photoshop CS3. The 'mask unsharp' filter was applied; this especially enhances the visibility of setae on the appendages. Photographs of some structures with setae were recorded twice, under different exposure times. The brighter image was then placed as a separate layer above the image with shorter exposure time in Adobe Photoshop. Alignment of the two layers was based on significant landmark structures. The magic wand tool was applied to mark overexposed areas; a high feather was applied to the edge before these areas were cut. The resulting image shows all parts well illuminated (see e.g., Haug et al., 2013aHaug, C.; Shannon, K. R.; Nyborg, T. and Vega, F. J. 2013a. Isolated mantis shrimp dactyli from the Pliocene of North Carolina and their bearing on the history of Stomatopoda. Bolétin de la Sociedad Geológica Mexicana, 65: 273-284.; Rötzer and Haug, 2015Rötzer, M.A.I.N. and Haug, J.T. 2015. Larval development of the European lobster and how small heterochronic shifts lead to a more pronounced metamorphosis. International Journal of Zoology, 2015: 345172, 17p. ).
Presentation
Description follows the descriptive matrix approach outlined by Haug et al. (2012bHaug, J.T.; Briggs, D.E.G. and Haug, C. 2012b. Morphology and function in the Cambrian Burgess Shale megacheiran arthropod Leanchoilia superlata and the application of a descriptive matrix. BMC Evolutionary Biology, 12: 162, 20p.). The entire description of the morphological structures is presented as a descriptive matrix (Appendix). Simplified representations were made in Adobe Illustrator CS3.
Evolutionary reconstruction
Character reconstruction was based on earlier phylogenetic reconstructions. As a basis, a simplified phylogeny was used, largely based on Ahyong and Harling (2000Ahyong, S.T. and Harling, C. 2000. The phylogeny of the stomatopod Crustacea.Australian Journal of Zoology, 48: 607-642.), Richter and Scholtz (2001Richter, S. and Scholtz, G. 2001. Phylogenetic analysis of the Malacostraca (Crustacea). Journal of Zoological Systematics and Evolutionary Research, 39: 113-136. ), Schram (2007Schram, F. R. 2007. Paleozoic proto-mantis shrimp revisited. Journal of Paleontology, 81: 895-916.), and Haug et al. (2010bHaug, J.T.; Waloszek, D.; Haug, C. and Maas, A. 2010c. High-level phylogenetic analysis using developmental sequences: The Cambrian †Martinssonia elongata, †Musacaris gerdgeyeri gen. et sp. nov. and their position in early crustacean evolution. Arthropod Structure & Development, 39: 154-173.; 2015Haug, C.; Wiethase, J. H. and Haug, J. T. 2015. New records of Mesozoic mantis shrimp larvae and their implications on modern larval traits in stomatopods. Palaeodiversity, 8: 121-133.). Reconstruction follows strict outgroup comparison and the general argumentation scheme of phylogenetic systematics in the sense of Hennig (1966Hennig, W. 1966. Phylogenetic Systematics. Urbana, Univ. Illinois Press.), Ax (1995Ax, P. 1995. Das System der Metazoa: ein Lehrbuch der phylogenetischen Systematik 1. Stuttgart, Gustav Fischer, 226p.) and Reif (2002Reif, W. E. 2002. Evolution of organ systems: phylogeny, function and reciprocal illumination. Senckenbergiana lethaea, 82: 356-366.; 2005Reif, W. E. 2005. Problematic issues of cladistics: 9. Hennig’s “Phylogenetic Systematics”. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 235: 289-342.). Based on comparison of states at specific nodes, heterochronic events were identified (Haug et al., 2010aHaug, J.T.; Maas, A. and Waloszek, D. 2010a. †Henningsmoenicaris scutula, †Sandtorpia vestrogothiensis gen. et sp. nov. and heterochronic events in early crustacean evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 100: 311-350.).
RESULTS
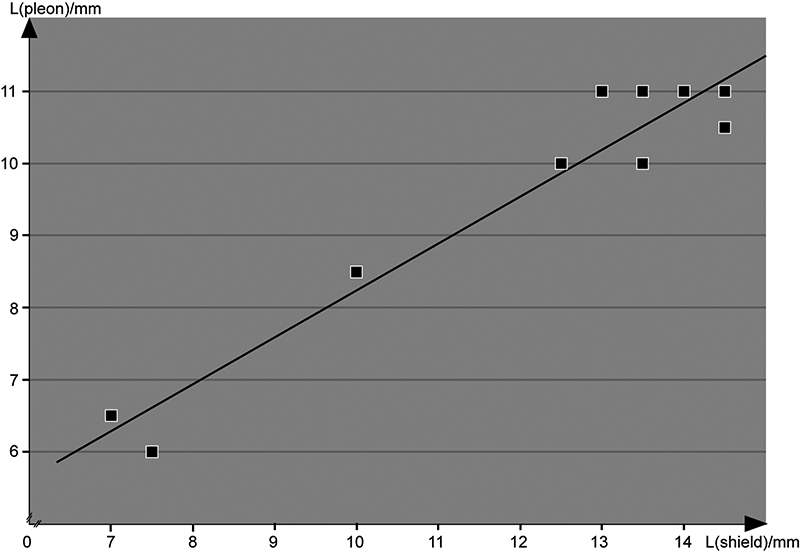
The specimens investigated fall into three size classes (Tab. 1; Fig. 1). To facilitate a reference to these size classes, we apply an open nomenclature. All specimens clearly represent larvae of the erichthus type; we consecutively number the stages without implying that these numbers represent the actual instar; it is in fact likely that we have only late instars and lack early ones. Based on observed morphology, we suggest that the smallest specimen represents the earliest developmental stage present in the material, erichthus stage 1 (Fig. 2), followed by a next larger size, erichthus stage 2 (Fig. 3), followed by the largest of the three, erichthus stage 3 (Fig. 4). Stage 3 potentially represents the ultimate larval stage (based on the morphological similarities to ultimate larval stages in larval sequences known from the literature, e.g., Pyne, 1972Pyne, R.R. 1972. Larval development and behaviour of the mantis shrimp, Squilla armata Milne Edwards (Crustacea: Stomatopoda). Journal of Royal Society of New Zealand, 2: 121-146. or Hamano and Matsuura, 1987Hamano, T. and Matsuura, S. 1987. Egg size, duration of incubation, and larval development of the Japanese mantis shrimp in the laboratory. Nippon Suisan Gakkaishi, 53: 23-29.). For stage 2 only an overview image was recorded (Fig. 3), and no appendages were dissected, in order to keep this single specimen for later reference.
Investigated specimens of mantis shrimp larvae; measurements and suggested developmental stage.

Diagram of approximate shield length vs approximate pleon length measured on 11 specimens of mantis shrimp larvae.

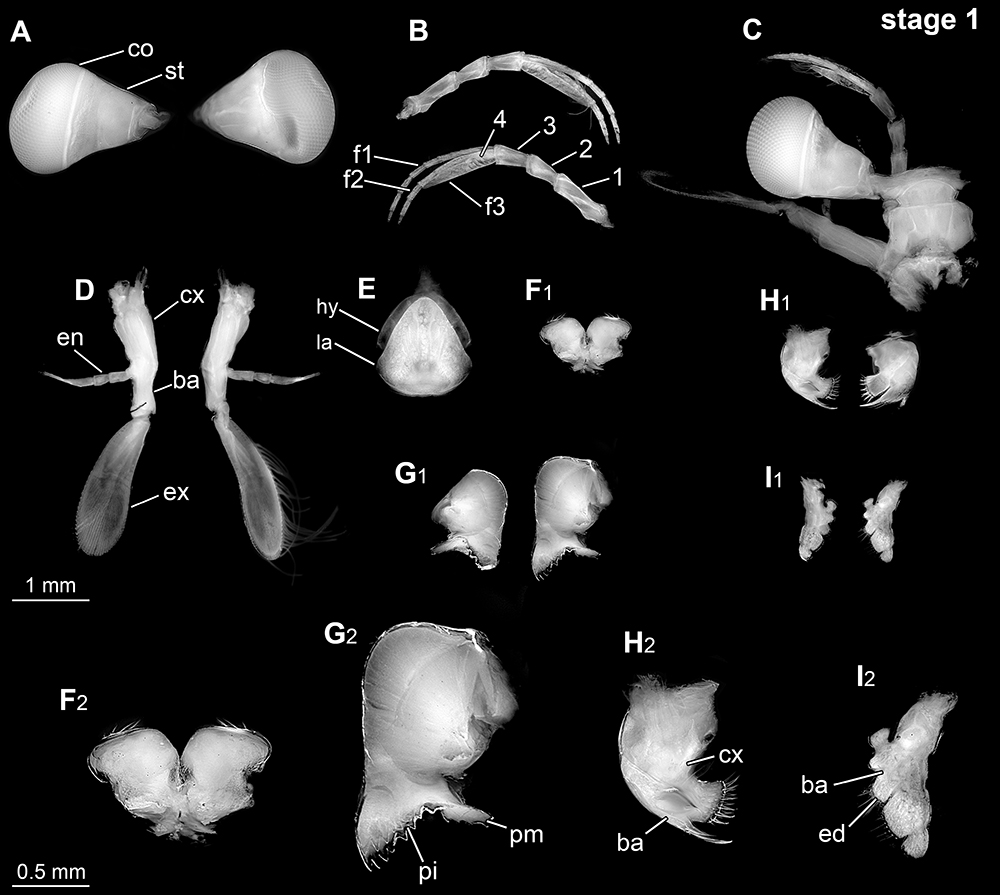
Erichthus stage 1. Overview in ventral view (left) with colour-marked appendages (right). Macro-image in grayscale, pleon inverted for providing more contrast. ant: antenna, atl: antennula, ce: compound eye, mp: maxilliped, plp: pleopod, tp: thoracopod, up: uropod.

Erichthus stage 2. Overview in ventral view (left) with colour-marked appendages (right). Composite autofluorescence images. ant: antenna, atl: antennula, ce: compound eye, mp: maxilliped, plp: pleopod, tp: thoracopod, up: uropod.

Erichthus stage 3. Overview in ventral view (left) with colour-marked appendages (right). Composite autofluorescence images. ant: antenna, atl: antennula, ce: compound eye, mp: maxilliped, plp: pleopod, tp: thoracopod, up: uropod.
Detailed documentation of the appendages shows a size and developmental transition from stage 1 (Figs. 5-7) to stage 3 (Figs. 8-10). Comparison of the tailfan supports further the developmental succession from erichthus stage 1 to stage 3 (Fig. 11).

