Abstract
Damselfishes (Pomacentridae, Perciformes) occur in all major oceans of the world and, with approximately 320 species, represent one of the most diverse families of marine Teleostei. The taxonomy of these reef fishes is problematic because of the large number of complex species and the range of color patterns they display, which vary among individuals and populations of the same species. In this study, we examined the cytogenetic composition of four species of Stegastes (S. pictus, S. fuscus, S. variabilis and S. leucostictus) found along the coast of Brazil. Stegastes pictus had a chromosomal number of 2n = 48 (14m+28sm+2st+4a, fundamental number (FN) = 92), S. fuscus had 2n = 48 (20m+22sm+6a, FN = 90), S. variabilis had 2n = 48 (18m+22sm+8a, FN = 88), and S. leucostictus had 2n = 48, (18m+22sm+8a, FN = 88). The nucleolar organizing regions were single and homologous in all of the species, and were located in the interstitial region on the short arm of the first submetacentric pair. The heterochromatin segments were reduced in size and were distributed conservatively over the centromeric and pericentromeric regions of most of the chromosomes. The marked divergence in the number of chromosomal arms, compared to other perciformes (2n = 48, FN = 48), indicated that varying degrees of multiple pericentric inversions had occurred during the karyotypic evolution of the Pomacentridae. Subtle karyotypic differences between S. variabilis and S. leucostictus suggested a recent divergence or that their karyotypes were less susceptible to changes. These results indicate that cytogenetic analyses could provide important complementary data for the characterization of populations and species of Stegastes and damselfishes in general.
pericentric inversions; pomacentridae; Stegastes; 5S genes
ANIMAL GENETICS
RESEARCH ARTICLE
Multiple pericentric inversions and chromosomal divergence in the reef fishes Stegastes (Perciformes, Pomacentridae)
Wagner Franco MolinaI; Pedro Manoel Galetti Jr.II
IUniversidade Federal do Rio Grande do Norte, Departamento de Biologia Celular e Genética, Natal, RN, Brazil
IIUniversidade Federal de São Carlos, Departamento de Genética e Evolução, São Carlos, SP, Brazil
Correspondence Correspondence to Wagner Franco Molina Universidade Federal do Rio Grande do Norte Departamento de Genética e Biologia Molecular Campus Universitário 59078-970 Natal, RN, Brazil E-mail: molinawf@yahoo.com.br
ABSTRACT
Damselfishes (Pomacentridae, Perciformes) occur in all major oceans of the world and, with approximately 320 species, represent one of the most diverse families of marine Teleostei. The taxonomy of these reef fishes is problematic because of the large number of complex species and the range of color patterns they display, which vary among individuals and populations of the same species. In this study, we examined the cytogenetic composition of four species of Stegastes (S. pictus, S. fuscus, S. variabilis and S. leucostictus) found along the coast of Brazil. Stegastes pictus had a chromosomal number of 2n = 48 (14m+28sm+2st+4a, fundamental number (FN) = 92), S. fuscus had 2n = 48 (20m+22sm+6a, FN = 90), S. variabilis had 2n = 48 (18m+22sm+8a, FN = 88), and S. leucostictus had 2n = 48, (18m+22sm+8a, FN = 88). The nucleolar organizing regions were single and homologous in all of the species, and were located in the interstitial region on the short arm of the first submetacentric pair. The heterochromatin segments were reduced in size and were distributed conservatively over the centromeric and pericentromeric regions of most of the chromosomes. The marked divergence in the number of chromosomal arms, compared to other perciformes (2n = 48, FN = 48), indicated that varying degrees of multiple pericentric inversions had occurred during the karyotypic evolution of the Pomacentridae. Subtle karyotypic differences between S. variabilis and S. leucostictus suggested a recent divergence or that their karyotypes were less susceptible to changes. These results indicate that cytogenetic analyses could provide important complementary data for the characterization of populations and species of Stegastes and damselfishes in general.
Key words: pericentric inversions, pomacentridae, Stegastes, 5S genes
Introduction
The family Pomacentridae (Perciformes) contains 28 genera and approximately 320 species known as damselfishes. This family is one of the most diverse among marine teleosts and occurs in tropical, sub-tropical, and temperate regions of all the major oceans (Nelson, 1994). The taxonomy of damselfishes is complicated by the large number of complex species and the color patterns that vary among individuals and populations of the same species. Several species are of growing economic interest because of their diverse color patterns, and this has led to their exploitation.
Several reports have provided karyotypic descriptions for the Pomacentridae, especially from the Pacific region, which has the greatest variety of damselfishes (Ojima, 1983; Klinkhardt et al., 1995). However, very little is known about structural aspects of the karyotype, such as the heterochromatic patterns and the number, position and frequency of the nucleolar organizing regions (NORs).
