ABSTRACT
Although there are evidences that the proper supply of mineral nutrients to plants relieves water stress, little is known on the approach of how the drought affects the absorption and accumulation of nutrients by distinct sugar cane genotypes, or in different parts of a same plant. Thus, the objective of this study was to determine the content and accumulation of N, P and K in the aerial part of plant from three genotypes of sugar cane, submitted to three water potentials in the soil, and check the relationship of these variables with the tolerance of plants to prolonged drought. In order to access this objective, an experiment under greenhouse conditions, comprised by a fatorial 3 × 3, in a randomized block design, and four replicates was carried out. After 90 days from treatment imposition, the plant transpiration rate, plant dry mass, concentration of N, P and K were determined in leaves and culms, as well as in total plant shoot were measured. It was found that tolerance to drought in sugar cane is related to higher levels of N and K in the leaves and stems, and larger accumulations of K and P in the plant shoot. There is high positive correlations among accumulation of N, P and K in the plant shoot and dry matter production by plants submitted to drought. There are intermediate correlations among plant transpiration and nutrient uptake by plants under drought. Plants of the genotype SP81-3250 are more tolerante to prolonged drought, than the RB855453 and IACSP95-5000 plants.
Index terms:
Saccharum officinarum L.; water limitations; mineral nutrition; soil water potentials; plant growth
RESUMO
Embora existam evidências de que o adequado fornecimento de nutrientes minerais para plantas alivia o estresse hídrico, pouco se sabe sobre a abordagem de como a seca afeta a absorção e acumulação de nutrientes por distintos genótipos de cana-de-açúcar, ou por diferentes partes de uma mesma planta. Assim, o objetivo deste estudo foi determinar o teor e acúmulo de N, P e K na parte aérea da planta de três genótipos de cana-de-açúcar, submetidos a três potenciais hídricos do solo e verificar se existe relação dessas variáveis com a tolerância das plantas ao déficit hídrico prolongado. Para isso, foi realizado um experimento em casa de vegetação, em um esquema fatorial 3×3, em delineamento de blocos casualizados, com quatro repetições. Após 90 dias da imposição dos tratamentos, foi determinada a massa seca e a taxa de transpiração das plantas, a concentração de N, P e K nas folhas e colmos, além do acúmulo desses nutrientes na parte aérea das plantas. Verificou-se que a tolerância à seca em cana está relacionada com níveis mais altos de N e K nas folhas e caules e maiores acumulações de K e P na parte aérea das plantas. Existem fortes correlações positivas entre acúmulo de N, P e K na parte aérea das plantas e produção de matéria seca. Existem correlações intermediárias entre a transpiração da planta e absorção de nutrientes pelas plantas sob seca. Plantas do genótipo SP81-3250 são mais tolerantes à seca prolongada, que as plantas RB855453 e IACSP95-5000.
Termos para indexação:
Saccharum officinarum L.; limitação hídrica; nutrição mineral; potenciais hídricos do solo; crescimento da planta
INTRODUCTION
Due to the increase in global warming, the climate changes and the risk of extreme events, such as prolonged droughts, are expected to increase in coming years (IPCC, 2014INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE - IPCC. Climate Change 2014: Impacts, adaptation, and vulnerability. Cambridge: Cambridge University Press, 2014.). The drought has been the environmental stress that most have affected the world population, mainly because it reduces crop yields (Hu; Xiong, 2014HU, H.; XIONG, L. Genetic engineering and breeding of drought-resistant crops. Annual Review of Plant Biology, 65:715-741, 2014.). Important detrimental effects also have been reported for sugar cane production (Unica, 2014UNIÃO DA INDÚSTRIA DE CANA-DE-AÇÚCAR - UNICA. Quebra agrícola se acentua e previsão de moagem diminui para 546 milhões de toneladas na região centro-sul. 2014. Available in: <Available in: http://www.unicadata.com.br/listagem.php?idMn=80
>. Access in: 15 jul. 2016.
http://www.unicadata.com.br/listagem.php...
