Abstract
Rice (Oryza sativa L.) breeding seeking to combine high productivity and cold tolerance for the temperate Latin America region is an important challenge. We estimated some useful parameters which can be used to investigate the genetic control of agronomic characters in crosses combining cold tolerance and productivity. A partial diallel design was used in crosses between six tropical indica rice cold susceptible genotypes (group 1) and seven japonica or indica/japonica cold tolerant rice genotypes (group 2). Parents and crosses were evaluated for agronomic characters under field conditions in two different experiments in 2005. The results showed significant mid-parent heterosis for all characters (plant height, tiller number, days to 50% flowering, panicle length, grains per panicle, sterility, and one-hundred grain weight). The predominant direction of dominance effects was negative for days to 50% flowering, and positive for all the other characters. General combining ability (GCA) and specific combining ability (SCA) were significant for all characters, although the GCA effects of the two groups were more important than the SCA effects.
general combining ability; heterosis; hybrid rice; specific combining ability
PLANT GENETICS
RESEARCH ARTICLE
Partial diallel analysis of agronomic characters in rice (Oryza sativa L.)
Edgar Alonso TorresI; Isaias O. GeraldiII
IFundo Latinoamericano de Arroz Irrigado, Centro Internacional de Agricultura Tropical, Cali, Colômbia
IIDepartamento de Genética, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba, SP, Brazil
Send correspondence to Send correspondence to: I.O.Geraldi Departamento de Genética Escola Superior de Agricultura "Luiz de Queiroz" Universidade de São Paulo, Caixa Postal 83 13400-970 Piracicaba, SP, Brazil E-mail: iogerald@esalq.usp.br
ABSTRACT
Rice (Oryza sativa L.) breeding seeking to combine high productivity and cold tolerance for the temperate Latin America region is an important challenge. We estimated some useful parameters which can be used to investigate the genetic control of agronomic characters in crosses combining cold tolerance and productivity. A partial diallel design was used in crosses between six tropical indica rice cold susceptible genotypes (group 1) and seven japonica or indica/japonica cold tolerant rice genotypes (group 2). Parents and crosses were evaluated for agronomic characters under field conditions in two different experiments in 2005. The results showed significant mid-parent heterosis for all characters (plant height, tiller number, days to 50% flowering, panicle length, grains per panicle, sterility, and one-hundred grain weight). The predominant direction of dominance effects was negative for days to 50% flowering, and positive for all the other characters. General combining ability (GCA) and specific combining ability (SCA) were significant for all characters, although the GCA effects of the two groups were more important than the SCA effects.
Key words: general combining ability, heterosis, hybrid rice, specific combining ability.
Introduction
New rice (Oryza sativa L.) varieties for temperate Southern America (southern Brazil, Argentina, Uruguay and Chile) with high yield potential and cold tolerance are an important breeding challenge. This region has a considerable area under actual and potential rice cultivation. In order to increase rice yield two complementary strategies have been followed, the introduction of high yielding tropical germplasm into a temperate background and early planting dates (late September and October) to capture high solar radiation in critical growth stages and to avoid low temperature stress after flowering.
The combination of cold tolerance and high yielding ability is possible in indica by japonica crosses (Peter Jennings, personal communication). The Japonica subspecies represents the majority of cultivars grown in the temperate regions and is cold tolerant in seedling, vegetative and reproductive stages (Mackill and Lei, 1997). Japonicas generally have poor plant type, disease susceptibility and are low yielding. In contrast, tropical elite indica varieties have good plant type, lodging and disease resistance and better yielding ability but are cold susceptible and have a long growth duration in temperate regions.
Indica by japonica crosses present problems. Many studies show that rice yield related characters (tiller number, grain number and grain weight) and agronomic characters (plant height and days to flowering) are inherited quantitatively, related genetically to one another and influenced by growing environments (Kobayashi et al., 2003). Furthermore, hybrid sterility caused by several genetic mechanism, limits the use of indica/japonica crosses (Ikeshashi and Araki, 1986). Other constraints could be the presence of blocks of genes for adaptation to specific environmental conditions segregating as one super gene that limited free recombination.