In this work, the karyotypes of the damselfishes Stegastes fuscus, S. pictus, S. variabilis, and S. leucostictus found along the coast of Brazil have been described. FISH with probes of the ribosomal 5S subunit were used on the species S. fuscus and S. variabilis, and rDNA 18S probes on S. fuscus.
Material and Methods
Cytogenetic analyses were done using 24 Stegastes fuscus (11 males and 13 females) and seven S. variabilis (4 males and 3 females) from the coast of the State of Rio Grande do Norte in northeastern Brazil, and four S. leucostictus (2 juveniles, 1 female and 1 male) and five S. pictus (3 males and 2 females) collected off the coast of the State of Bahia in northeastern Brazil. Voucher specimens were deposited in the Museum of Zoology of the Universidade Federal da Paraíba. Chromosomal preparations were obtained from kidney tissue dissociated in 9.5 mL of RPMI 1640 culture medium with five drops of colchicine for 30 min followed by hypotonic treatment for 25 min at room temperature. The material was then fixed in methanol:acetic acid (3:1, v/v). The NORs and the heterochromatic regions were identified by the methods of Howell and Black (1980) and Sumner (1972), respectively. Fluorescence in situ hybridization (FISH) was done in S. fuscus and S. variabilis using a probe from the 5S rRNA gene of the fish Leporinus elongatus (Anostomidae, Characiformes) (Martins and Galetti, 1999). An 18S rDNA probe was obtained from total DNA of Prochilodus affinis (Characiformes) (kindly provided by Dr. T. Hatanaka Universidade Federal de São Carlos) by PCR amplification using the probes NS1 (5'-GTAGTCATATGCTTGTCTC-3') and NS8 (5'-tccgcaggttcacctacgga-3') (White et al., 1990; Hizume, 1994). This rDNA probe was used to investigate the major ribosomal sites in S. fuscus.
Results
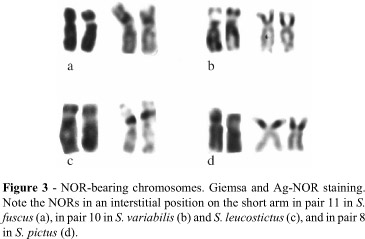
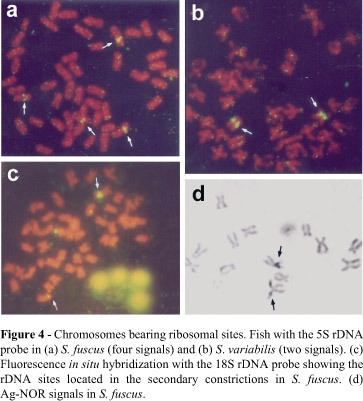
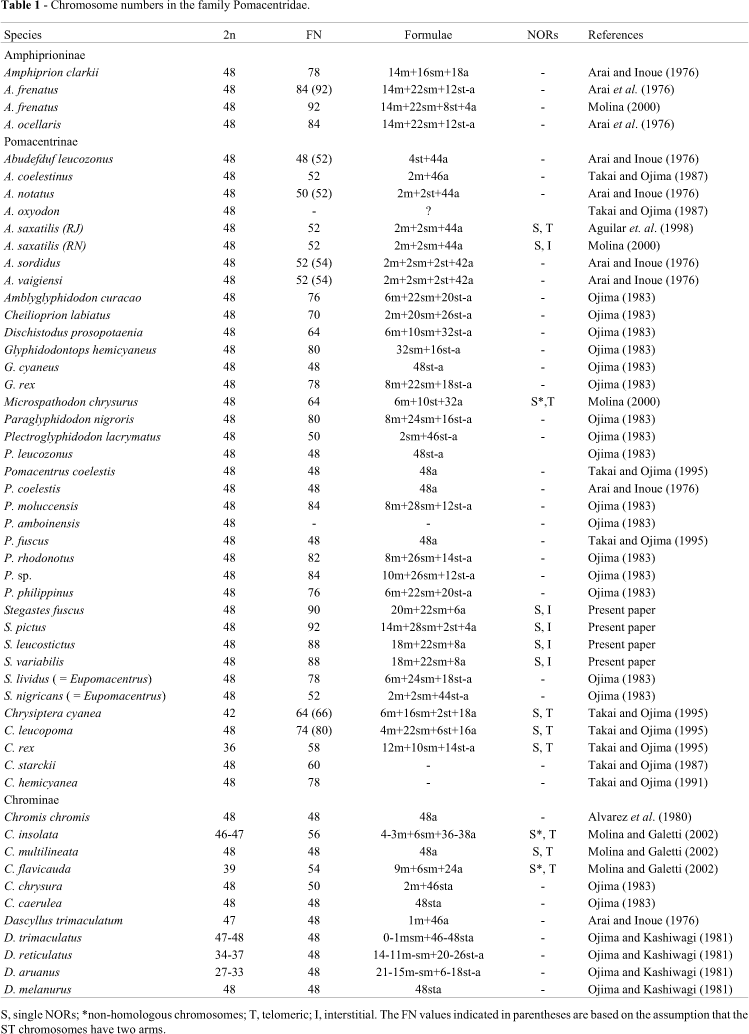
Stegastes fuscus (20m+22sm+6a), S. variabilis (18m+22sm+8a) (Figure 1), S. leucostictus (18m+22sm+8a), and S. pictus (14m+28sm+2st+4a) (Figure 2) had 2n = 48 and an elevated number of chromosomal arms (FN = 90, 88, 88 and 92, respectively). The heterochromatic regions were distributed in the centromeric and pericentromeric portions in most of the chromosomal pairs, with blocks that were more evident in the secondary constriction (Figures 1 and 2). NORs occurred at an interstitial position on the short arm of the largest submetacentric pair (pair 11 in S. fuscus, 10 in S. variabilis and S. leucostictus, and 12 in S. pictus) (Figure 3). In S. fuscus, hybridization with the 5S rDNA probe identified sequences located interstitially on the short arms of two unidentified chromosomal pairs, with one of these pairs having marks that were more evident than in the other. In contrast, signals were detected on only a single pair of chromosomes in S. variabilis (Figure 4). In S. fuscus, FISH using the 18S rDNA probe confirmed the sites identified by silver staining (Figure 4).
Discussion
The cytogenetic data currently available for marine Perciformes indicates a high degree of chromosomal conservation in which a large number of species show only minor deviations in the chromosomal organization and fundamental number. A karyotype with 48 chromosomes is considered ancestral for the Teleosts (Ohno, 1974), and occurs in 211 of the 660 Perciformes species analyzed so far (Klinkhardt et al., 1995).
