Abstracts
Hoplias malabaricus, the common trahira, extended its range into the Iguaçu River sometime in the last decades. To determine levels of genetic differentiation in neighboring basins of this nominal taxon, as well as to unveil the source of the invading population, we analyzed patterns of similarity of RAPD-PCR genomic markers of samples from Paraná, Iguaçu and Tibagi Rivers. The high genetic diversity of samples from the Paraná and Tibagi Rivers suggested strong population structuring or even the occurrence of undescribed species. All alleles of the sample from the headwaters of Tibagi River were present in the sample from the Iguaçu River, which suggests that the former population or another population with similar genetic makeup, may be the source of the trahiras of the Iguaçu River.
Hoplias malabaricus, a traíra, colonizou o Rio Iguaçu nas últimas décadas. Numa tentativa de determinar os níveis de diferenciação genética deste taxon nominal em bacias vizinhas, assim como a origem da população colonizadora, foram analisados os padrões de similaridade de marcadores genômicos do tipo randomly amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR) de amostras dos rios Paraná, Iguaçu, e Tibagi. As amostras do Paraná e do Tibagi apresentaram elevados índices de diversidade genética, os quais sugerem a ocorrência de espécies diferentes nestes rios. Todos os alelos presentes na população da cabeceira do rio Tibagi ocorreram também na amostra do rio Iguaçu. Esta semelhança permite-nos sugerir que a população da cabeceira do rio Tibagi (ou alguma população com características genéticas semelhantes) possa ser a origem da população amostrada no rio Iguaçu.
Molecular biogeography of the Neotropical fish Hoplias malabaricus (Erythrinidae:Characiformes) in the Iguaçu, Tibagi, and Paraná Rivers* * Part of a thesis presented by J.A.D. to the Colorado State University, in partial fulfillment of the Ph.D. degree.
Jorge A. Dergam1, Harumi Irene Suzuki2, Oscar A. Shibatta3, Luiz F. Duboc4, Horácio F. Júlio Jr.2, Lúcia Giuliano-Caetano3 and William C. Black IV4
1 Departamento de Biologia Animal, Universidade Federal de Viçosa, 36571-000 Viçosa, MG, Brasil. Send correspondence to J.A.D. E-mail: dergam@mail.ufv.br
2 NUPELIA, Bloco H-90, Av. Colombo, 5790, Universidade Estadual de Maringá, 87020-900 Maringá, PR, Brasil.
3 CCB/BAV, UEL, 86051-970 Londrina, PR, Brasil.
4 Department of Microbiology, Colorado State University, Fort Collins, CO 80523, USA.
ABSTRACT
Hoplias malabaricus, the common trahira, extended its range into the Iguaçu River sometime in the last decades. To determine levels of genetic differentiation in neighboring basins of this nominal taxon, as well as to unveil the source of the invading population, we analyzed patterns of similarity of RAPD-PCR genomic markers of samples from Paraná, Iguaçu and Tibagi Rivers. The high genetic diversity of samples from the Paraná and Tibagi Rivers suggested strong population structuring or even the occurrence of undescribed species. All alleles of the sample from the headwaters of Tibagi River were present in the sample from the Iguaçu River, which suggests that the former population or another population with similar genetic makeup, may be the source of the trahiras of the Iguaçu River.
INTRODUCTION
Knowledge of geographical distribution patterns of freshwater fish in the Neotropical Region is still quite scarce. Research is complicated by insufficient sampling, poor accessibility of type material, and high levels of diversity of the fish fauna of this region (Böhlke et al., 1979). This is the case for the characin Hoplias malabaricus (Bloch, 1794), which has considerable morphological variation among and within populations (Eigenmann and Eigenmann, 1889) making it difficult to determine the origin of the recently established population in the Iguaçu River (southeastern Brazil) (Figure 1).
- Geographic location of samples. A, Localities from Argentina: Paraná River in Corrientes (PA/CO), and Paraná River in Reconquista (PA/RE); the square represents the region detailed in B; B, localities from the Rio Tibagi: Sertanópolis = TB/ST; Londrina = TB/LO, and Ponta Grossa = TB/PG; localities the Paraná River in Porto Rico = PA/PR; the Iguaçu: Segredo Reservoir (IG/SE). The Iguaçu falls (Cataracts) are also indicated.