Erichthus stage 1. Eyes and appendages of the head region. For all appendages: left/upper in anterior view; right/lower in posterior view. A: compound eyes; B: antennula; C: dorsal overview of isolated head region; D: antenna; E: hypostome-labrum complex, ventral view; F: paragnaths, ventral view; G: mandibles; H: maxillula; I: maxilla; F2-I2: enlarged versions of F1-I1. Composite autofluorescence images. 1-4: main elements 1-4, ba: basipod, co: cornea, cx: coxa, en: endopod, ex: exopod, f1-3: flagellum 1-3, hy: hypostome, la: labrum, pi: pars incisivus, pm: pars molaris, st: eye stalk.

Erichthus stage 1. Appendages of post-ocular segments 6-10. A-E: maxilliped 1-5. Left/upper in anterior view; right/lower in posterior view. Composite autofluorescence images. 1-6: main element 1-6, ep: epipod.

Erichthus stage 1. Appendages of post-ocular segments 11-18. A-C: thoracopod; A2-C2: enlarged versions of A1-C1; D-H: pleopod; I: uropod. Left in anterior view; right in posterior view. Composite autofluorescence images. ba: basipod, cx: coxa, en: endopod, ex: exopod, g: gills, vp: vermicular process.

Erichthus stage 3. Eyes and appendages of the head region. For all appendages: left/upper in anterior view; right/lower in posterior view; exception F: right oblique. A: compound eyes; B: antennula; C: antenna; D: dorsal overview of isolated head region; E: hypostome-labrum complex, ventral view; F: mandibles; G: maxillula; H: maxilla; F2-I2: enlarged versions of F1-I1. Composite autofluorescence images. 1-4: main elements 1-4, ba: basipod, co: cornea, cx: coxa, en: endopod, ex: exopod, f1-3: flagellum 1-3, hy: hypostome, la: labrum, pi: pars incisivus, pm: pars molaris, st: eye stalk.

Erichthus stage 3. Appendages of post-ocular segments 6-10. A-E: maxilliped 1-5. Left in anterior view; right in posterior view. Composite autofluorescence images. 1-6: main element 1-6, ep: epipod.

Erichthus stage 3. Appendages of post-ocular segments 11-18. A-C: thoracopod; D-H: pleopod; I: uropod. Left in anterior view; right in posterior view. All composite autofluorescence images. ba: basipod, cx: coxa, en: endopod, ex: exopod, g: gills, vp: vermicular process.

Tailfans of erichthus stages 1 and 3. Tailfan with telson and uropods in dorsal/posterior (top) and ventral/anterior (bottom) view. A: tailfan of erichthus stage 1. larva; B: tailfan of erichthus stage 3. Composite autofluorescence images. an: anus, te: telson, u: uropod.
The detailed morphological description which the discussion is based on is provided as a descriptive matrix in the Appendix Appendix Descriptive matrix for larvae of erichthus stage 1 and 3. Structure Description Erichthus stage 3 Erichthus stage 1 Body organised in 20 segments, 1 ocular segment followed by 19 appendage-bearing segments. x x Ocular segment x x bearing stalked compound eyes and rostral plate dorsally. x x Compound eye x x differentiable into two parts. x x Proximal part of compound eye (stalk) x x truncated cone-shaped. x x About 0.8× as long (proximo-distally) as distal margin of the element. x About 0.9× as long (proximo-distally) as distal margin of the element. x Distal part of compound eye (cornea) x x proximally-truncated ovoid like. x x about 0.6× as long (proximo-distally) as wide (latero-medially). x x with cornea with about 35 proximo-distally rows of ommatidia in anterior view. x x with up to about 20 ommatidia per proximo-distally row of cornea in anterior view visible. x x Appendage derivative of ocular segment x x hypostome-labrum complex. x x Labrum x x lanceolate in ventral view. x x about 0.9× as long (along posterior-anterior axis) as posterior-facing margin of the element. x x Hypostome x x overlapped by labrum in ventral view; anterior facing margin projecting under labrum. x x 1st post-ocular segment x x Appendage of 1st post-ocular segment (antennula) x x with 4 main elements and 3 flagella. x x 1st main element x x tube-shaped. x x about 3.2× as long (proximo-distally) as wide (latero-medially). x about 3.6× as long (proximo-distally) as wide (latero-medially). x 2nd main element x x truncated cone-shaped. x x about 0.5× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x about 0.6× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x 3rd main element x x truncated cone-shaped. x x about 0.5× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x about 0.6× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x about 0.7× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. 4th main element x x tube-shaped. x x not yet set off from flagella 2 and 3. x set off from flagella 2 and 3. x about 0.2× as long (proximo-distally) as 1st main element. Distal margin about 0.6× as wide (latero-medially) as that of 1st element. x about 0.4× as long (proximo-distally) as 1st main element. Distal margin about 0.8× as wide (latero-medially) as that of 1st element. x with single setae distally. Flagellum 1 x x arising from element 3. x x subdivided into 6 elements. x subdivided into about 8 elements. subdivided into about 32 elements. x with proximal element showing indications of future subdivision into 4 elements. x set off from antennula element 4 with apparent joints. x x Flagellum 2 arising from element 4. x x subdivided into 4 elements. x subdivided into about 27 elements. x with proximal element showing indications of future subdivision into 3 elements. x Flagellum 3 x x arising from element 4. x x undivided. x subdivided into about 14 elements. x about as long as the proximal element of flagellum 2. x with about 2 setae on the distal margin of every single element medially. x x 2nd post-ocular segment. x x Appendage of 2nd post-ocular segment (antenna) x x with coxa, basipod, endopod and exopod. x x Coxa x x tube-like shaped. x truncated-cone shaped. x about 0.9× as long (proximo-distally) as wide (latero-medially). x about 1.4× as long (proximo-distally) as wide (latero-medially). x about 2× as long (proximo-distally) as maximum width of the element (latero-medially). x Basipod x x tube-shaped. x truncated-cone shaped. x not clearly set off from coxa. x about 1.5× as long (proximo-distally) as wide (latero-medially). x x bearing endopod proximally and exopod distally. x x Endopod x x consisting of 3 main elements and flagellum. x x 1st main element x x tube-shaped. x x about 0.5× as long (proximo-distally) as wide (latero-medially). x about 0.8× as long (proximo-distally) as wide (latero-medially). x 2nd main element x x tube-shaped. x x about 2.5× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x about 2.9× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x 3rd main element x x truncated cone-shaped. x x about 3.3× as long (proximo-distally) as 1st main element. Distal margin about 1.3× as wide (latero-medially) as that of 1st element. x about 3.5× as long (proximo-distally) as 1st main element. Distal margin as wide (latero-medially) as that of 1st element. x Flagellum x x subdivided in about 3 distal elements and one large, undivided element following proximally. x subdivided in about 25 elements. x about 1.5× as long (proximo-distally) as total length of main elements 1-3 of the endopod. x about 1.7× as long (proximo-distally) as total length of main elements 1-3 of the endopod. x Exopod x x subelliptic in anterior view. divided in proximal and distal part. x x Proximal part x x tube-shaped. x tube-shaped; twisted. x about 1.3× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 3.3× as long (proximo-distally) as maximum width of the element (medio-laterally). x Distal Part x x paddle-like shaped. x subelliptic in anterior view. x x with about 45 setae indicated by insertions. x with about 80 setae indicated by insertions. x about 3× as long (proximo-distally) as maximum width (latero-medially) of the element. x about 3.4× as long (proximo-distally) as maximum width (latero-medially) of the element. x 3rd post-ocular segment x x Appendage of 3rd post-ocular segment (mandible) x x without palpus. x x with toothed part divided in pars incisivus and pars molaris by an almost orthogonal angle. x x Pars incisivus x x as single curved row. x x with about 8 teeth. x x Pars molaris x x with single- and two-rowed part. x x with single row proximal to pars incisivus and bearing about 5 teeth. x x with two-rowed part distal to pars incisivus and bearing about 10 teeth. x x Sternal outgrowth of 3rd post-ocular segment x paragnath x x bipartite. x lobate; about 1.1× as long (proximo-distally) as maximum width (latero-medially). x guiding the movement of the mandibles. x with both parts bearing about 3 setae at proximal margin. x 4th post-ocular segment x x dorsally contributing to the headshield. x x Appendage of 4th post-ocular segment (maxillula) x x divided in coxa with coxal endite and basipod with basipodal endite. x x coxa x x subelliptic in ventral view (without the endite); with almost straight medial margin. x subrectangular in ventral view (without the endite). x about as long (proximo-distally) as maximum width (medio-laterally). x about 1.2× as long (proximo-distally) as maximum width (medio-laterally). x bearing lobe-like endite at the distal part of the medial margin. x x coxal endite x x subrectangular in posterior view; as medial outgrowth of coxa. x x about 0.9× as long (medio-laterally) as maximum width (proximo-distally). x about 1.2× as long (medio-laterally) as maximum width (proximo-distally). x with distal margin bearing about 15 movable spines. x x basipod x x subtriangular in posterior view (without endite). x subtrapezoid in posterior view (without endite). x with distal margin about 0.5× as wide (latero-medially) as proximo-distal axis of the element. x with proximal margin about 0.6× as wide (latero-medially) as proximo-distal axis of the element. x with set off, lobe-like, subtriangular appendage (endo-/exopod?) on distal part of lateral margin. x with set off, subelliptic appendage (endo-/exopod?) on distal part of lateral margin. x with appendage (endo-/exopod?) about 2.3× as long (proximo-distally) as its proximal margin; with two setae visible on distal margin. x with appendage (endo-/exopod?) about 3× as long (proximo-distally) as maximum with of the element; without visible setae on distal margin. x basipodal endite x x subtriangular in ventral view. x x with one large spine and two slightly smaller/thinner spines on the distal end of the element. x x with largest distal spine about 0.3× as long as the whole basipod (proximo-distally). x with largest distal spine about 0.4× as long as the whole basipod (proximo-distally). x 5th post-ocular segment x x dorsally contributing to the headshield. x x Appendage of 5th post-ocular segment (maxilla) x x divided in coxa, basipod and endopod (?). x x coxa not yet set off from basipod; with endopod (?) distally. x coxa x x with one lobe-like endite on medial margin. x x with lobe-like endite bearing about 5 setae. x with lobe-like endite bearing about 10 setae. x with basal part subrectangular in proximal view; about 0.5× as long (proximo-distally) as maximum width (medio-laterally). x basipod x with two lobe-like endites on medial margin. x with proximal lobe-like endite bearing about 5 setae and distal endite bearing about 10 setae. x with basal part subrectangular in proximal view; about 0.4× as long (proximo-distally) as maximum width of the element (medio-laterally). x coxa + basipod x x as complex structure; basal part subrectangular with 3 lobe-like endites on medial margin. x About 2× as long (proximo-distally) as maximum width of the element (medio-laterally) (without the outgrowths). x with about 10 setae visible on medial margin of the third (distal) outgrowth. x endopod (?) x x appears bipartite. x x proximal part x x subrectangular on ventral view; with almost straight lateral margin and rounded medial margin. x x with about 5 setae visible on medial margin. x with about 15 setae visible on medial margin. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about as long (proximo-distally) as maximum width of the element (medio-laterally). x distal part x x lobe-like in ventral view. x x without visible setae. x with about 15 setae visible on medial margin. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.3× as long (proximo-distally) as maximum width of the element (medio-laterally). x 6th post-ocular segment x x Appendage of 6th post-ocular segment (maxiliped 1) x x consisting of 6 main elements (numbered form distal to proximal). x x 1st main element x x claw-like, poorly developed. x x not bearing any spines. x about 2× as long (proximo-distally) as proximal margin of the element. x about 2.3× as long (proximo-distally) as proximal margin of the element. x about 0.3× as long (proximo-distally) as lateral margin of 2nd main element. x x 2nd main element x x elliptic in lateral view. x subcircular in lateral view. x with almost straight inner (posterior-facing) margin. x bearing about 10 setae. x bearing about 60 setae. x about 1.3× as long (proximo-distally) as maximum width of the element (antero-posteriorly). x about 1.5× as long (proximo-distally) as maximum width of the element (antero-posteriorly). x 3rd main element x x tube-shaped. x x bearing about 20 setae. x bearing about 50 setae. x about 3.2× as long (proximo-distally) as wide (latero-medially). x about 5.4× as long (proximo-distally) as wide (latero-medially). x 4th main element x x slightly curved inwards. x x tube-like shaped; with thickened distal end. x x without visible setae. with about 5 setae visible on medial margin. x with medial and distal margin bearing about 20 setae. x about 7.5× as long (proximo-distally) as wide (latero-medially). x about 8.3× as long (proximo-distally) as wide (latero-medially). x with distal margin about 1.5× as wide as proximal margin (latero-medially). x with distal margin about 2× as wide as proximal margin (latero-medially). x 5th main element x x tube shaped, strongly curved inwards. x x about 8× as long (proximo-distally) as wide (latero-medially). x about 10× as long (proximo-distally) as wide (latero-medially). x 6th main element x x tube-shaped. x truncated-cone shaped. x about 2.2× as long (proximo-distally) as wide (latero-medially). x about 2.3× as long (proximo-distally) as proximal margin of the element. x Epipod x x kidney-shaped in lateral view. x x leaf-like arising from proximo-lateral edge of 6th main element. x x about 0.7× as long (proximo-distally) as wide (latero-medially). x about 0.9× as long (proximo-distally) as wide (latero-medially). x missing. 7th post-ocular segment x x Appendage of 7th post-ocular segment (maxiliped 2) x x consisting of 6 main elements (numbered from distal to proximal). x x 1st main element x x claw-like, strongly developed. x x straight, spear-like and slender. not bearing any spines. x with distal end slightly curved inwards. curved inwards. x x about 5.5× as long (proximo-distally) as proximal margin of the element. x about 6.7× as long (proximo-distally) as proximal margin of the element. x about 2.1× as long (proximo-distally) as maximum width of the element (latero-medially). x about 0.9× as long (proximo-distally) as lateral margin of 2nd main element. x x 2nd main element x x elliptic in lateral view. x x about 2.2× as long (proximo-distally) as maximum width of the element (latero-medially). x about 2.5× as long (proximo-distally) as maximum width of the element (latero-medially). x with lateral margin bearing about 30 small spines. x with lateral margin bearing about 60 small spines. x with small groove at the lateral margin at the height of the distal end of main element 1 when folded back. x x with triangular spine projecting at the proximal margin from the groove. x x with proximal margin of the spine about 0.6× as wide (latero-medially) as proximo-distally axis of the spine. x x 3rd main element x x bell-shaped. x x with distal margin about 0.9× as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about as wide (latero-medially) as proximo-distal axis of the element. x 4th main element x x subelliptic in lateral view, with saddle on distal half of the element. x x proximally tapered tube-like, with saddle on distal half of the element. slightly curved outwards. x about 2.7× as long (proximo-distally) as maximum width of the element (latero-medially). x about 2.9× as long (proximo-distally) as maximum width of the element (latero-medially). x with saddle about 0.1× as deep (latero-medially) as the maximum width of the element (latero-medially). x with saddle about 0.2× as deep (latero-medially) as latero-medially axis of the element. x 5th main element x x truncated cone-shaped. x x with distal margin about 0.4× as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about 0.5× as wide (latero-medially) as proximo-distal axis of the element. x 6th main element x x tube-shaped. x x about 1.5× as long (proximo-distally) as wide (latero-medially). x about 1.7× as long (proximo-distally) as wide (latero-medially). x Epipod x x missing. x subcircular in lateral view. x leaf-like arising from proximo-lateral edge of 6th main element. x about 0.6× as long (proximo-distally) as maximum width of the element (latero-medially). x 8th post-ocular segment x x Appendage of 8th post-ocular segment (maxiliped 3) x x consisting of 6 main elements (numbered form distal to proximal). x x 1st main element x x claw-like. x x not bearing any spines. x about 4.2× as long (proximo-distally) as proximal margin of the element. x about 4.3× as long (proximo-distally) as proximal margin of the element. x about 0.8× as long (proximo-distally) as lateral margin of 2nd main element. x 2nd main element x x subcircular in lateral view. x x about 1.1× as long (proximo-distally) as maximum width of the element (latero-medially). x about 1.3× as long (proximo-distally) as wide (latero-medially). x with almost straight distal and proximal margin. x with almost straight medial margin. x with proximal margin of the spine about 0.5× as wide (latero-medially) as proximo-distally axis of the spine. x with about 5 setae visible. x with about 15 setae visible. x with single spine projecting from proximal part of the lateral margin. x x with proximal margin of the spine about 0.3× as wide (latero-medially) as proximo-distally axis of the spine. x with spine about 0.1× as long as lateral margin of the 2nd main element. x x 3rd main element x x truncated cone-shaped. x x with distal margin about 0.8× as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about 1.2× as wide (latero-medially) as proximo-distal axis of the element. x bearing about 5 setae. x bearing about 10 setae. x with single spine projecting from the distal end of the lateral margin. x x with proximal margin of the spine about 0.2× as wide (latero-medially) as proximo-distally axis of the spine. x x with spine about 0.3× as long as lateral margin of the 3rd main element. x x 4th main element x x tube-like shaped, slightly curved outwards. x truncated cone-shaped, slightly curved inwards. x with distal margin about 0.3× as wide (latero-medially) as proximo-distal axis of the element. x about 4.2× as long (proximo-distally) as wide (latero-medially). x without visible setae. x with about 3 setae visible on medial margin. x 5th main element x x tube shaped, slightly curved inwards. x tube-like shaped, slightly s-shaped. x about 8.5× as long (proximo-distally) as wide (latero-medially). x about 8.8× as long (proximo-distally) as wide (latero-medially). x 6th main element x x tube shaped. x truncated cone-shaped. x with proximal margin about 0.9× as wide (latero-medially) as proximo-distal axis of the element. x about 2.8× as long (proximo-distally) as wide (latero-medially). x Epipod x x missing. x heart-like shaped in lateral view. x about 0.7× as long (proximo-distally) as wide (latero-medially). x leaf-like arising from proximo-lateral edge of 6th main element. x with distinct vein-like structure central and surrounding the epipod. x 9th post-ocular segment x x Appendage of 9th post-ocular segment (maxiliped 4) x x consisting of 6 main elements (numbered form distal to proximal). x x 1st main element x x claw-like. x x not bearing any spines. x about 5× as long (proximo-distally) as proximal margin of the element. x x about 0.7× as long (proximo-distally) as lateral margin of 2nd main element. x about 0.8× as long (proximo-distally) as lateral margin of 2nd main element. x 2nd main element x x subcircular in lateral view. x x with almost straight distal and proximal margin. x about as long (proximo-distally) as wide (latero-medially). x about 1.2× as long (proximo-distally) as wide (latero-medially). x without visible spines on lateral margin. x with lateral margin bearing about 10 small spines and two slightly larger spines. x without visible setae. x about 20 setae visible. x 3rd main element x x truncated cone-shaped. x x without visible setae. x with distal margin about as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about 0.8× as wide (latero-medially) as proximo-distal axis of the element. x with about 10 setae visible. x without visible spines on lateral margin. x with single spine projecting from the distal end of the lateral margin. x with proximal margin of the spine about 0.2× as wide (latero-medially) as proximo-distally axis of the spine. x with spine about 0.3× as long as lateral margin of the 3rd main element. x 4th main element x x tube-like shaped. x truncated cone-shaped. x with distal margin about 0.4× as wide (latero-medially) as proximo-distal axis of the element. x about 2.3× as long (proximo-distally) as wide (latero-medially). x without visible setae. x with about 10 setae visible on medial margin. x 5th main element x x tube shaped, slightly curved outwards. x x about 5.9× as long (proximo-distally) as wide (latero-medially). x about 8.3× as long (proximo-distally) as wide (latero-medially). x 6th main element x x tube-like shaped. x truncated cone-shaped. x with proximal margin about as wide (latero-medially) as proximo-distal axis of the element. x about 1.6× as long (proximo-distally) as wide (latero-medially). x Epipod x x heart-like shaped in lateral view. x x leaf-like arising from proximo-lateral edge of 6th main element. x x with distinct vein-like structure central and surrounding the epipod. x about 0.8× as long (proximo-distally) as wide (latero-medially). x about 0.9× as long (proximo-distally) as wide (latero-medially). x 10th post-ocular segment x x Appendage of 10th post-ocular segment (maxiliped 5) consisting of 6 main elements (numbered from distal to proximal). x x 1st main element x x claw-like. x x not bearing any spines. x about 0.6× as long (proximo-distally) as lateral margin of 2nd main element. x x about 2.3× as long (proximo-distally) as proximal margin of the element. x about 3.6× as long (proximo-distally) as proximal margin of the element. x 2nd main element x x subelliptic in lateral view. x x about 1.3× as long (proximo-distally) as wide (latero-medially). x about 1.7× as long (proximo-distally) as wide (latero-medially). x without visible setae. x bearing about 10 setae. x without visible spines on lateral margin. x with single spine projecting from proximal part of the lateral margin. x with proximal margin of the spine about 0.4× as wide (latero-medially) as proximo-distally axis of the spine. x with spine about 0.1× as long as lateral margin of the 2nd main element. x 3rd main element x x truncated cone-shaped. x x with distal margin about 0.9× as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about as wide (latero-medially) as proximo-distal axis of the element. x without visible setae. x bearing about 5 setae. x without visible spines on lateral margin. x with single spine projecting from the distal end of the lateral margin. x with proximal margin of the spine about 0.2× as wide (latero-medially) as proximo-distally axis of the spine. x with spine about 0.3× as long as lateral margin of the 3rd main element. x 4th main element x x truncated cone-shaped. x tube-like shaped. x with distal margin about 0.4× as wide (latero-medially) as proximo-distal axis of the element. x with distal margin about 0.6× as wide (latero-medially) as proximo-distal axis of the element. x without visible setae. x with about 5 setae visible on medial margin. x 5th main element x x tube shaped, curved outwards. x x about 4.6× as long (proximo-distally) as wide (latero-medially). x about 7.3× as long (proximo-distally) as wide (latero-medially). x without visible setae. x with about 