Almost all of the species of Pomacentridae that have been analyzed cytogenetically have 2n = 48, with fundamental numbers that vary between 48 and 92 (Table I). A karyotype with 2n = 48 and FN = 48, considered ancestral in the group, has been observed in only a few species, such as Pomacentrus coelestis (Arai and Inoue, 1976), Chromis chromis (Alvarez et al., 1980) and C. multilineata (Molina and Galetti, 2002). Some representatives of the subfamily Chrominae show marked numerical polymorphism in their chromosomes while maintaining the number of chromosomal arms. This pattern suggests Robertsonian fusion in the karyotypic evolution of this group. Nevertheless, only 10% of the family has diploid values below the basal number. Despite the importance of polymorphism, this does not appear to be the main evolutionary tendency in the karyotype of this family, but may be characteristic of specific groups.
Even with a conserved chromosomal number there is considerable variation in the FN values in this family. This finding is a strong indication that pericentric inversions probably play an important role in the karyotypic diversification of this group of fish. Very little is known about structural aspects of the karyotype in the Pomacentridae. The first indications of C-banding in the Pomacentridae showed a pattern with reduced heterochromatic regions that was basically restricted to centromeric and pericentric areas. This pattern is common to most Perciformes analyzed so far (Galetti et al., 2000; Molina et al., 2002), and seems to define an array of characteristics that is common to karyotypes with little heterochromatin. A small content of heterochromatin appears to reduce or eliminate the dynamics provided by the heterochromatic segments in the karyotype of a given species. One of the most evident consequences of this small content of heterochromatin is the presence of single NORs in Perciformes and their limited distribution in the karyotype.
The occurrence of two chromosomal pairs with sites for the 5S rRNA gene in S. fuscus represents a derived condition in fish (Martins and Galetti, 1999). The presence of only one site in S. variabilis may reflect the fact that some sites are so small that they are incapable of emitting a detectable signal. Most probably, however, this single site represents an ancestral condition. Other species of this family have a single 5S rDNA locus, e.g. Abudefduf saxatilis, or two loci for the 5S gene, as in Chromis insolata and Chromis flavicauda (Molina and Galetti, 2002). In the latter two species, the 5S sequences are located in the pericentromeric region of two pairs involved in a Robertsonian fusion.
In contrast to other fish groups (Galetti et al., 1984), the NORs were not very informative cytotaxonomic markers in the Stegastes species examined here. FISH using an 18S rDNA probe confirmed the location of the major ribosomal sites identified by silver staining in S. fuscus. In this group, maintenance of the rDNA cluster on the same chromosomal pair among species reflected both the narrow phylogenetic proximity of the species and a lower dynamism of the internalized sites in relation to those located in telomeric positions.