). In recent years the expansion of sugar cane cultives has been driven by increasing demand for biofuels around the world, requiring the occupation of new agricultural areas where water availability is highly variable or limited (Azevedo et al., 2011AZEVEDO, R. A. et al. Sugar cane under pressure: An overview of biochemical and physiological studies of abiotic stress. Tropical Plant Biology, 4:42-51, 2011.). Therefore, to tackle those conditions, the plants need to be able of uptake and use efficiently the mineral nutrients, in order to produce biomass even under water limitation (Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.). This ability is especially decisive in initial plant development, when the root system is still under formation (Zhao; Glaz; Comstock, 2010ZHAO, D.; GLAZ, B.; COMSTOCK, J. C. Sugarcane response to water-deficit stress during early growth on organic and sand soils. American Journal of Agricultural and Biological Sciences, 5(3):403-414, 2010.). Certainly the drought affects directly or indirectly the absorption and use of all mineral nutrients by plants (Mauad; Crusciol; Grassi Filho, 2011MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.). However, considering that the macronutrientes are required in large amounts to keep plant growth, the absorption of these elements would be greatly impaired under stressful conditions limiting plant production. Besides having vital nutritional and physiological importance in plants (Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.), the N, P and K together, correspond to about 25% of the cost of production for sugar cane plantation. There are strong evidences that the proper supply of these nutrients to plants relieves water stress (Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.). However, little is known on the opposite approach of how the drought affects the absorption and accumulation of these nutrients, by distinct sugar cane genotypes, or by different parts of a same plant. In first instance, it seems to be very clear that the movement of nutrients from the soil towards the root depends on the presence of water. In suitable moisture, it is known that N moves in the soil toward the plant roots mainly through the mass-flow phenomenon (Bonfim-Silva; Anicésio; Silva, 2013BONFIM-SILVA, E. M.; ANICÉSIO, E. C. A.; SILVA, T. J. A. Características morfológicas de cultivares de trigo submetidas à compactação do solo. Conhecer, 9(17):559-569, 2013.; Matimati; Verboom; Cramer, 2013MATIMATI, I.; VERBOOM, G. A.; CRAMER, M. D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. Journal of Experimental Botany, 14:2-10, 2013.), which is intense when the plants have high transpiration rates (Vieira; Mochel Filho, 2010VIEIRA, M. M. M.; MOCHEL FILHO, W. J. E. Influência dos fatores abióticos no fluxo de biomassa e na estrutura do dossel. Archivos de Zootecnia, 59(R):15-24, 2010.). At short distances, diffusion is the main way of P and K reach the plant roots surface, whereas over long distances the plant acquires these nutrients by the direct contact root-soil when the root system is in full growth (Crusciol et al., 2013CRUSCIOL, C. A. C. et al. Desenvolvimento radicular e aéreo, nutrição e eficiência de absorção de macronutrientes e zinco por cultivares de arroz de terras altas afetadas pela adubação fosfatada. Semina, 34(5):2061-2076, 2013.). The typically phisical phenomena, like difusion, are certainly affected in the same way, independently of the plant type cultured on the soil. However, the biological phenomena, as the root growth, evapotranspiration, nutrient uptake and its transport and use by the plants, may be very related to specific characteristics of each genotype. Some studies with soybean (Mauad; Crusciol; Grassi Filho, 2011MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.), wheat (Sun et al., 2012SUN, M. et al. Absorption and accumulation characteristics of nitrogen in diferent wheat cultivars under irrigated and dryland conditions. Australian Journal of Crop Science, 6(4):613-617, 2012.) and canola (Ashraf; Shahbaz; Ali, 2013ASHRAF, M.; SHAHBAZ, M.; ALI, Q. Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pakistan Journal of Botany, 45(1):93-98, 2013.), suggest that the degree of drought tolerance may be related to the ability of absorption and accumulation of mineral nutrients by plants, when subjected to low soil moisture conditions. In general, drought tolerance is the characteristic of a plant species, or of different varieties of the same species, to adapt in environments with low water potentials (Pincelli; Silva, 2012PINCELLI, R. P.; SILVA, M. A. Alterações morfológicas foliares em cultivares de cana-de-açúcar em resposta à deficiência hídrica. Bioscience Journal, 28(4):546-556, 2012.). Similarly, it is expected that differents cultivars show differents responses in the absorption and accumulation of N, P and K, when under water deficit. By its turn, it should be related to the ability to better withstand the stress phase (Waraish et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.). Regarding to sugar cane, these kind of information is still very scarce in the literature. Thus, if different varieties of sugar cane present different responses on this issue, this information could be used, independently or in conjunction with other features, to better choose those most suitable genotypes to be planted in regions subject to extended drought periods. Certainly, the use of tolerant cultivars in areas subjected to these extreme events is the best solution to address climate changes that are forecasted for the coming years (Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.; Lefèvre et al., 2012LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.). Considering the aforementioned, the objective of this study was to determine the content and accumulation of N, P and K in the aerial part of plant from three genotypes of sugar cane, submitted to three water potential in the soil, and check for relationship of these variables with the tolerance of plants to prolonged drought.
MATERIAL AND METHODS
Physical and chemical characteristics of the soil used in the assay
The soil used in the experiment was classified as a typical eutrophic oxisol, with a medium texture (EMBRAPA, 2013EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Sistema Brasileiro de Classificação de Solos. 3.ed. Brasília: Embrapa, 2013. 353p.). It was chemically analyzed according to described by Raij et al. (2001RAIJ, B. V. et al. Análise química para avaliação da fertilidade do solo. Campinas: IAC, 2001. 285p.), and presented the following results: pH (CaCl2) = 5.8; organic matter = 23 g dm-3; P (resin) = 76 mg dm-3; K = 5.7 mmolc dm-3; Ca = 49.0 mmolc dm-3; Mg = 26.0 mmolc dm-3; H+Al = 22.0 mmolc dm-3; SB = 80.7 mmolc dm-3; CEC = 102.7 mmolc dm-3; Saturation by bases = 79%.