The basin of the Iguaçu River extends from the western slope of a coastal relief, Serra do Mar, to the Paraná River, along with several other rivers including the Tibagi and Ivaí. The Iguaçu is isolated from the Paraná River by the Iguaçu falls. These falls date from the Oligocene (Sampaio, 1988). The fall of approximately 100 m constitutes a formidable barrier for upstream dispersal of fish fauna of the Paraná into the Iguaçu. This long term isolation certainly played a critical role in the endemism of fish fauna of the Iguaçu, which was considered by Haseman (1911) as its most remarkable feature. A recent survey by the limnology research group (NUPELIA) of the Universidade Estadual de Maringá identified 52 fish species in the Iguaçu River (FUEM/NUPELIA/COPEL, 1995), 75% of which were considered endemic, 17% indigenous but not endemic, and 8% introduced.
Haseman (1911) reported the absence of H. malabaricus in the Iguaçu River, while Godoy (1979) gave the first published account of its presence. Locals assert that H. malabaricus was likely introduced during the last decades, perhaps during aquaculture stocking of other exotics such as carp and tilapia (Sampaio, 1988). These exotics are also established in the Iguaçu (FUEM/NUPELIA/COPEL, 1995).
To unravel the question of the origin of the Iguaçu River population of H. malabaricus, we analyzed patterns of variation of nuclear markers of one sample from the Iguaçu, and six samples from two neighboring basins. This information may be relevant for preventing (or explaining) invasions of other species of exotic fishes in the Iguaçu, as well as for future assessments of the degree of endemism of the Iguaçu River fish fauna.
MATERIAL AND METHODS
One hundred thirty-two specimens were collected from Brazil and Northern Argentina (Figure 1 and Table I). Tissue collection and DNA extraction are described in Dergam (1996). Molecular analyses were done at the Department of Microbiology, Colorado State University, Fort Collins, CO, USA.
We used RAPD-PCR (Williams et al., 1990), with primers OPA09, OPA05, and OPAC13 (Operon Technologies). All samples were run simultaneously for each primer reaction to minimize repeatability problems (Black, 1993). We used a binary matrix ("1" and "0" for presence and absence of bands, respectively) for each specimen to record bands for cluster analysis. A measure of distance was calculated for each pairwise comparison, yielding a matching matrix (where similarity was assessed by the shared presence or absence of bands) with the RAPDPLOT program (Kambhampati et al., 1991; Black, 1996). This program calculates the fraction of matches between all individuals in the matrix:
M = NAB/NT
where NAB = total number of matches (shared presence or absence of bands) between individuals A and B, and NT is the total number of loci of the binary matrix.
Molecular similarity was analyzed by unweighted pair-group mean average (UPGMA) cluster analysis computed with the NEIGHBOR option of PHYLIP 3.5C (Felsenstein, 1993). A phenogram was drawn with the DRAWGRAM program from the same computer package. Because this similarity analysis is exclusively qualitative and does not deal with allelic frequencies or variances at each locus, we assumed that uneven sample sizes did not affect our results.
RESULTS
RAPD primers amplified a total of 74 bands in all samples taken together (Figure 2). High levels of within-population genetic variation were suggested by a positive correlation between the number of scorable bands and sample size (r = 0.82). The population with the highest number of scorable bands was Porto Rico (51 bands), while the lowest number of bands was observed for Londrina (27 bands).
- RAPD bands amplified with primer C-13 (45 specimens, one per lane). Codes as in Figure 1. "L" lanes are molecular markers of 1 kb, numbers on the left indicate bands with known molecular weight.
Although in many cases the alleles were not fixed (not present in all individuals of a given sample), some alleles were unique for some populations: in the Tibagi River, five alleles were unique to the Sertanópolis population, and one was unique to the Londrina population. In the Iguaçu, only one allele was unique to the Segredo population, and in the Paraná River, three alleles characterized the Porto Rico population, while two alleles were typical of the Reconquista population.
Five alleles were shared by at least one member of each population. Excluding alleles shared by members of all populations, the number of shared alleles among populations was also significantly correlated with sample size (r = 0.77).