Multiple pericentric inversions are an important mechanism of post-zygotic isolation (King, 1992). Events of this magnitude could favor speciation in extremely territorial coral species such as Stegastes. High fundamental numbers indicate that pericentric inversions in concert are widespread in the karyotypic evolution of the Pomacentrinae and Amphiprioninae. In both of these groups, biological characteristics that regulate the gene flow (territorialism, adhesive eggs and a sessile form) contribute to the break/split of the marine environment in defined micro-regions that may favor the fixation of chromosomal rearrangements.
The high FN values observed in S. variabilis (88), S. leucostictus (88), S. fuscus (90), and S. pictus (92) suggest the occurrence of rearrangements in chain acting upon the karyotypes of the species. The increase in the fundamental numbers seen here may be indicative of different levels of phylogenetic relationship among the species analyzed.
Acknowledgements
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Brazil.
Received: August 14, 2003; Accepted: June 3, 2004
Associate Editor: Fausto Foresti
- Aguilar CT and Galetti Jr. PM (1997) Chromosomal studies in South Atlantic serranids (Pisces, Perciformes). Cytobios 89:105-114.
- Alvarez MC, Cano J and Thode G (1980) DNA content and chromosome complement of Chromis chromis (Pomacentridae, Perciformes). Caryologia 33:267-274.
- Arai R and Inoue M (1976) Chromosomes of seven species of Pomacentridae and two species of Acanthuridae from Japan Bull Natn Sci Mus Ser A2, 73-78.
- Arai R, Inoue M and Ida H (1976) Chromosomes of four species of coral fishes from Japan. Bull Natn Sci Mus Ser A2, 137-141.
- Galetti Jr PM, Aguilar CT and Molina WF (2000) An overview on marine fish cytogenetics. Hydrobiologia 420:55-62.
- Galetti Jr PM, Foresti F, Bertollo LAC and Moreira FO (1984) Characterization of eight species of Anostomidae (Cypriniformes) fish on the basis of the nucleolar organizing region. Caryologia 37:401-406.
- Howell WM and Black A (1980) Controlled silver staining of nucleolus organizer regions with protective colloidal developer: 1- step method. Experientia 36:1014-1015.
- Hizume M (1994) Allodiploid nature of Allium wakegi Araki revealed by genomic in situ hybridization and localization of 5S and 18S rDNAs. Jpn J Genet 69:407-415.
- King M (1992) A dual level model for speciation by multiple pericentric inversions. Heredity 68:437-440.
- Klinkhardt M, Tesche M and Greven H (1995) Database of Fish Chromosomes. Westarp Wissenschaften, Magdeburg.
- Martins C and Galetti Jr PM (1999) Chromosomal localization of 5S rDNA genes in Leporinus fish (Anostomidae, Characiformes). Chromosome Res 7:363-367.
- Molina WF (2000) Análise da diversidade genética em Pomacentridae (Perciformes), através do uso combinado da citogenética, marcadores moleculares e morfometria multivariada. PhD Thesis, Universidade Federal de São Carlos, São Carlos.
- Molina WF, Affonso PRAM and Maia-Lima FA (2002) Divergence between karyotypical pattern and speciation events in Serranidae fish (Perciformes). Caryologia 55:299-305.
- Molina WF and Galetti Jr PM (2002) Robertsonian rearrangements in the reef fish Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genet Mol Biol 25:373-377.
- Nelson JS (1994) Fishes of the World. 3rd edition. John Wiley & Sons Inc., New York.
- Ohno s (1974) Animal Cytogenetics. Chordata 1 Protochordata, Cyclostomata and Pisces. v. 4. Gebrüder Burntraeger, Berlin.
- Ojima Y (1983) Fish cytogenetics. In: Sharma AK and Sharma A (eds) Chromosomes in Evolution of Eukaryotic Groups. v. 1. CRC Press, Boca Raton, pp 111-145.
- Ojima Y and Kashiwagi E (1981) Chromosomal evolution associated with Robertsonian fusion in the genus Dascyllus (Chrominae, Pisces). Proc Jap Acad 57B:368-70.
- Sumner AT (1972) A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res 75:304-306.
- Takai A and Ojima Y (1987) Comparative studies of karyotypes and distribution of nucleolus organizer regions in pomacentrid fish. 1. Proc Jap Acad 63b:17-20.
- Takai A and Ojima Y (1991) Comparative studies of karyotypes and distribution of nucleolus organizer regions in pomacentrid fish. 2. Cytobios 65:199-205.
- Takai A and Ojima Y (1995) Chromosome evolution associated with Robertsonian rearrangements in pomacentrid fish (Perciformes). Cytobios 84:103-110.
- White TJ, Bruns T, Lee S and Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ and White TJ. (eds) PCR Protocols: A Guide to Methods and Applications. Part 3. Academic Press, New York, pp 315-322.
Publication Dates
-
Publication in this collection
14 Jan 2005 -
Date of issue
2004
History
-
Received
14 Aug 2003 -
Accepted
03 June 2004