10 setae visible on medial margin. x 6th main element x x truncated cone-shaped. x tube shaped. x about 2.3× as long (proximo-distally) as wide (latero-medially). x with proximal margin about 0.7× as wide (latero-medially) as proximo-distal axis of the element. x Epipod x x missing. heart-like shaped in lateral view. x x with distinct vein-like structure central and surrounding the epipod. x leaf-like arising from proximo-lateral edge of 6th main element. x x about 0.7× as long (proximo-distally) as wide (latero-medially). x about 0.9× as long (proximo-distally) as wide (latero-medially). x 11th post-ocular segment x x Appendage of 11th post-ocular segment (thoracopod 6) x x as bifid, but undivided structure. Future subdivision into coxa, basipod, endopod and exopod indicated, but no functional joints apparent. x with coxa, basipod, endopod and exopod. x Coxa x x tube-shaped to slightly conical (tapered distally). x tube-shaped. x about 1.2× as long (proximal-distally) as wide (latero-medially). x with proximal margin about 2× as wide (latero-medially) as proximo-distal axis of the element. x Basipod x x tube-shaped. x x about 2× as long (proximal-distally) as wide (latero-medially). x about 4.1× as long (proximal-distally) as wide (latero-medially). x Inner branch (endopod?) x x as undifferentiated lobe. x as a single, elongate paddle. x about 4× as long (proximal-distally) as wide (latero-medially). x about 6× as long (proximal-distally) as wide (latero-medially). x about 0.6× as long (proximal-distally) as outer branch. x about 0.7× as long (proximal-distally) as outer branch. x Outer branch (exopod?) x x as undifferentiated lobe. x about 2.5× as long (proximal-distally) as wide (latero-medially). x bipartite. x Proximal element of outer branch x tube-shaped. x with distal margin drawn-out and slightly overlapping distal element of outer branch (in medial view). x about 4.6× as long (proximal-distally) as wide (latero-medially). x Distal element of outer branch x paddle-shaped. x about 3× as long (proximal-distally) as wide (latero-medially). x 12th post-ocular segment x x Appendage of 12th post-ocular segment (thoracopod 7) x x as bifid, but undivided structure. Future subdivision into coxa, basipod, endopod and exopod indicated, but no functional joints apparent. x vermicular, undifferentiated, hardly developed. with coxa, basipod, endopod and exopod. x Coxa x x tube-shaped. x tube-shaped to slightly conical (tapering distally). x about 1.1× as long (proximal-distally) as wide (latero-medially). x with proximal margin about 1.8× as wide (latero-medially) as proximo-distal axis of the element. x Basipod x x tube-shaped. x x about 2× as long (proximal-distally) as wide (latero-medially). x about 4.7× as long (proximal-distally) as wide (latero-medially). x Inner branch (endopod?) x x as undifferentiated lobe. x about 3.3× as long (proximal-distally) as wide (latero-medially). x about 0.7× as long (proximal-distally) as outer branch. x as a single, elongate paddle. x about 4× as long (proximal-distally) as wide (latero-medially). x about 0.6× as long (proximal-distally) as outer branch. x Outer branch (exopod?) x x as undifferentiated lobe. x about 3.3× as long (proximal-distally) as wide (latero-medially). x bipartite. x Proximal element of outer branch x tube-shaped. x with distal margin drawn-out and slightly overlapping distal element of outer branch (in medial view). x about 5.6× as long (proximal-distally) as wide (latero-medially). x Distal element of outer branch x paddle-shaped. x about 2.7× as long (proximal-distally) as wide (latero-medially). x 13th post-ocular segment x x Appendage of 13th post-ocular segment (thoracopod 8) x x as bifid, but undivided structure. Future subdivision into coxa, basipod, endopod and exopod indicated, but no functional joints apparent. x vermicular, undifferentiated, hardly developed. with coxa, basipod, endopod and exopod. x Coxa x x tube-shaped to slightly conical (tapering distally). x tube-shaped. x about as long (proximal-distally) as wide (latero-medially). x with proximal margin about 1.4× as wide (latero-medially) as proximo-distal axis of the element. x Basipod x x tube-shaped. x x about 2.5× as long (proximal-distally) as wide (latero-medially). x about 6.2× as long (proximal-distally) as wide (latero-medially). x Inner branch (endopod?) x x as undifferentiated lobe. x about 3.8× as long (proximal-distally) as wide (latero-medially). x about 0.7× as long (proximal-distally) as outer branch. x as a single, elongate paddle. x about 5.8× as long (proximal-distally) as wide (latero-medially). x about 0.6× as long (proximal-distally) as outer branch. x Outer branch (exopod?) x x as undifferentiated lobe. x about 3.8× as long (proximal-distally) as wide (latero-medially). x bipartite. x Proximal element of outer branch x tube-shaped. x with distal margin drawn-out and slightly overlapping distal element of outer branch (in medial view). x about 6× as long (proximal-distally) as wide (latero-medially). x Distal element of outer branch x paddle-shaped. x about 2.8× as long (proximal-distally) as wide (latero-medially). x 14th post-ocular segment x x Appendage of 14th post-ocular segment (pleopod 1) x x with basipod carrying endopod and exopod. x x Basipod x x square-shaped in anterior view. x rectangular in anterior view. x about as long (proximo-distally) as wide (medio-laterally). x about 1.5× as long (proximo-distally) as wide (medio-laterally). x Endopod x x bipartite. x tripartite. x with clear, functional subdivision by straight lines. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x straight, paddle-shaped. x x Proximal part x x semi-elliptic in anterior view. x x distally with oblique transition line. x x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.3× as long maximum (proximo-distally) as maximum width of the element (medio-laterally). x with about 15 setae indicated by insertions on medial margin. x x with future subdivison in proximal and medial part indicated by slight notches on lateral and medial margin. x with almost straight medial margin. Medial part x x triangular in anterior view. x with lateral margin about 0.8× as long (proximo-distal axis of the endopod) as maximum width of the element (medio-lateral axis of the endopod). x not yet set of from proximal part. x Distal part x x semi-elliptic in anterior view. x x proximally with oblique transition line. x with transition to proximal part indicated by notch on medial and lateral margin. x about 1.2× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.3× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 25 setae indicated by insertions. x with about 30 setae indicated by insertions. x with medial margin bearing vermicular process proximally. x x with process about 0.2× as long (proximo-distally) as length of the distal part (proximo-distally). x with process about 0.3× as long (proximo-distally) as length of the distal part (proximo-distally). x Exopod x x bipartite. x x straight, paddle-shaped. x x with clear, functional subdivision. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x Proximal part x x subtriangular in anterior view, proximally truncated; carrying a gill antero-medially. x x distally with transition line perpendicular to main axis of the exopod. distally with slightly convex transition line. x x about 1.4× as long (proximo-distally) as maximum width of the element (medio-laterally). x x without indications of setae. x with about 3 setae indicated by insertions at the distal indentations. x with about 5 setae indicated by insertions on lateral margin. x Gill x x branched, with 3 branches. x with about 15 branches. x Distal part x x ovoid in anterior view. x x about as long (proximo-distally) as maximum width of the element (latero-medially). x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 40 setae indicated by insertions. x x 15th post-ocular segment x x Appendage of 15th post-ocular segment (pleopod 2) x x with basipod carrying endopod and exopod. x x Basipod x x sub-square shaped in anterior view. x rectangular in anterior view. x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.3× as long (proximo-distally) as wide (medio-laterally). x Endopod x x bipartite. x tripartite. x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x Proximal part x x semi-circular in anterior view. x semi-elliptic in anterior view. x distally with oblique transition line. x x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.2× as long (proximo-distally) as maximum width of the element (medio-laterally). x with future subdivison in proximal and medial part indicated by slight notches on lateral and medial margin. x with about 10 setae indicated by insertions on medial margin. x with about 20 setae indicated by insertions. x Medial part x x triangular in anterior view. x with lateral margin about 0.7× as long (proximo-distal axis of the endopod) as maximum width of the element (medio-lateral axis of the endopod). x not yet set of from proximal part. x Distal part x x semi-elliptic in anterior view. x x proximally with oblique transition line. x with transition line to proximal part indicated by notch on medial and lateral margin. x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x about as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 30 setae indicated by insertions. x with about 35 setae indicated by insertions. x with medial margin bearing vermicular process proximally. x x with process about 0.2× as long (proximo-distally) as length of the distal part (proximo-distally). x with process about 0.3× as long (proximo-distally) as length of the distal part (proximo-distally). x Exopod x x bipartite. x x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x Proximal part x x subtriangular in anterior view, proximally truncated; carrying a gill antero-medially. x x distally with slightly convex transition line. x x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.6× as long (proximo-distally) as maximum width of the element (medio-laterally). x without indications of setae. x x with 1 setae indicated by insertion on lateral margin and 2 on medial margin. x Gill x x branched, with 3 branches. x with about 30 branches. x Distal part x x subcircular in anterior view. x ovoid in anterior view. x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 30 setae indicated by insertions. with about 40 setae indicated by insertions. x x 16th post-ocular segment x x Appendage of 16th post-ocular segment (pleopod 3) x x with basipod carrying endopod and exopod. x x Basipod x x sub-square shaped in anterior view. x rectangular in anterior view. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.3× as long (proximo-distally) as wide (medio-laterally). x about 1.9× as long maximum (proximo-distally) as maximum width of the element (latero-medially). Endopod x x bipartite. x tripartite. x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x Proximal part x x semi-circular in anterior view. x semi-elliptic in anterior view. x distally with oblique transition line. x x with future subdivison in proximal and medial part indicated by slight notches on lateral and medial margin. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 15 setae indicated by insertions on medial margin. x x Medial part x x triangular in anterior view. x with lateral margin about 0.6× as long (proximo-distal axis of the endopod) as maximum width of the element (medio-lateral axis of the endopod). x not yet set of from proximal part. x Distal part x x semi-elliptic in anterior view. x x future subdivision (straight transition line proximally) indicated by notch on medial and lateral margin. x proximally with oblique transition line. x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x about as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 30 setae indicated by insertions. x with about 35 setae indicated by insertions. x with medial margin bearing vermicular process proximally. x x with process about 0.3× as long (proximo-distally) as length of the distal part (proximo-distally). x x Exopod x x bipartite. x x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x with subdivision in proximal and distal part indicated by notch on medial and lateral margin. x Proximal part x x subtriangular in anterior view, proximally truncated; carrying a gill antero-medially. x x distally with transition line almost perpendicular to main axis of the exopod. distally with slightly convex transition line. x x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.4× as long (proximo-distally) as maximum width of the element (medio-laterally). x without indications of setae. x with about 3 setae indicated by insertions on the lateral margin. x Gill x x branched, with 4 branches. x branched, with about 25 branches. x Distal part x x ovoid in anterior view. x subcircular in anterior view. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 40 setae indicated by insertions. x with about 55 setae indicated by insertions. x 17th post-ocular segment x x Appendage of 17th post-ocular segment (pleopod 4) x x with basipod carrying endopod and exopod. x x Basipod x x sub-square shaped in anterior view. x subrectangular in anterior view. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x Endopod x x bipartite. x tripartite. x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x x Proximal part x x semi-circular in anterior view. x x distally with oblique transition line. x x with future subdivison in proximal and medial part indicated by slight notches on lateral and medial margin. x about 0.7× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 20 setae indicated by insertion on medial margin. x with about 22 setae indicated by insertion on medial margin. x Medial part x x triangular in anterior view. x with lateral margin about 0.6× as long (proximo-distal axis of the endopod) as maximum width of the element (medio-lateral axis of the endopod). x not yet set of from proximal part. x Distal part x x semi-elliptic in anterior view. x x proximally with oblique transition line. x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x x with about 30 setae indicated by insertions. x with about 35 setae indicated by insertions. x with medial margin bearing vermicular process proximally. x x with process about 0.3× as long (proximo-distally) as length of the distal part (proximo-distally). x x Exopod x x bipartite. x x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x x Proximal part x x subtriangular in anterior view, proximally truncated; carrying a gill antero-medially. x x distally with slightly convex transition line. x x about 1.1× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.2× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 1 setae indicated by insertions on the lateral margin distally. x x Gill x x branched, with 4 branches. x branched, with about 25 branches. x Distal part x x ovoid in anterior view. x semi-circular in anterior view. x about 0.7× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 45 setae indicated by insertions. x with about 50 setae indicated by insertions. x 18th post-ocular segment x x Appendage of 18th post-ocular segment (pleopod 5) x x with basipod carrying endopod and exopod. x x Basipod x x trapezoidal in anterior view; with rounded corners. x x with convex distal edge. x x about 0.4× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.7× as long (proximo-distally) as maximum width of the element (medio-laterally). x distal edge about 1.3× as wide (medio-laterally) as proximal edge (medio-laterally). x distal edge about 1.4× as wide (medio-laterally) as proximal edge (medio-laterally). x Endopod x x bipartite. x tripartite. x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x x Proximal part x x semi-circular in anterior view. x x distally with transition line almost perpendicular to main axis of the endopod. x distally with oblique transition line. x with future subdivison in proximal and medial part indicated by slight notches on lateral and medial margin. x about 0.6× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.7× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 10 setae on medial margin. x with about 15 setae indicated by insertions on medial margin. x Medial part x x triangular in anterior view. x with lateral margin about 0.5× as long (proximo-distal axis of the endopod) as maximum width of the element (medio-lateral axis of the endopod). x not yet set of from proximal part. x Distal part x x semi-elliptic in anterior view. x x proximally with oblique transition line. x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.9× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 30 setae indicated by insertions. x with about 40 setae indicated by insertions. x with medial margin bearing vermicular process proximally. x x with process about 0.3× as long (proximo-distally) as length of the distal part (proximo-distally). x with process about 0.4× as long (proximo-distally) as length of the distal part (proximo-distally). x Exopod x x bipartite. x x straight, paddle-shaped. x x with clear, functional subdivision by transition line. x x Proximal part x x subtriangular in anterior view, proximally truncated; carrying a gill antero-medially. X X distally with slightly convex transition line. x x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 1.2× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 3 setae indicated by insertions on lateral margin distally. x with about 15 setae indicated by insertions on lateral margin. x Gill x x branched, with 4 branches. x branched, with about 30 branches. x Distal part x x subcircular in anterior view. x x about 0.7× as long (proximo-distally) as maximum width of the element (medio-laterally). x about 0.8× as long (proximo-distally) as maximum width of the element (medio-laterally). x with about 40 setae indicated by insertions. x with about 55 setae indicated by insertions. x 19th post-ocular segment x x Appendage of 19th post-ocular segment (uropod) x x with basipod carrying endopod and exopod. x x Basipod x x with basal part and drawn-out part. x x about 3.3× as long (proximo-distally) as maximum width (latero-medially) of the total element. x about 4.2× as long (proximo-distally) as maximum width (latero-medially) of the total element. x Basal part of basipod x x subrectangular in ventral view. x x with straight lateral margin and medial margin curved inwards. x x about 1.1× as long (proximo-distally) as maximum width (latero-medially). x about 1.2× as long (proximo-distally) as maximum width (latero-medially). x Drawn-out part of basipod x x with two distal spines of different length. x x with medial spine about 2.0× as long (proximo-distally) maximum width of the spine (medio-laterally). x with medial spine about 3.4× as long (proximo-distally) maximum width of the spine (medio-laterally). x with lateral spine about 2.6× as long (proximo-distally) maximum width of the spine (medio-laterally). x with lateral spine about 3.7× as long (proximo-distally) maximum width of the spine (medio-laterally). x with lateral spine about 3.3× as long (proximo-distally) as medial spine. x with lateral spine about 3.6× as long (proximo-distally) as medial spine. x Endopod x x subelliptic in ventral view. x x paddle-like shaped. x x about 2.4× as long (proximo-distally) as maximum width of the element (latero-medially). x about 2.8× as long (proximo-distally) as maximum width of the element (latero-medially). x with about 25 setae indicated by insertions. x with about 40 setae indicated by insertions. x Exopod x x bipartite. x not yet separated. x subrectangular in ventral view; with distal margin rounded. x with distal part of lateral margin bearing 2 spines (length increasing from proximal to distal). x with about 15 setae indicated by insertions. x Proximal part x truncated-cone shape like. x about 1.3× as long (proximo-distally) as distal margin of the element. x with distal part of lateral margin bearing 6 movable spines (length increasing from proximal to distal). x Distal part x subcircular in ventral view. x about 1.1× as long (proximo-distally) as maximum width (latero-medially) of the element. x with about 30 setae indicated by insertions. x Telson x x articulated at the 19th post-ocular segment. x x subelliptic in ventral/dorsal view. subcircular in ventral/dorsal view. x x with anus ventrally on proximal part of medial axis of the telson. x x without spines about 0.8× as long (proximal-distal axis) as maximum width of the element (latero-medial axis). x x with two large spines on each lateral margin and two larger spines on the distal margin. x x with first lateral spines (counted from proximal to distal) about 0.1× as long (proximo-distally) as main proximo-distal axis of the telson. x with first lateral spines (counted from proximal to distal) about 0.2× as long (proximo-distally) as main proximo-distal axis of the telson. x with second lateral spines (counted from proximal to distal) about 0.1× as long (proximo-distally) as main proximo-distal axis of the telson. x x with distally drawn-out spines about 0.4× as long (proximo-distally) as main proximo-distal axis of the telson. x x with distally drawn-out spines about 0.7× as long (proximo-distally) as the base of the drawn-out part. x x with gap between distally drawn-out spines (between tips) about 0.5× as wide (latero-medially) as maximum width of the telson (latero-medially). x with gap between distally drawn-out spines (between tips) about 0.6× as wide (latero-medially) as maximum width of the telson (latero-medially). x with about 36 small spines on the distal margin between the two distally drawn-out spines. x x with single small spine on each lateral margin at the base of the first and second lateral spine (from proximal to distal) distally. x x with single small spine on each lateral margin between the second lateral spine (from proximal to distal) and the distally drawn-out spines. x x .
DISCUSSION
In the investigated specimens, multiple morphological changes are documented throughout the observed ontogeny, some of which are more prominent than others. We focus on the drastic changes as well as those of interest in a wider evolutionary comparison, including also fossil mantis shrimps.
Sensorial apparatus
The differentiation of the sensory appendages, antennula and antenna, is well accessible. In the antennula only one flagellum is fully developed in stage 1 (Fig. 5B); all flagella are subdivided into numerous annuli in stage 3 (Fig. 8B).
The antennula is only known for a single fossil mantis shrimp larva (Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.), and remains difficult to interpret (cf. Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.; 2015Haug, C.; Wiethase, J. H. and Haug, J. T. 2015. New records of Mesozoic mantis shrimp larvae and their implications on modern larval traits in stomatopods. Palaeodiversity, 8: 121-133.). The pattern of flagella development in the investigated larvae seems to be similar to what is observable in the fossil: the annulation of the dorsal flagellum is more pronounced, and the further “ventral” flagellum appears to be shorter.
The antenna is preserved in the same fossil specimen. In congruence to the described pattern it appears to be subdivided into annuli only distally while the proximal region appears to be not (yet) subdivided (Haug et al., 2008Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.).
Feeding apparatus
The mouth parts barely change over the developmental stages (Fig. 5E-IvsFig. 8E-H) and appear similar to mouth parts of adults of other mantis shrimp species (Haug et al., 2012aHaug, C.; Sallam, W.S.; Maas, A.; Waloszek, D.; Kutschera, V. and Haug, J. T. 2012a. Tagmatization in Stomatopoda - reconsidering functional units of modern-day mantis shrimps (Verunipeltata, Hoplocarida) and implications for the interpretation of fossils. Frontiers in Zoology, 9: 31, 14p. ) apart from the hypostome-labrum complex. This does not yet show the extreme elongation seen in adults (Haug et al., 2012aHaug, C.; Sallam, W.S.; Maas, A.; Waloszek, D.; Kutschera, V. and Haug, J. T. 2012a. Tagmatization in Stomatopoda - reconsidering functional units of modern-day mantis shrimps (Verunipeltata, Hoplocarida) and implications for the interpretation of fossils. Frontiers in Zoology, 9: 31, 14p. ). This similarity between larval and adult mouth parts indicates that a fully functional mouth-part apparatus is developed early in ontogeny.