In order to determine the water retention curve of this soil, some samples were collected from the pots with volumetric rings, and submitted to different potentials, in a pressure chamber (Klute, 1986KLUTE, A. Water retention: Laboratory methods. In: KLUTE, A. (ed.) . Methods of soil analysis. Part 1. 2 ed. Madison: ASA and SSSA, p. 635-662, 1986.), and the obtained water retention curve is shown in the Figure 1.

Curve of water retention of the soil (medium texture eutrophic oxisol) as function of soil water potential (Ψw). MPa_ megapascal.
Experimental conditions
The experiment was conducted from January to June 2012 in a greenhouse (temperature of 20 ± 4.3 °C, 67 ± 10% of relative humidity, average photoperiod of 12 hour, and 1800 mmol m2 h-1 irradiance) at São Paulo State University (coordinates: 21°15’22’’ S, 48°18’58’’ W, elevation: 575 m). Based on soil analysis, and following the indications of Raij et al. (1997RAIJ, B. V. et al. (Ed.). Recomendações de adubação e calagem para o estado de São Paulo. 2.ed. Campinas: IAC, 1997. 285p.), soil was fertilized with 222.0 mg dm-3 of simple superphosphate, and then 50 dm3 were transferred to pots. During filling of the pots was placed at the geometric center of each pot, a PVC tube (2.54 cm diameter) with perforations along the part that stayed below soil surface. This tube was used to ensure the rapid, and uniform distribution of water in all layers of the soil, during water reposition to maintain desired moisture.
Treatments and experimental design
The experiment consisted in 9 treatments, with 5 replicates, conducted in a completely randomized block design. The treatments were composed by the combination of three genotypes of sugar cane (SP81-3250, RB855453 and IACSP95-5000), under three levels of soil water potentials. The levels of soil hydric potentials studied were ranges of: -0.010 to -0.015 MPa, -0.050 to -0.055 MPa, and -0.075 to -0.080 MPa, which corresponds to no water restriction (ΨNR), moderated stress (ΨM), and severe water limitation (ΨS), respectively. Culm billets with 5 cm long containing one bud were used. These culm billets were planted in plastic cups of 0.5 dm3, filled with washed sand. At 30 days after plantlets emergence, the more uniform ones were selected to be used in the assay. Then, two plantlets were planted on each experimental units (pots containing 50 dm3 fertilized soil). At this time, also two tensiometers were installed in each experimental unit, the first one at 10 cm and the second at 30 cm depth. At 50 days after planting, a topdressing fertilization was performed with 66.0 mg dm-3 of potassium chloride and 91.0 mg dm-3 of urea (Raij et al., 1997RAIJ, B. V. et al. (Ed.). Recomendações de adubação e calagem para o estado de São Paulo. 2.ed. Campinas: IAC, 1997. 285p.). When the plants reached 60 days from planting, it was initiated the imposition of the desired levels of soil water potentials. The measurements of water potentials were performed daily, in the morning, of both tensiometers of each pot. Considering the average of these measurements, the estimation of soil moisture were determined by using information obtained from the curve (Figure 1). Then the amount of water needed to keep the desired soil water potential of each experimental unit, was calculated individually and added at each pot.
Plant transpiration measurements
When the plants reached 150 days after planting (90 days after imposition of distinct water potentials) the measurement of the transpiration was carried out in the morning (between 8h and 10h), in the middle portion of the +1 leaf, with a portable analyzer (LCPro + SYSTEM, ADC) connected to a light source, according to the methodology described by Marenco, Gonçalves and Vieira (2001MARENCO, R. A.; GONÇALVES, J. F. C.; VIEIRA, G. Leaf gas exchange and carbohydrates in tropical trees differing in successional status in two light environments in Central Amazonia. Tree Physiology, 21(18):1311-1318, 2001.). The evaluations were performed by adjusting the apparatus with a flux of CO2 equivalente to 370 ± 10 μmol m-2s-1, and PAPF (photosynthetically active photon flux) of 2000 μmol m-2s-1.
Plant samplings and its physical evaluations
Plants were cut close to the ground surface, the leaves and culms were separated and washed with distilled water. Then the samples were put into an oven with forced air circulation, at 65 °C, until constant weight. The dry mass of individual samples was determined with a 0,1g precision scale. Then the samples were ground in a Wiley mill and sieved (2 mm mesh), before submitted to chemical analysis.
Chemical analysis of samples
The N content was determined by the Kjeldahl method, after sulfuric digestion. Following nitric-perchloric digestion of the samples, the P content was determined spectrophotometrically by the method of molybdate-vanadate, and the K content was determined by atomic absorption spectrophotometry (Battaglia et al., 1983BATAGLIA, O. C. et al. Métodos de análise química de plantas. Campinas: IAC, 1983. 48p.). Based on these data, the concentration and the accumulation of nutrients in the plants shoots were calculated.