Diallel mating designs are an important tool in plant breeding programs to obtain information on the inheritance of quantitative traits. A partial diallel consisting of crosses between two parents groups is useful when there are distinct groups of populations and the breeder is not interested in evaluating the crosses between parents of the same group (Miranda Filho and Geraldi, 1984). Essentially this mating system corresponds to Design II proposed by Comstock and Robinson (1952), with fixed parents. In order to diminish the number of crosses to be used, Miranda Filho and Geraldi (1984) and Geraldi and Miranda Filho (1988) presented the models, estimation formulae and tests for effects and for obtaining the sum of squares in the analysis of variance of the partial diallel by adapting the Gardner and Eberhart (1966) and Griffing (1956) methods.
Despite the advances of "molecular breeding", classical quantitative genetics remains useful in practical rice improvement. Thus, estimation of genetic parameters such as heterosis and combing ability gives inferences about the predominant action of the genes, indicates the appropriate selection strategy to be applied in the breeding program and allows the identification of the best parents. However, in Latin America little is known about the expression or inheritance of agronomic characters in crosses designed to combine yield potential and low temperature tolerance in rice. This study used partial diallel analysis to determine the combining ability of tropical indica cold susceptible and japonica or indica/japonica cold tolerant rice lines and to study the genetic basis of important agronomic and grain yield related traits.
Material and Methods
The material used in this study involved six tropical cold susceptible indica parents (L3000, Oryzica 1, Irga 417, BR-Irga 410, FL03188 and Epagri 108) and seven japonica or indica-japonica parents (Quilla 145601, Quilla 173201, CT6748-8-CA-17P, L201, FL04423-CA-2P, FL04402-CA-4P and FL04452-CA-14P). The susceptible parents, designated group 1, were commercial varieties from Uruguay, Rio Grande do Sul and Santa Catarina in Southern Brazil, and Colombia. The tolerant parents, designated group 2, were composed of genotypes from Chile, California, and lines produced by temperate breeding programs from the International Center for Tropical Agriculture (CIAT) and the Latin American Fund for Irrigated Rice (FLAR). The seed of this material was kindly donated by Luis Berrio from the FLAR Germplasm Bank. This material was crossed according to a partial diallel mating scheme (Miranda Filho and Geraldi, 1984) in 2004 and 2005 at CIAT, Colombia, by using a modified method of rice crossing (Sarkarung, 1991).
Parent lines and F1 hybrids were evaluated in the planting dates of December 2004 and July 2005, for agronomic characters under field conditions in CIAT, Palmira, Colombia, 3°30' N, 75°22' W, 965 m above sea level, in a typical Pellustert soil. The temperature varied from 19.8 °C to 30.4 °C (mean: 25.1 °C) in the first experiment and from 19.3 °C to 30.6 °C (mean: 25.0 °C) in the second experiment. Each experiment was conducted using a randomized complete block design with four replications. Entries were grown in single row plots (5 m x 0.3 m) and 17 plants were transplanted in each row. Flood irrigation, fertilizer (200 N, 60 P2O5, 60 K2O, and 30 Zn, kg ha-1) and chemical weed control were used. The measured agronomic characters were: days to 50% flowering (DF), tiller number (TN), plant height (PH), panicle length (PL), number of total grains per panicle (GN), percent sterility (PE) and one thousand grain weight (GW). Grain yield was not recorded due to the high sterility in the indica/japonica crosses.
Individual and joint analyses of variance were performed by the GLM procedure of the SAS program (SAS Institute, 1999) according to the randomized complete block design, considering experiments and genotypes as fixed effects. Genotypes sums of squares were partitioned as parents, crosses and parent versus (vs.) crosses. Additionally, parents were partitioned as susceptible, tolerant and susceptible vs tolerant. Similarly, the interaction sum of squares were partitioned as experiment by parents, experiment by hybrids and experiment by parents versus hybrids, experiment by parents group 1, experiment by parents group 2 and experiment by group 1 versus group 2. Genetic analysis to estimate general combining ability (GCA), specific combining ability (SCA) and their interaction effect with location, was performed using the Griffing (1956) method 4, model I, adapted for a partial diallel by the methods of Geraldi and Miranda Filho (1988) and Oliveira et al. (1987), according to the following model:

In this model Yijkl is the observed value relative of the ijth hybrid at the kth block of the lth experiment; µ is the population mean; pl is the planting date or experiment effect (l = 1, 2); bk/l is the block or replication effect within experiments (k = 1, 2, ..., 4); gi is the GCA effect for the ith parent of group 1 (i = 1, 2, ..., 6); gj is the GCA effect for the jth parent of group 2 (j = 1, 2, ..., 7); sij is the SCA effect for ijth F1 hybrid (ij = 1, 2, ..., 42); gpil is the interaction between group 1 GCA effects and experiments, gpjl is the interaction between group 2 GCA effects and experiments; spijl is the interaction between SCA effects and experiments; and eijkl is the experimental error.