The “cleaning appendage”, maxilliped 1, remains almost unchanged between the two investigated larval stages (Fig. 6AvsFig. 9A), indicating that its use does not differ early in ontogeny, i.e., its function for cleaning the eyes is established early within the ontogenetic sequence.
The shape of the large raptorial appendage, maxilliped 2, is also almost unchanged (Fig. 6BvsFig. 9B). Based on the size and shape in stage 1, its primary function, catching prey, is presumably established early within the ontogenetic sequence. Its armature resembles that of many known larvae. It lacks the differentiation of most adults identifying them as representatives of the two standard ecotype categories, smasher and spearer. This means it does neither possess a swelling on the dactylus (as it is characteristic for smasher) nor does it have the elongate spines of the dactylus (which is a spearer feature) (Giesbrecht, 1910Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p. ; Caldwell and Dingle, 1975Caldwell, R.L. and Dingle, H. 1975. Ecology and evolution of agonistic behavior in stomatopods. Naturwissenschaften, 62: 214-222. ; Kunze, 1981Kunze, J.C. 1981. The functional morphology of stomatopod Crustacea. Philosophical Transactions of the Royal Society B: Biological Sciences, 292: 255-328.; Weaver et al., 2012Weaver, J.C.; Milliron, G.M.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E. and Kisailus, D. 2012. The stomatopod dactyl club: a formidable damage-tolerant biological hammer. Science, 336: 1275-1280. ). The morphology seen in the larvae resembles that of adult fossil representatives of the mantis shrimps lineages, i.e., Jurassic mantis shrimps of species of Sculda (Fig. 12A2; Haug et al., 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.). Following available phylogenetic reconstructions (Schram, 2007Schram, F. R. 2007. Paleozoic proto-mantis shrimp revisited. Journal of Paleontology, 81: 895-916.; Haug et al., 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.), the condition in Sculda is plesiomorphic, while the one in modern forms is apomorphic. With this polarity, modern larvae retain a plesiomorphic condition, and only modern adults show the apomorphic state. This pattern indicates that the morphology of the adults of the modern species evolved through peramorphic heterochrony, more precisely hypermorphosis (Fig. 12A; Klingenberg, 1998Klingenberg, C.P. 1998. Heterochrony and allometry: the analysis of evolutionary change in ontogeny. Biological Reviews of the Cambridge Philosophical Society, 73: 79-123.; McKinney and McNamara, 1991McKinney, M.L. and McNamara, K.J. 1991. Heterochrony. p. 1-12. In: M. McKinney and K.J. McNamara (eds), Heterochrony: The Evolution of Ontogeny. New York, London, Plenum Press.; Smith, 2001Smith, K.K. 2001. Heterochrony revisited: the evolution of developmental sequences. Biological Journal of the Linnean Society, 73: 169-186.; Webster and Zelditch, 2005Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.).

Reconstructions of elements 1 & 2 of maxilliped 2 (A) and pleopods (B) of different Mesozoic and extant malacostracans. Appearance of heterochronic events highlighted. Upper: adult condition. Lower: early ontogenetic condition. A1: Tyrannophontes (from Haug and Haug, 2011Haug, C. and Haug, J.T. 2011. Die Evolution der Fangschreckenkrebse (Stomatopoda, Malacostraca) im Lichte neuer mesozoischer Fossilfunde. Archaeopteryx, 29: 13-19.), A2: Sculda sp. (from Haug et al., 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.), A3: extant mantis shrimp (upper from Caldwell and Dingle, 1975Caldwell, R.L. and Dingle, H. 1975. Ecology and evolution of agonistic behavior in stomatopods. Naturwissenschaften, 62: 214-222. , smasher and spearer; lower as observed by the authors), B1: Leptostraca: Nebalia sp. (from Martin et al., 1996Martin, J.W.; Vetter, E.W. and Cash-Clark, C.E. 1996. Description, external morphology, and natural history observations of Nebalia hessleri, new species (Phyllocarida: Leptostraca), from southern California, with a key to the extant families and genera of the Leptostraca. Journal of Crustacean Biology, 16: 347-372.; Olesen and Walossek, 2000Olesen, J. and Walossek, D. 2000. Limb ontogeny and trunk segmentation in Nebalia species (Crustacea, Malacostraca, Leptostraca). Zoomorphology, 120: 47-64.), B2: Stomatopoda: Verunipeltata (as observed by the authors), B3: Decapoda (from Rötzer and Haug, 2015Rötzer, M.A.I.N. and Haug, J.T. 2015. Larval development of the European lobster and how small heterochronic shifts lead to a more pronounced metamorphosis. International Journal of Zoology, 2015: 345172, 17p. ; lower as observed), B4: Mysidacea (from Wittmann, 1986Wittmann, K.J. 1986. Untersuchungen zur Lebensweise und Systematik von Leptomysis truncata und zwei verwandten Formen (Crustacea, Mysidacea). Annalen des Naturhistorischen Museums in Wien, Serie B für Botanik und Zoologie, 87: 295-323.). Note the hypermorphotic events resulting in the short pleopodal basipod and slender 2nd element of maxilliped 2 in adult stomatopods.
Maxillipeds 3-5 are rather underdeveloped in stage 1 (Fig. 6C-E) and most likely not yet (fully) functional. In stage 3 they are significantly better developed (Fig. 9C-E). A significant size increase, especially of the propodus and the dactylus, indicates that these appendages have become fully functional at this stage. Hence further anterior appendages of the feeding apparatus are developed earlier than the further posterior ones. This pattern could reflect the ancestral developmental pattern of Eucrustacea, developing further anterior appendages before the more posterior ones (e.g., Walossek, 1993Walossek, D. 1993. The upper Cambrian Rehbachiella and the phylogeny of Branchiopoda and Crustacea. Lethaia, 26: 318-318.).
Walking appendages
The walking appendages develop drastically, as they are barely developed in stage 1 (Fig. 7A-C), but well-differentiated in stage 3 (Fig. 10A-C). They are, however, still comparably small.
The identity of the branches remains challenging. Among other crustaceans, the walking branch is the inner branch (endopod); in mantis shrimps it is the topologically outer branch (Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.). Literature suggests that this is due to a rotation of early limb buds (e.g., Schram, 1986Schram, F.R. 1986. Crustacea. Oxford, Oxford University Press , 606p.); consequently, the outer branch would represent the endopod, while the inner would be the exopod in mantis shrimps.
We can consider the morphology of the branches without this pre-assumption. The basic morphology of the outer branch has in principle reached adult condition (cf. e.g., Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.) in stage 3, and appears only stouter to a certain degree. It is bipartite, the distal part forming a paddle (Fig. 10A-C), a morphology that could be assumed for an exopod, but would in fact be unusual to a certain degree for an endopod, at least more so than for an exopod. Still in fact also the inner is rather elongated to paddle-shaped (Fig. 10A-C). Hence the basic shape gives no useful hint here. Yet, the insertion area of the outer branch is more informative. The basipod area, where the outer branch arises, is sloped (Fig. 10A-C); this is a typical arrangement for the insertion area of the exopod (Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.; Haug et al., 2013bHaug, J.T.; Maas, A.; Haug, C. and Waloszek, D. 2013b. Chapter 2. Evolution of crustacean appendages. p. 34-73. In: L. Watling and M. Thiel (eds), Functional Morphology and Diversity. The Natural History of the Crustacea, Vol. 1. Oxford, Oxford University Press .).
Walking appendages are only preserved in few fossils, and only in non-larval forms (Haug et al., 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.). These are additionally restricted to the more distal elements and thus do not contribute to the discussion of the identity of the branches. Still, in the fossils the distal element is not paddle-shaped (Haug et al., 2010bHaug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.).
The data do not allow to make a definite decision on the identity of the branches. Yet, they suggest that there was not necessarily a rotation of limb buds in the walking limbs, and mantis shrimps might indeed walk on their exopods.
Pleopods
The principle development of the pleopods is characterised by a significant shape change of the basipod, as well as the development of the gills (Fig. 7D-HvsFig. 10D-H). The basipod shape is generally more elongate in proximal-distal axis than in medial-lateral axis in stage 1 (Fig. 7D-H). In stage 3 the basipods appear stouter; the proximal-distal axis is either as long as the medial-lateral axis or shorter (Fig. 10D-H). The adult condition is a rather short, more broad than long (proximo-distally) (Giesbrecht, 1910Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p. ), continuing the trend of development between stage 1 and stage 3.
Caridoidan crustaceans (Caridoida and Hoplocarida (incl. Stomatopoda) are sister groups, together forming Eumalacostraca; Richter and Scholtz, 2001Richter, S. and Scholtz, G. 2001. Phylogenetic analysis of the Malacostraca (Crustacea). Journal of Zoological Systematics and Evolutionary Research, 39: 113-136. ) appear to retain the more elongate shape of the pleopodal basipod during life (Fig. 12B3, B4; e.g., Wittmann, 1986Wittmann, K.J. 1986. Untersuchungen zur Lebensweise und Systematik von Leptomysis truncata und zwei verwandten Formen (Crustacea, Mysidacea). Annalen des Naturhistorischen Museums in Wien, Serie B für Botanik und Zoologie, 87: 295-323.; Davie, 2002Davie, P.J. 2002. Crustacea: Malacostraca: Phyllocarida, Hoplocarida, Eucarida (Part 1). In: A. Wells and W.W.K. Houston (eds), Zoological Catalogue of Australia. Vol. 19.3A. Melbourne, ABRS & CSIRO Publishing, 551p.; Rötzer and Haug, 2015Rötzer, M.A.I.N. and Haug, J.T. 2015. Larval development of the European lobster and how small heterochronic shifts lead to a more pronounced metamorphosis. International Journal of Zoology, 2015: 345172, 17p. ). This is also true for the developmental pattern of representatives of the phyllocaridan Nebalia (Fig. 12B1; Martin et al., 1996Martin, J.W.; Vetter, E.W. and Cash-Clark, C.E. 1996. Description, external morphology, and natural history observations of Nebalia hessleri, new species (Phyllocarida: Leptostraca), from southern California, with a key to the extant families and genera of the Leptostraca. Journal of Crustacean Biology, 16: 347-372.; Olesen and Walossek, 2000Olesen, J. and Walossek, D. 2000. Limb ontogeny and trunk segmentation in Nebalia species (Crustacea, Malacostraca, Leptostraca). Zoomorphology, 120: 47-64.; Phyllocarida (including Leptostraca with only extant representatives) is sister group to Eumalacostraca; Richter and Scholtz, 2001Richter, S. and Scholtz, G. 2001. Phylogenetic analysis of the Malacostraca (Crustacea). Journal of Zoological Systematics and Evolutionary Research, 39: 113-136. ). Although the latter do not pass through distinct larval stages, this upright rectangular shape (in anterior or posterior view), retained throughout ontogeny, may represent the ancestral condition for Malacostraca. Within Stomatopoda, at least Verunipeltata (the modern forms; still unclear for the earlier off-shoots of this evolutionary lineage) have evolved the described diverging pattern, starting with elongate basipods in early stages and becoming more stout successively. The early larval stages, in this case, appear to recapitulate the ancestral morphology of Malacostraca. Adults deviate from this morphology. This morphology could therefore be recognized as an evolutionary shift of developmental timing, or heterochronic shift, more precisely a peramorphosis (leading to a more “adultised” appearance).