Statistical analysis
Data were submitted to analysis of variance by F test, in a 3 x 3 factorial arrangement (three varieties x three water potentials of soil). The means were compared by Tukey test at 5% probability. The calculations were performed by using the Sisvar program (Ferreira, 2011FERREIRA, D. F. Sisvar: A computer statistical analysis system. Ciência e Agrotecnologia, 35(6):1039-1042, 2011.).
RESULTS AND DISCUSSION
The F test pointed out significant effect (P≤0.01) for the interaction (G×Ψ), in almost all studied variables (Table 1). Plant transpiration was the only variable that showed significant value (P≤0.01) of F test for effect of genotype (G) and water potential (Ψ), independently. The data of P content in the culms were not affected by treatments (Table 1).
Summary of the analysis of variance (F test) for plant dry mass, content of nitrogen (N), phosphorus (P) and potassium (K) in leaves and culms, accumulation (in plant shoot) of N, P and K, and transpiration rate, of plants from three sugar cane genotypes (G) grown under three soil water potentials (Ψ).
Plants from genotypes SP81-3250 (SP), RB855453 (RB) and IACSP95-5000 (IAC), showed differents potentials to accumulate phytomass (dry mass) when grown with no water restrictions (ΨNR), with SP>RB>IAC (Figure 2). The biomass production was impaired by severe water limitation (ΨS) in all tested genotypes (Figure 2).
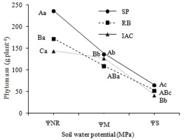
Phytomass (dry mass) of plants from three sugar cane genotypes (SP= SP81-3250, RB= RB855453, IAC= IACSP95-5000), grown under three soil water potentials (ΨNR = -0,010 to -0,015 MPa = no water restriction; ΨM= -0,050 to -0,055 MPa = moderate stress; ΨS= -0,075 to -0,080 MPa = severe water limitation). Capital letters were used to compare values within each water potential, while lowercase letters to compare values of a same genotype. Values followed by at least one same letter do not differ statistically (Tukey test, at P=0.05).
In spite of the IAC plants have shown smaller potential to accumulate dry mass under normal condition, these plants tended to maintain its biomass production when submitted to a moderate stress (ΨM), while those from other genotypes tended to decrease dry mass continuously with decreasing the water potential in the soil (Figure 2). By comparing dry mass of plants grown under ΨS, it was found the same sequence (SP>RB>IAC) observed in ΨNR. However, values were kept in a smaller range, without significant difference (P≤0.05) between adjacent values (Figure 2). Under water stress, Pincelli and Silva (2012PINCELLI, R. P.; SILVA, M. A. Alterações morfológicas foliares em cultivares de cana-de-açúcar em resposta à deficiência hídrica. Bioscience Journal, 28(4):546-556, 2012.) also obtained similar results in plants of the SP and RB genotypes, regarding biomass production.
It was observed an intense interaction of the genotype effect and water potential in the soil, on the results of N concentration in the leaf and culms, as well as total N accumulated in the aerial part of the plants (Table 1, Figure 3).

Content of nitrogen (N) in leaves (a) and culms (b), and accumulation in the shoot (c) of plants from three sugar cane genotypes (SP= SP81-3250, RB= RB855453, IAC= IACSP95-5000), grown under three soil water potentials (ΨNR= -0,010 to -0,015 MPa = no water restriction; ΨM= -0,050 to -0,055 MPa = moderate stress; ΨS= -0,075 to -0,080 MPa = severe water limitation). Capital letters were used to compare values within each water potential, while lowercase letters to compare values of a same genotype. Values followed by at least one same letter do not differ statistically (Tukey test, at P=0.05).
Although in plants under moderate stress, the N content have varied differently among genotypes, when under severe drought all plants shown lower N content, in comparison to non stressed plants (Figure 3a), what is in accordance to results found for rice (Mauad; Crusciol; Grassi Filho, 2011MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.) and wheat (Sun et al., 2012SUN, M. et al. Absorption and accumulation characteristics of nitrogen in diferent wheat cultivars under irrigated and dryland conditions. Australian Journal of Crop Science, 6(4):613-617, 2012.). The N content in culm tended to increase in RB plants under moderate water potential, while there was no significant variation (P≤0.05) in culms of plants from other studied genotypes (Figure 3b). In severe water limitation, plants of all tested genotypes shown less N accumulation, in comparison to plants under no water restriction (Figure 3c). Although plants of distinct genotypes have showed different N accumulation when grown without water limitation (SP> IAC> RB), total N uptake did not differ among genotypes under moderate and severe drought (Figure 3c).