The samples from Tibagi River and the Paraná River displayed high levels of variation, while the samples from Ponta Grossa and Segredo were very similar (Figure 2). All alleles of Ponta Grossa (Tibagi River) were also present in the population of Segredo (Iguaçu). The reverse did not happen, because the population from Segredo had 48 alleles, one third more than the population of Ponta Grossa (Figure 2).
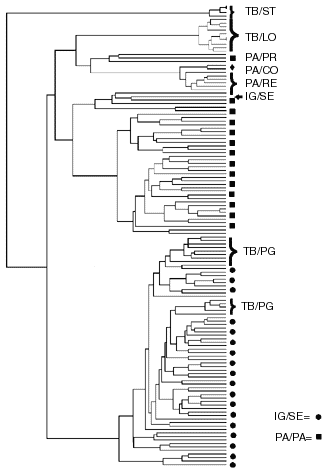
Molecular similarity was represented by a phenogram (Figure 3). The matching matrix grouped all specimens into three major clusters. The first cluster was composed of four specimens from the Tibagi River at Sertanópolis. The second cluster included all specimens from samples of the Paraná River at Porto Rico, Reconquista, and Corrientes. This major cluster also included all specimens from Tibagi at Londrina, plus one of the specimens from the Tibagi at Sertanópolis, and another from the Iguaçu at Segredo. The third cluster joined all the specimens from the Tibagi at Ponta Grossa, and almost all the specimens from Segredo.
- Phenogram of overall molecular similarity of RAPD bands among specimens from all localities at Tibagi, Paraná, and Iguaçu Rivers. Codes as in Figure 1.
DISCUSSION
The RAPD-PCR patterns indicate strong genetic differentiation within and among populations in the Paraná and the Tibagi Rivers. In the Paraná River at Porto Rico, the high levels of allelic polymorphism may represent the pristine composition of different species or populations within the H. malabaricus species complex, or they may be the outcome of faunal mixing caused by the closing of the Itaipu Dam in 1978 and the consequent flooding of Sete Quedas, another Oligocenic geographic barrier (Sampaio, 1988). Sete Quedas was the natural boundary for the Upper and Middle Paraná basins. The presence of an outlier in the Paraná River similar to the Iguaçu population suggests that the Paraná may indeed be a recipient of trahiras of diverse origins.
The Londrina sample from the Tibagi River is most similar to the populations of the Paraná (Figures 1, 2, and 3). On the other hand, while the Londrina sample is genetically homogeneous, the sample from Sertanópolis included one specimen with a molecular profile similar to Londrina, and the remaining four specimens different from all other samples along the Tibagi River (Figure 3). These well-defined clusters suggest that at least three yet undescribed species, or highly differentiated populations of H. malabaricus, co-exist in the Tibagi River. A similar situation was reported for H. malabaricus in the Aguapey River (Province of Corrientes, Argentina) (Dergam, 1996). To test these hypotheses, sequencing and morphological analyses of voucher material from the Tibagi River are currently being done by J.A.D. The patterns of genomic variation obtained with RAPDs suggest a close kinship between the newly established population of H. malabaricus in the Iguaçu and the sample from the Tibagi at Ponta Grossa. Lower genetic variation of RAPDs in the Tibagi sample than that observed in the Iguaçu/Segredo sample may be either an artifact of small sample size, the outcome of random genetic drift, or a combination of both factors: the Tibagi/Ponta Grossa sample came from a lagoon close to the headwaters of the Tibagi.
Although Haseman (1911) described the division between the Tibagi and the Iguaçu as "short and low", molecular similarity is not sufficient to support or reject the possibility that founder animals came from the Tibagi. The possibility of the existence of other source populations is supported by the fact that small samples (N = 3 each) from coastal populations of this species (Perequê River in Paranaguá and Ribeira River at Registro) display patterns of variation of RAPD alleles similar to the sample from Tibagi/Ponta Grossa (Dergam, 1996). Phylogenetic sequence analyses of the mitochondrial DNA of the Ponta Grossa, Segredo, and coastal populations corroborate that they are more closely related to each other than to any of the populations from the Tibagi or Paraná, and that all those populations are members of a more inclusive coastal clade (Dergam, 1996).