There are three sub-types of peramorphosis, which can be identified in a global heterochrony, i.e., when the whole organism is affected (Webster and Zelditch, 2005Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.). Since the described phenomenon is local, affecting just a specific structure, it is more difficult to assign the exact subtype of peramorphosis. In the present case, a new developmental state is most likely added “on top” of the original state. This should therefore be considered to be a case hypermorphosis (Fig. 12B). As the new state is not added at the end of ontogeny (as it is not a global shift), but within the sequence, it should be better described as hypermorphosis+predisplacement (see Haug et al., 2010aHaug, J.T.; Maas, A. and Waloszek, D. 2010a. †Henningsmoenicaris scutula, †Sandtorpia vestrogothiensis gen. et sp. nov. and heterochronic events in early crustacean evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 100: 311-350., c) to underline this difference.
The gills are barely developed in stage 1 (Fig. 7D-H), but become more pronounced in stage 3 (Fig. 10D-H). They are significantly less well developed than in the adult (Giesbrecht, 1910Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p. ). The gills are presumably not yet fully functional in the larval stages. This is most likely compensated by the relatively large leaf-shaped epipods on the cleaning appendage and the raptorial appendages (see also Maas et al., 2009Maas, A.; Haug, C.; Haug, J.T.; Olesen, J.; Zhang, X. and Waloszek, D. 2009. Early crustacean evolution and the appearance of epipodites and gills. Arthropod Systematics & Phylogeny, 67: 255-273. ). These are significantly smaller in relation to the main parts of the appendages in the adults (Giesbrecht, 1910Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p. ; Maas et al., 2009Maas, A.; Haug, C.; Haug, J.T.; Olesen, J.; Zhang, X. and Waloszek, D. 2009. Early crustacean evolution and the appearance of epipodites and gills. Arthropod Systematics & Phylogeny, 67: 255-273. ). Thus, the gas exchange is most likely shifted from the thorax in larval stages to the pleon in the adult. In other malacostracans, the gas exchange is maintained in the thoracic region throughout ontogeny (Richter and Scholtz, 2001Richter, S. and Scholtz, G. 2001. Phylogenetic analysis of the Malacostraca (Crustacea). Journal of Zoological Systematics and Evolutionary Research, 39: 113-136. ; Boxshall and Jaume, 2009Boxshall, G.A. and Jaume, D. 2009. Exopodites, epipodites and gills in crustaceans.Arthropod Systematics & Phylogeny, 67: 229-254.). It is therefore likely that the ontogenetic shift in mantis shrimps does not represent a plesiomorphy (suggested in Maas et al., 2009Maas, A.; Haug, C.; Haug, J.T.; Olesen, J.; Zhang, X. and Waloszek, D. 2009. Early crustacean evolution and the appearance of epipodites and gills. Arthropod Systematics & Phylogeny, 67: 255-273. ) but represents an apomorphic heterochronic shift. This would also represent a case of hypermorphosis+predisplacement.
This interpretation is complicated by the general developmental pattern of thoracopods and pleopods. Plesiomorphically, anterior appendages are further developed than more posterior ones (Walossek, 1993Walossek, D. 1993. The upper Cambrian Rehbachiella and the phylogeny of Branchiopoda and Crustacea. Lethaia, 26: 318-318.). Mantis shrimps deviate from this pattern to a certain extent, as the pleopods develop quite early while appendages of the thorax develop later. The timing of development of pleopods and thoracopods has therefore been affected by developmental shifts. A clear identification of parts affected by peramorphosis (by early appearance, accelerated or prolongated growth, Webster and Zelditch, 2005Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.) or by paedomorphosis (by late appearance, retarded or abbreviated growth, Webster and Zelditch, 2005Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.) demands for a detailed comparison, i.e. more data from more larval sequences.
Tailfan
The uropods change most drastically at the exopod, but in general increase significantly in size (Fig. 7IvsFig. 10I). The exopod is undivided in stage 1 and equipped with only three immovable teeth (Fig. 7I). In stage 3 the distal paddle is jointed against the proximal part. There are more lateral teeth (six) and the most proximal ones appear to be jointed (Fig. 10I). The uropod has principally reached a typical “post-larval” state in this late larval stage (Hamano and Matsuura, 1987Hamano, T. and Matsuura, S. 1987. Egg size, duration of incubation, and larval development of the Japanese mantis shrimp in the laboratory. Nippon Suisan Gakkaishi, 53: 23-29.; Morgan and Goy, 1987Morgan, S.G. and Goy, J.W. 1987. Reproduction and larval development of the mantis shrimp Gonodactylus bredini (Crustacea: Stomatopoda) maintained in the laboratory. Journal of Crustacean Biology, 7: 595-618.).
This type of development is different from representatives of the earlier evolutionary lineage towards Verunipeltata. The Jurassic mantis shrimp Spinosculda ehrlichi has an undivided uropod throughout its ontogeny (Haug et al., 2009bHaug, C.; Haug, J.T. and Waloszek, D. 2009b. Morphology and ontogeny of the Upper Jurassic mantis shrimp Spinosculda ehrlichi n. gen. n. sp. from southern Germany. Palaeodiversity, 2: 111-118.); the subdivision of the uropodal exopod is an autapomorphy of Verunipeltata (Haug et al., 2013cHaug, C.; Kutschera, V.; Ahyong, S.T.; Vega, F.J.; Maas, A.; Waloszek, D. and Haug, J.T. 2013c. Re-evaluation of the Mesozoic mantis shrimp Ursquilla yehoachi based on new material and the virtual peel technique. Palaeontologia Electronica, 16: 16.2.5T. ). Yet, the distinction of larval versus non-larval exopod can be made based on its shape and armature. The larvae often possess more lanceolate-like endopods and exopods, and the non-larval stages paddle-shaped ones (Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.). The larval stages appear to lack armature (at least the earlier ones) (Fig. 7I; Manning and Provenzano, 1963Manning, R.B. and Provenzano Jr., A.J. 1963. Studies on development of stomatopod Crustacea I. Early larval stages of Gonodactylus oerstedii Hansen. Bulletin of Marine Science of the Gulf and Caribbean, 13: 467-487.; Pyne, 1972Pyne, R.R. 1972. Larval development and behaviour of the mantis shrimp, Squilla armata Milne Edwards (Crustacea: Stomatopoda). Journal of Royal Society of New Zealand, 2: 121-146.; Morgan and Provenzano, 1979Morgan, S.G. and Provenzano, A.J. 1979. Development of pelagic larvae and postlarva of Squilla empusa (Crustacea, Stomatopoda), with an assessment of larval characters within the Squillidae. Fishery Bulletin, 77: 61-90.; Hamano and Matsuura, 1987Hamano, T. and Matsuura, S. 1987. Egg size, duration of incubation, and larval development of the Japanese mantis shrimp in the laboratory. Nippon Suisan Gakkaishi, 53: 23-29.), while the exopods of the non-larval stages possess movable teeth (Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.). An exopod possessing armature and being paddle-shaped already in the late larval stages, as in the stage 3 larvae, could result from a heterochronic shift, a predisplacement. This shift could characterize Verunipeltata or an in-group of it as it is also known in other verunipeltatan mantis shrimps (e.g., Alikunhi, 1944Alikunhi, K.H. 1944. Final pelagic larva of Squilla hieroglyphica Kemp. Current Science, 13: 237-238.; Diaz, 1998Diaz, G.A. 1998. Description of the last seven pelagic larval stages of Squilla sp. (Crustacea, Stomatopoda). Bulletin of Marine Science, 62: 753-762.). This conclusion must remain an assumption until larval stages of more fossil and extant representatives have been investigated for this particular feature.
The spination of the telson changes gradually from stage 1 to stage 3 (Fig. 11AvsFig. 11B). All spines present in stage 3 appear to have been developed already in stage 1, and become more pronounced in stage 3 (Fig. 11). The telson is one of the highly variable features among different extant mantis shrimp species (Ahyong, 1997Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.) and many of its structures are difficult to homologize with those of the earlier representatives of the lineage. A detailed comparison demands more data especially on the extant larval development of this structure.
CONCLUSIONS AND OUTLOOK
Conclusions that can be made with the here presented data are, of course, limited, as only larvae of a single species were studied. Nevertheless, the data already strongly indicate heterochronic events in the evolution of mantis shrimps. To further corroborate this, more data from larval sequences (fossil and extant), documented with the method used in this study, are needed.
ACKNOWLEDGEMENTS
We thank Nils Brenke, Wilhelmshaven, and Björn von Reumont, formerly Bonn, now Gießen, for kindly providing us with specimens of mantis shrimp larvae; Stefan Liebau, formerly Ulm, now Tübingen, for help with microscopy; J. Matthias Starck, Munich, for his support. Christopher Tudge and two anonymous reviewers kindly provided helpful comments on the manuscript. This project was part of JTH's project 'Palaeo-Evo-Devo of Malacostraca' funded by the German Research Foundation (DFG) under HA-6300/3-1. CH was funded via the LMUexcellent Junior Researcher Fund. We also thank all people providing free or low cost software such as OpenOffice, CombineZM/ZP, Microsoft Image Composite Editor.
REFERENCES
- Ahyong, S.T. 1997. Phylogenetic analysis of the Stomatopoda (Malacostraca). Journal of Crustacean Biology, 17: 695-715.
- Ahyong, S.T. and Harling, C. 2000. The phylogeny of the stomatopod Crustacea.Australian Journal of Zoology, 48: 607-642.
- Ahyong, S.T.; Haug, J.T. and Haug, C. 2014. Stomatopoda. p. 185-189. In: J.W. Martin; J. Olesen and J.T. Høeg (eds), Atlas of Crustacean Larvae. Baltimore, The Johns Hopkins University Press.