The P contents in the leaves were lower in all plants under ΨS, when compared to those plants under ΨNR, and no difference among genotypes was observed (Figure 4a). Under moderate water limitation, plants of genotypes RB and IAC showed the same P levels observed in the leaves of the same plants under normal condition (ΨNR), while the values of this variable decreased in SP plants (Figure 4a). No effect of genotype and soil water potential was detected on P content and on P accumulation in the culms of the studied plants (Table 1).
The sequence of P accumulation in the shoot of plants of studied genotypes, was: SP>RB>IAC (Figure 4b). The P uptake by SP plants was reduced when the soil water potential was decreased. For IAC genotype, the P uptake was maintained similar only in plants under ΨNR and ΨM, but decreased in plants under ΨS. By other hand, plants of RB genotype were able to maintain the same P uptake even when the water limitation was changed from moderate to severe (Figure 4b).

Content of Phosphorus (P) in leaves (a), and P accumulation in the shoot (b) of plants from three sugar cane genotypes (SP= SP81-3250, RB= RB855453, IAC= IACSP95-5000), grown under three soil water potentials (ΨNR= -0,010 to -0,015 MPa = no water restriction; ΨM= -0,050 to -0,055 MPa = moderate stress; ΨS= -0,075 to -0,080 MPa = severe water limitation). Capital letters were used to compare values within each water potential, while lowercase letters to compare values of a same genotype. Values followed by at least one same letter do not differ statistically (Tukey test, at P=0.05).
For genotypes RB and SP, the K content in the leaves was higher for plants in ΨS, than under ΨNR. By the other hand, leaves of plants from the IAC genotype presented lower K levels under severe water limitation (Figure 5a). The data of K content in culms showed similar behaviour to that of K content in the leaves (Figure 5a, b). When comparing plants under ΨNR and ΨS, it can be observed that only the RB plants were able to maintain the K uptake, while plants of other tested genotypes reduced the K accumulation in the shoot (Figure 5c). The studied genotypes tended to show similar behaviour for accumulation of P and K in the plants shoot (Figure 4c, 5c).

Content of potassium (K) in leaves (a) and culms (b), and accumulation in the shoot (c) of plants from three sugar cane genotypes (SP= SP81-3250, RB= RB855453, IAC= IACSP95-5000), grown under three soil water potentials (ΨNR= -0,010 to -0,015 MPa = no water restriction; ΨM= -0,050 to -0,055 MPa = moderate stress; ΨS= -0,075 to -0,080 MPa = severe water limitation). Capital letters were used to compare values within each water potential, while lowercase letters to compare values of a same genotype. Values followed by at least one same letter do not differ statistically (Tukey test, at P=0.05).
As for plant transpiration rate, the RB and IAC genotypes did not differ, but showed higher values when compared to SP genotype (Figure 6a). The ΨM had small effect on plant transpiration rate, but ΨS significantly reduced plant transpiration for all studied genotypes (Figure 6b).

Medium effect of genotypes (a), and of soil water potential (b) on the transpiration rate (E) measured in leaves of plants from three sugar cane genotypes (SP= SP81-3250, RB= RB855453, IAC= IACSP95-5000), grown under three soil water potentials (ΨNR= -0,010 to -0,015 MPa = no water restriction; ΨM= -0,050 to -0,055 MPa = moderate stress; ΨS= -0,075 to -0,080 MPa = severe water limitation). Means followed by a same letter do not differ statistically (Tukey test, at P=0.05).
The total accumulation of N, P and K in the plants shoot were the variables that most significantly correlated (coefficients ≥ 0.89) among themselves, and with the production of plant dry matter, with highly significant values (P≤0.01) for correlation of Pearson (Table 2). For the aforementioned variables, it were also found significant correlations (P≤0.01) with plant transpiration rate, but with smaller coefficients (between 0.50 and 0.65). The content of nutrients in the leaves and culms showed no significant correlations (P≤0.05), or very low correlations (coefficients < 0.50) with transpiration rates, or with dry matter of studied plants (Table 2).
Pearson correlation among variables: plant dry mass (PDM), plant transpiration rate (TRA), and contents (g kg-1) of nitrogen (N), phosphorous (P) and potassium (K) in leaves (NL, PL and KL, respectively) and culms (NC, PC and KC, respectively), and accumulation (g plant-1) of N, P and K, in the plant shoot (NPS, PPS and KPS, respectively) measured on plants from three sugar cane genotypes grown under three soil water potentials. **_ significant at P≤0.01, *_ significant at P≤0.05, ns_ not significant (P≤0.05).