High levels of genetic diversity (genomic and mtDNA data) in the Tibagi and the Paraná Rivers suggest the existence of differentiated populations or species within the H. malabaricus species complex. Moreover, the introduced population of H. malabaricus in the Iguaçu was most likely derived from populations with a genetic composition similar to the sample from the headwaters of the Tibagi.
ACKNOWLEDGMENTS
Alberto Fenocchio kindly provided samples from Argentina. J.A.D. was supported by a travel and study from CAPES (Process No. 2732/92).
RESUMO
Hoplias malabaricus, a traíra, colonizou o Rio Iguaçu nas últimas décadas. Numa tentativa de determinar os níveis de diferenciação genética deste taxon nominal em bacias vizinhas, assim como a origem da população colonizadora, foram analisados os padrões de similaridade de marcadores genômicos do tipo randomly amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR) de amostras dos rios Paraná, Iguaçu, e Tibagi. As amostras do Paraná e do Tibagi apresentaram elevados índices de diversidade genética, os quais sugerem a ocorrência de espécies diferentes nestes rios. Todos os alelos presentes na população da cabeceira do rio Tibagi ocorreram também na amostra do rio Iguaçu. Esta semelhança permite-nos sugerir que a população da cabeceira do rio Tibagi (ou alguma população com características genéticas semelhantes) possa ser a origem da população amostrada no rio Iguaçu.
(Received July 7, 1997)
- Black IV, W.C. (1993). PCR with arbitrary primers: approach with care. Insect Mol. Biol. 2: 1-6.
- Black IV, W.C. (1996). Statistical analysis of arbitrarily primed PCR patterns in molecular taxonomic studies. In: Methods in Molecular Biology, Vol. 50: Species Diagnostics. PCR and Other Nucleic Acid Methods (J.P. Clapp Humana Press Inc., ed.), Totowa, NJ, pp. 39-55.
- Bloch, M.E. (1794). Der malabarische hecht. In: Naturgeschichte des Auslandische Fische Berlin, pp. 149-150.
- Böhlke, J.E., Weitzman, S.H. and Menezes, N.A. (1979). Estado atual da sistemática dos peixes de água doce da América do Sul. Acta Amazonica 8: 657-677.
- Dergam, J.A. (1996). Phylogeography and character congruence within the Hoplias malabaricus Bloch, 1794 (Erythrinidae, Characiformes, Ostariophysi) species complex. Ph.D. Diss., Colorado State University, Fort Collins, Colorado.
- Eigenmann, C.H. and Eigenmann, R.S. (1889). A review of the Erythrininae. Proc. Calif. Acad. Sci. 2: 100-117.
- Felsenstein, J. (1993). PHYLIP (phylogeny inference package) v3.5c Department of Genetics, University of Seattle, WA.
- FUEM/NUPELIA/COPEL (1995). Relatório anual do Projeto "Ictiologia e aquicultura experimental para o Reservatório de Segredo". March 1993 - February 1994. Fundação Universidade Estadual de Maringá, Maringá, PR, Brasil.
- Godoy, M.P. (1979). Rio Iguaçu, Paraná, Brasil - Reconhecimento da ictiofauna, modificaçőes ambientais e usos múltiplos dos reservatórios. Eletrosul, Rio de Janeiro, pp. 33.
- Haseman, J.D. (1911). Some new species of fishes from the Rio Iguassu. Ann. Carnegie Mus. 7: 374-387.
- Kambhampati, S., Black IV, W.C. and Rai, K.S. (1991). RAPD-PCR for identification and differentiation of mosquito species and populations: techniques and statistical analysis. J. Med. Ent. 4: 203-221.
- Sampaio, F.A.A. (1988). Estudos taxonômicos preliminares dos Characiformes (Teleostei, Ostariophysi) da bacia do Rio Iguassu, com comentários sobre o endemismo dessa fauna. Unpubl. Master's thesis, Universidade Federal de Săo Carlos, Săo Carlos, SP, Brasil.
- Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A. and Tingey, S.V. (1990). DNA polymorphisms amplified by arbitrary primers are useful genetic markers. Nucleic Acids Res. 18: 6531-6535.
Publication Dates
-
Publication in this collection
01 Mar 1999 -
Date of issue
Dec 1998
History
-
Received
07 July 1997