- Alikunhi, K.H. 1944. Final pelagic larva of Squilla hieroglyphica Kemp. Current Science, 13: 237-238.
- Ax, P. 1995. Das System der Metazoa: ein Lehrbuch der phylogenetischen Systematik 1. Stuttgart, Gustav Fischer, 226p.
- Boxshall, G.A. and Jaume, D. 2009. Exopodites, epipodites and gills in crustaceans.Arthropod Systematics & Phylogeny, 67: 229-254.
- Caldwell, R.L. and Dingle, H. 1975. Ecology and evolution of agonistic behavior in stomatopods. Naturwissenschaften, 62: 214-222.
- Davie, P.J. 2002. Crustacea: Malacostraca: Phyllocarida, Hoplocarida, Eucarida (Part 1). In: A. Wells and W.W.K. Houston (eds), Zoological Catalogue of Australia. Vol. 19.3A. Melbourne, ABRS & CSIRO Publishing, 551p.
- Diaz, G.A. 1998. Description of the last seven pelagic larval stages of Squilla sp. (Crustacea, Stomatopoda). Bulletin of Marine Science, 62: 753-762.
- Eiler, S.M.; Haug, C. and Haug, J.T. 2016. Detailed description of a giant polychelidan Eryoneicus-type larva with modern imaging techniques. Spixiana, 39: 39-60.
- Giesbrecht, W. 1910. Stomatopoden, Erster Theil. Fauna und Flora des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Vol. 33. Berlin, R. Friedländer & Sohn, 239p.
- Hamano, T. and Matsuura, S. 1987. Egg size, duration of incubation, and larval development of the Japanese mantis shrimp in the laboratory. Nippon Suisan Gakkaishi, 53: 23-29.
- Haug, C. and Haug, J.T. 2011. Die Evolution der Fangschreckenkrebse (Stomatopoda, Malacostraca) im Lichte neuer mesozoischer Fossilfunde. Archaeopteryx, 29: 13-19.
- Haug, C.; Haug, J.T.; Waloszek, D.; Maas, A.; Frattigiani, R. and Liebau, S. 2009a. New methods to document fossils from lithographic limestones of southern Germany and Lebanon. Palaeontologia Electronica, 12: 12p.
- Haug, C.; Haug, J.T. and Waloszek, D. 2009b. Morphology and ontogeny of the Upper Jurassic mantis shrimp Spinosculda ehrlichi n. gen. n. sp. from southern Germany. Palaeodiversity, 2: 111-118.
- Haug, C.; Sallam, W.S.; Maas, A.; Waloszek, D.; Kutschera, V. and Haug, J. T. 2012a. Tagmatization in Stomatopoda - reconsidering functional units of modern-day mantis shrimps (Verunipeltata, Hoplocarida) and implications for the interpretation of fossils. Frontiers in Zoology, 9: 31, 14p.
- Haug, C.; Shannon, K. R.; Nyborg, T. and Vega, F. J. 2013a. Isolated mantis shrimp dactyli from the Pliocene of North Carolina and their bearing on the history of Stomatopoda. Bolétin de la Sociedad Geológica Mexicana, 65: 273-284.
- Haug, C.; Kutschera, V.; Ahyong, S.T.; Vega, F.J.; Maas, A.; Waloszek, D. and Haug, J.T. 2013c. Re-evaluation of the Mesozoic mantis shrimp Ursquilla yehoachi based on new material and the virtual peel technique. Palaeontologia Electronica, 16: 16.2.5T.
- Haug, C.; Wiethase, J. H. and Haug, J. T. 2015. New records of Mesozoic mantis shrimp larvae and their implications on modern larval traits in stomatopods. Palaeodiversity, 8: 121-133.
- Haug, J. T. In press. Chapter 9. Metamorphosis in Crustaceans: Towards a Synthesis. p. X-X. In: K. Anger, S. Harzsch and M. Thiel (eds), Developmental Biology and Larval Ecology. The Natural History of the Crustacea, Vol. 7. Oxford, Oxford University Press.
- Haug, J.T.; Haug, C. and Ehrlich, M. 2008. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity, 1: 103-109.
- Haug, J.T.; Maas, A. and Waloszek, D. 2010a. †Henningsmoenicaris scutula, †Sandtorpia vestrogothiensis gen. et sp. nov. and heterochronic events in early crustacean evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 100: 311-350.
- Haug, J.T.; Haug, C.; Maas, A.; Kutschera, V. and Waloszek, D. 2010b. Evolution of mantis shrimps (Stomatopoda, Malacostraca) in the light of new Mesozoic fossils. BMC Evolutionary Biology, 10: 290, 17p.
- Haug, J.T.; Waloszek, D.; Haug, C. and Maas, A. 2010c. High-level phylogenetic analysis using developmental sequences: The Cambrian †Martinssonia elongata, †Musacaris gerdgeyeri gen. et sp. nov. and their position in early crustacean evolution. Arthropod Structure & Development, 39: 154-173.
- Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K. and Waloszek, D. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. Journal of Microscopy, 244: 259-272.
- Haug, J.T.; Briggs, D.E.G. and Haug, C. 2012b. Morphology and function in the Cambrian Burgess Shale megacheiran arthropod Leanchoilia superlata and the application of a descriptive matrix. BMC Evolutionary Biology, 12: 162, 20p.
- Haug, J.T.; Maas, A.; Haug, C. and Waloszek, D. 2013b. Chapter 2. Evolution of crustacean appendages. p. 34-73. In: L. Watling and M. Thiel (eds), Functional Morphology and Diversity. The Natural History of the Crustacea, Vol. 1. Oxford, Oxford University Press .
- Hennig, W. 1966. Phylogenetic Systematics. Urbana, Univ. Illinois Press.
- Jirikowski, G.J.; Wolff, C. and Richter, S. 2015. Evolution of eumalacostracan development - new insights into loss and reacquisition of larval stages revealed by heterochrony analysis. EvoDevo, 6: 4.
- Klingenberg, C.P. 1998. Heterochrony and allometry: the analysis of evolutionary change in ontogeny. Biological Reviews of the Cambridge Philosophical Society, 73: 79-123.
- Kunze, J.C. 1981. The functional morphology of stomatopod Crustacea. Philosophical Transactions of the Royal Society B: Biological Sciences, 292: 255-328.
- Maas, A.; Haug, C.; Haug, J.T.; Olesen, J.; Zhang, X. and Waloszek, D. 2009. Early crustacean evolution and the appearance of epipodites and gills. Arthropod Systematics & Phylogeny, 67: 255-273.
- Manning, R.B. and Provenzano Jr., A.J. 1963. Studies on development of stomatopod Crustacea I. Early larval stages of Gonodactylus oerstedii Hansen. Bulletin of Marine Science of the Gulf and Caribbean, 13: 467-487.
- Marshall, N.J.; Land, M.F.; King, C.A. and Cronin, T.W. 1991. The compound eyes of mantis shrimps (Crustacea, Hoplocarida, Stomatopoda). II. Colour pigments in the eyes of stomatopod crustaceans: polychromatic vision by serial and lateral filtering. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 334: 57-84.
- Martin, J.W.; Vetter, E.W. and Cash-Clark, C.E. 1996. Description, external morphology, and natural history observations of Nebalia hessleri, new species (Phyllocarida: Leptostraca), from southern California, with a key to the extant families and genera of the Leptostraca Journal of Crustacean Biology, 16: 347-372.
- McKinney, M.L. and McNamara, K.J. 1991. Heterochrony. p. 1-12. In: M. McKinney and K.J. McNamara (eds), Heterochrony: The Evolution of Ontogeny. New York, London, Plenum Press.
- Morgan, S.G. and Goy, J.W. 1987. Reproduction and larval development of the mantis shrimp Gonodactylus bredini (Crustacea: Stomatopoda) maintained in the laboratory. Journal of Crustacean Biology, 7: 595-618.
- Morgan, S.G. and Provenzano, A.J. 1979. Development of pelagic larvae and postlarva of Squilla empusa (Crustacea, Stomatopoda), with an assessment of larval characters within the Squillidae. Fishery Bulletin, 77: 61-90.
- Olesen, J. and Walossek, D. 2000. Limb ontogeny and trunk segmentation in Nebalia species (Crustacea, Malacostraca, Leptostraca). Zoomorphology, 120: 47-64.
- Pyne, R.R. 1972. Larval development and behaviour of the mantis shrimp, Squilla armata Milne Edwards (Crustacea: Stomatopoda). Journal of Royal Society of New Zealand, 2: 121-146.
- Reif, W. E. 2002. Evolution of organ systems: phylogeny, function and reciprocal illumination. Senckenbergiana lethaea, 82: 356-366.
- Reif, W. E. 2005. Problematic issues of cladistics: 9. Hennig’s “Phylogenetic Systematics”. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 235: 289-342.
- Richter, S. and Scholtz, G. 2001. Phylogenetic analysis of the Malacostraca (Crustacea). Journal of Zoological Systematics and Evolutionary Research, 39: 113-136.
- Rötzer, M.A.I.N. and Haug, J.T. 2015. Larval development of the European lobster and how small heterochronic shifts lead to a more pronounced metamorphosis. International Journal of Zoology, 2015: 345172, 17p.
- Schram, F.R. 1986. Crustacea. Oxford, Oxford University Press , 606p.
- Schram, F. R. 2007. Paleozoic proto-mantis shrimp revisited. Journal of Paleontology, 81: 895-916.
- Smith, K.K. 2001. Heterochrony revisited: the evolution of developmental sequences. Biological Journal of the Linnean Society, 73: 169-186.
- Walossek, D. 1993. The upper Cambrian Rehbachiella and the phylogeny of Branchiopoda and Crustacea. Lethaia, 26: 318-318.
- Weaver, J.C.; Milliron, G.M.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E. and Kisailus, D. 2012. The stomatopod dactyl club: a formidable damage-tolerant biological hammer. Science, 336: 1275-1280.
- Webster, M. and Zelditch, M.L. 2005. Evolutionary modifications of ontogeny: heterochrony and beyond. Palaeobiology, 31: 354-372.
- Wittmann, K.J. 1986. Untersuchungen zur Lebensweise und Systematik von Leptomysis truncata und zwei verwandten Formen (Crustacea, Mysidacea). Annalen des Naturhistorischen Museums in Wien, Serie B für Botanik und Zoologie, 87: 295-323.
Appendix
Publication Dates
-
Publication in this collection
14 Feb 2020 -
Date of issue
2020
History
-
Received
08 Apr 2019 -
Accepted
22 Oct 2019