The lower biomass of plants kept under ΨS, compared to plants under ΨNR (Figure 2), must have been due to the reduction of the speed of physiological processes, with negative effects on plant growth, caused by lack of water (Mahmood; Ashraf ; Shahbaz, 2009MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a pre-sowing seed treatment improve growth and regulate some key physiological atributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, 41(3):1291-1302, 2009.), as well as due to the lower levels of N and P in the plants (Figure 3a, 3b, 4a). The smaller N and P levels observed in plants under ΨS, compared to plants grown in ΨNR (Figure 3a, 4a), probably was due to the limitations of the movement of these nutrients in the soil, what may have affected the absorption by plant roots. In the case of N, this statement is supported on the fact that N in the soil reaches the roots by mass flow (Bonfim-Silva; Anicésio; Silva, 2013BONFIM-SILVA, E. M.; ANICÉSIO, E. C. A.; SILVA, T. J. A. Características morfológicas de cultivares de trigo submetidas à compactação do solo. Conhecer, 9(17):559-569, 2013.; Crusciol et al., 2013CRUSCIOL, C. A. C. et al. Desenvolvimento radicular e aéreo, nutrição e eficiência de absorção de macronutrientes e zinco por cultivares de arroz de terras altas afetadas pela adubação fosfatada. Semina, 34(5):2061-2076, 2013.). By its turn, the intensity of this process depends on the level of plant transpiration, and also depends on the concentration of N in the soil solution, and soil water content (Matimati; Verboom; Cramer, 2013MATIMATI, I.; VERBOOM, G. A.; CRAMER, M. D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. Journal of Experimental Botany, 14:2-10, 2013.). Regarding to P, its supply to the roots is mainly driven by the diffusion process, which depends on soil moisture and root surface in contact with soil (Crusciol et al., 2013CRUSCIOL, C. A. C. et al. Desenvolvimento radicular e aéreo, nutrição e eficiência de absorção de macronutrientes e zinco por cultivares de arroz de terras altas afetadas pela adubação fosfatada. Semina, 34(5):2061-2076, 2013.). By its turn, soil moisture interferes directly in root development, influencing directly or indirectly in supplying P for plants (Mackay; Barber, 1985MACKAY, A. D.; BARBER, S. A. Soil moisture effect on potassium uptake by corn. Agronomy Journal, 77:519-523, 1985.). Taking into account that the soil water potential was reduced (from ΨNR to ΨS), the reduction of both concentration and accumulation of N and P were already expected in plants subjected to this very low soil water potentials (Figure 3a, 3b, 4a). Were also observed lower levels of N and P in shoots of canola, soybean and wheat plants subjected to intense water deficit (Mauad; Crusciol; Grassi Filho, 2011MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.; Sun et al., 2012SUN, M. et al. Absorption and accumulation characteristics of nitrogen in diferent wheat cultivars under irrigated and dryland conditions. Australian Journal of Crop Science, 6(4):613-617, 2012.; Ashraf; Shahbaz; Ali, 2013ASHRAF, M.; SHAHBAZ, M.; ALI, Q. Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pakistan Journal of Botany, 45(1):93-98, 2013.). The lower levels of N in the leaves and stems (Figure 3 a and b), in comparison to the that observed for P (Figure 4a), may be due to the low transpiration rate (Figure 6b), since it is known that this variable is responsible for regulating the mass flow process, which is preferably used by N to contact with plant roots (Bonfim-Silva; Anicésio; Silva, 2013BONFIM-SILVA, E. M.; ANICÉSIO, E. C. A.; SILVA, T. J. A. Características morfológicas de cultivares de trigo submetidas à compactação do solo. Conhecer, 9(17):559-569, 2013.; Matimati; Verboom; Cramer, 2013MATIMATI, I.; VERBOOM, G. A.; CRAMER, M. D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. Journal of Experimental Botany, 14:2-10, 2013.). Therefore, the low plant transpiration induced by drought should have impaired most intensely the contact of N with root surface, than that of the P, what is in accordance to tha mentioned by Mahmood, Ashraf and Shahbaz (2009)MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a pre-sowing seed treatment improve growth and regulate some key physiological atributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, 41(3):1291-1302, 2009., and to that observed in this research (Figure 6b). The K concentration in plants grown without water restriction remained in the range considered adequate by Raij et al. (1997RAIJ, B. V. et al. (Ed.). Recomendações de adubação e calagem para o estado de São Paulo. 2.ed. Campinas: IAC, 1997. 285p.). However, the content of this nutrient in the leaves and stems were increased in plants of SP genotype under ΨS, and in the leaves of RB plants (Figure 5a, 5b). This result was not expected, if considered only the soil phenomena, since the low humidity may reduce the movement of the K in the soil (Lefèvre et al., 2012LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.; Crusciol et al., 2013CRUSCIOL, C. A. C. et al. Desenvolvimento radicular e aéreo, nutrição e eficiência de absorção de macronutrientes e zinco por cultivares de arroz de terras altas afetadas pela adubação fosfatada. Semina, 34(5):2061-2076, 2013.). But there is increasing evidences that plants subjected to drought tends to increase K concentration, and have a greater exigency of K by its tissues (Mahmood; Ashraf ; Shahbaz, 2009MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a pre-sowing seed treatment improve growth and regulate some key physiological atributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, 41(3):1291-1302, 2009.; Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.). The reason for the increased need for K by the plants that undergo stress appears to be related to the fact that K be necessary in maintaining the CO2 photosynthetic fixation rate, also to maintain the osmotic potential and the turgor of the cells, and yet to keep the regulation of stomatal conductance (Cakmak, 2005CAKMAK, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 168:521-530, 2005.; Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.; Lefèvre et al., 2012LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.). The effect of K on the maintenance of photosynthetic CO2 fixation rate, seems to be related the participation of K on the oxidation of NADPH oxidase, thus preventing the formation of the reactive oxygen species (ROS), which are so harmful to the cells life (Cakmak, 2005CAKMAK, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 168:521-530, 2005.; Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.). Lefèvre et al. (2012)LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012. also observed higher K levels in potato cultivars tolerant to drought and ascribed such responses to the water homeostasis, and regulation of the turgor of the studied plants.
Generally it is assumed that the drought affects the availability of minerals from the soil for the plants (Mauad; Crusciol; Grassi Filho, 2011MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.; Sun et al., 2012SUN, M. et al. Absorption and accumulation characteristics of nitrogen in diferent wheat cultivars under irrigated and dryland conditions. Australian Journal of Crop Science, 6(4):613-617, 2012.; Ashraf; Shahbaz; Ali, 2013ASHRAF, M.; SHAHBAZ, M.; ALI, Q. Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pakistan Journal of Botany, 45(1):93-98, 2013.). However, in several literature reports (Cakmak, 2005CAKMAK, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 168:521-530, 2005.; Waraich et al., 2011WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.; Lefèvre et al., 2012LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.) as well as in the results of the present study (Figure 5b), the contents of K remained at levels considered adequate for good plant growth, despite the strong decrease in water potential of the soil. Lefèvre et al., (2012)LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012. ascribed the lack of damage in the K availability to plants under drought, to soil enzymes which may play an important role in mineralization and availability of nutrients for plants, especially in soil with high content of organic matter. However, this phenomenon should be much more studied to be completely elucidated. Anyway, the increment in K concentration seems to be an useful feature to take into account in the selection of cultivars tolerant to drought (Cakmak, 2005CAKMAK, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 168:521-530, 2005.; Lefèvre et al., 2012LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.). Pincelli and Silva (2012PINCELLI, R. P.; SILVA, M. A. Alterações morfológicas foliares em cultivares de cana-de-açúcar em resposta à deficiência hídrica. Bioscience Journal, 28(4):546-556, 2012.) also obtained similar biomass production for plants of the genotypes SP and RB, reinforcing the results of this study, on that these are plants with similar tolerance, to water deficit. The greater dry matter accumulation in plants of the genotypes SP and IAC (Figure 2), seems to be related to the capacity of these plants to absorb N from the soil, even under severe water limitation (Figure 3c, Table 2). The low accumulation of N, P and K in plants under ΨS, compared to plants grown in ΨNR, is consequence of lower contents of N, P and K and lower dry weight of plants (Figure 2 to Figure 5 and Table 2). It should be noted that the SP plants showed higher accumulation of P and K, in comparison to other tested genotypes, even when grown under ΨS (Figure 4b , 5c). In addition, these results indicated that the better development of plants SP under ΨS in relation to the other genotypes, was due to higher N and K contents in the leaves and culms (Figure 3a, 3b, 5a, 5b) and higher accumulation of P and K in the plant shoot (Figure 4b, 5c), resulting in a superior dry matter accumulation. Thus, this genotype is more suitable than the IAC or RB, for planting in environments that undergo long periods of water restriction. Interestingly, there was no significant (P≤0.05) correlation between transpiration rate with the concentration of K or P in the culms or leaves. However, highly positive significant correlations (P≤0.01) were observed among transpiration rate and accumulation of K and P in the plant shoot (Table 2). The N content in the leaves also seems to be important to maintain plant transpiration, as indicated by the positive and relatively high correlations values (Table 2). This findings is in accordance with other studies suggesting that N availability partially regulates the transpiration rate, and also that transpiration modulates the acquisition of other nutrients (Matimati; Verboom; Cramer, 2013MATIMATI, I.; VERBOOM, G. A.; CRAMER, M. D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. Journal of Experimental Botany, 14:2-10, 2013.).
CONCLUSIONS
The tolerance to drought in sugar cane is related to the high levels of N and K in the leaves and stems individually, and large accumulations of K and P in the plant shoot. There are strong positive correlations among accumulation of N, P and K in the plant shoot and dry matter production of plants submitted to drought, while there are only intermediate coeficients of correlations among plant transpiration and nutrient uptake by plants under drought. Plants belonging to the genotype SP81-3250 are more tolerant to prolonged drought, than the RB855453 and IACSP95-5000 plants.
ACKNOWLEDGMENTS
The authors Dr. José Frederico Centurion (Dep. solos e adubos - Unesp / Jaboticabal) and Dr. Luis Carlos Pavani (Dep. Engenharia Rural, Unesp/Jaboticabal), for their technical support on soil characterization, and on handling, calibration and instalations of tensiometers, used in this work. As well as the the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
REFERENCES
- ASHRAF, M.; SHAHBAZ, M.; ALI, Q. Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pakistan Journal of Botany, 45(1):93-98, 2013.
- AZEVEDO, R. A. et al. Sugar cane under pressure: An overview of biochemical and physiological studies of abiotic stress. Tropical Plant Biology, 4:42-51, 2011.
- BATAGLIA, O. C. et al. Métodos de análise química de plantas. Campinas: IAC, 1983. 48p.
- BONFIM-SILVA, E. M.; ANICÉSIO, E. C. A.; SILVA, T. J. A. Características morfológicas de cultivares de trigo submetidas à compactação do solo. Conhecer, 9(17):559-569, 2013.
- CAKMAK, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 168:521-530, 2005.
- CRUSCIOL, C. A. C. et al. Desenvolvimento radicular e aéreo, nutrição e eficiência de absorção de macronutrientes e zinco por cultivares de arroz de terras altas afetadas pela adubação fosfatada. Semina, 34(5):2061-2076, 2013.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Sistema Brasileiro de Classificação de Solos. 3.ed. Brasília: Embrapa, 2013. 353p.
- FERREIRA, D. F. Sisvar: A computer statistical analysis system. Ciência e Agrotecnologia, 35(6):1039-1042, 2011.
- HU, H.; XIONG, L. Genetic engineering and breeding of drought-resistant crops. Annual Review of Plant Biology, 65:715-741, 2014.
- INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE - IPCC. Climate Change 2014: Impacts, adaptation, and vulnerability. Cambridge: Cambridge University Press, 2014.
- KLUTE, A. Water retention: Laboratory methods. In: KLUTE, A. (ed.) . Methods of soil analysis. Part 1. 2 ed. Madison: ASA and SSSA, p. 635-662, 1986.
- LEFÈVRE, I. et al. Drought impacts mineral contentes in Andean potato cultivars. Journal of Agronomy and Crop Science, 198:196-206, 2012.
- MACKAY, A. D.; BARBER, S. A. Soil moisture effect on potassium uptake by corn. Agronomy Journal, 77:519-523, 1985.
- MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a pre-sowing seed treatment improve growth and regulate some key physiological atributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, 41(3):1291-1302, 2009.
- MARENCO, R. A.; GONÇALVES, J. F. C.; VIEIRA, G. Leaf gas exchange and carbohydrates in tropical trees differing in successional status in two light environments in Central Amazonia. Tree Physiology, 21(18):1311-1318, 2001.
- MATIMATI, I.; VERBOOM, G. A.; CRAMER, M. D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. Journal of Experimental Botany, 14:2-10, 2013.
- MAUAD, M.; CRUSCIOL, C. A. C.; GRASSI FILHO, H. Produção de massa seca e nutrição de cultivares de arroz de terras altas sob condições de déficit hídrico e adubação silicatada. Semina. Ciências Agrárias, 32(3):939-948, 2011.
- SUN, M. et al. Absorption and accumulation characteristics of nitrogen in diferent wheat cultivars under irrigated and dryland conditions. Australian Journal of Crop Science, 6(4):613-617, 2012.
- PINCELLI, R. P.; SILVA, M. A. Alterações morfológicas foliares em cultivares de cana-de-açúcar em resposta à deficiência hídrica. Bioscience Journal, 28(4):546-556, 2012.
- RAIJ, B. V. et al. Análise química para avaliação da fertilidade do solo. Campinas: IAC, 2001. 285p.
- RAIJ, B. V. et al. (Ed.). Recomendações de adubação e calagem para o estado de São Paulo. 2.ed. Campinas: IAC, 1997. 285p.
- UNIÃO DA INDÚSTRIA DE CANA-DE-AÇÚCAR - UNICA. Quebra agrícola se acentua e previsão de moagem diminui para 546 milhões de toneladas na região centro-sul. 2014. Available in: <Available in: http://www.unicadata.com.br/listagem.php?idMn=80 >. Access in: 15 jul. 2016.
» http://www.unicadata.com.br/listagem.php?idMn=80 - VIEIRA, M. M. M.; MOCHEL FILHO, W. J. E. Influência dos fatores abióticos no fluxo de biomassa e na estrutura do dossel. Archivos de Zootecnia, 59(R):15-24, 2010.
- WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science , 5(6):764-777, 2011.
- ZHAO, D.; GLAZ, B.; COMSTOCK, J. C. Sugarcane response to water-deficit stress during early growth on organic and sand soils. American Journal of Agricultural and Biological Sciences, 5(3):403-414, 2010.
Publication Dates
-
Publication in this collection
Mar-Apr 2017
History
-
Received
19 July 2016 -
Accepted
23 Nov 2016