Because experiments and genotypes were considered as fixed effects, and blocks within experiments and error as random effects, the source blocks within experiments was used to test the experiment effect, and the pooled error was used to test all other effects in the joint analysis of variance. In the diallel analysis, group 1 general combining ability (gi), group 2 general combining ability (gj), specific combining ability (sij) and the interactions of these effects with the experiments (gpil, gpjl, and spijl) were tested with the pooled error considering only hybrids evaluation. Average mid-parent heterosis was tested for significance using F-test in the contrast parents vs. crosses in the analysis of variance. Data were analyzed by a SAS program using the GLM procedure and the "CONTRAST" and "ESTIMATE" statements (Zhang and Kang, 1997; Wu and Matheson, 2000), in order to test hypothesis, estimate effects and standard errors. In the design matrix was used the following restrictions:  for each j and
for each j and  , for each i (Geraldi and Miranda Filho, 1988). The quadratic components associated with the GCA of the two groups and the SCA effects were estimated by the method of moments based on mean square expectation according to the formulas:
, for each i (Geraldi and Miranda Filho, 1988). The quadratic components associated with the GCA of the two groups and the SCA effects were estimated by the method of moments based on mean square expectation according to the formulas:

where I is the number of parents of group 1, J is the number of parents of group 2, K is the number of replications, L is the number of experiments, MSGCAg1 and MSGCAg2 are the mean squares of GCA effects of groups 1 and 2 respectively, MSSCA is the mean square of SCA effects, and MSE is the error mean square. The relative importance of general and specific combining ability in determining progeny performance was assessed by the ratio  (Baker, 1978).
(Baker, 1978).
Results
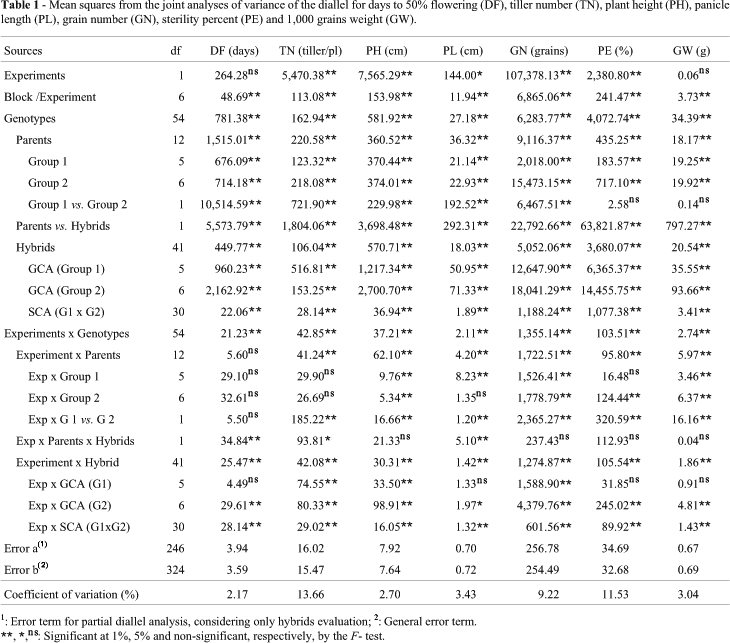
A joint analysis of variance was performed to determine the importance of the experiments, genotypes and their interactions (Table 1). Significant experiment effects were detected for tiller number, plant height, panicle length, grain number and sterility probably due to light and temperature variations between the planting dates. Genotype effects and experiment by genotype interaction were highly significant for all characters.
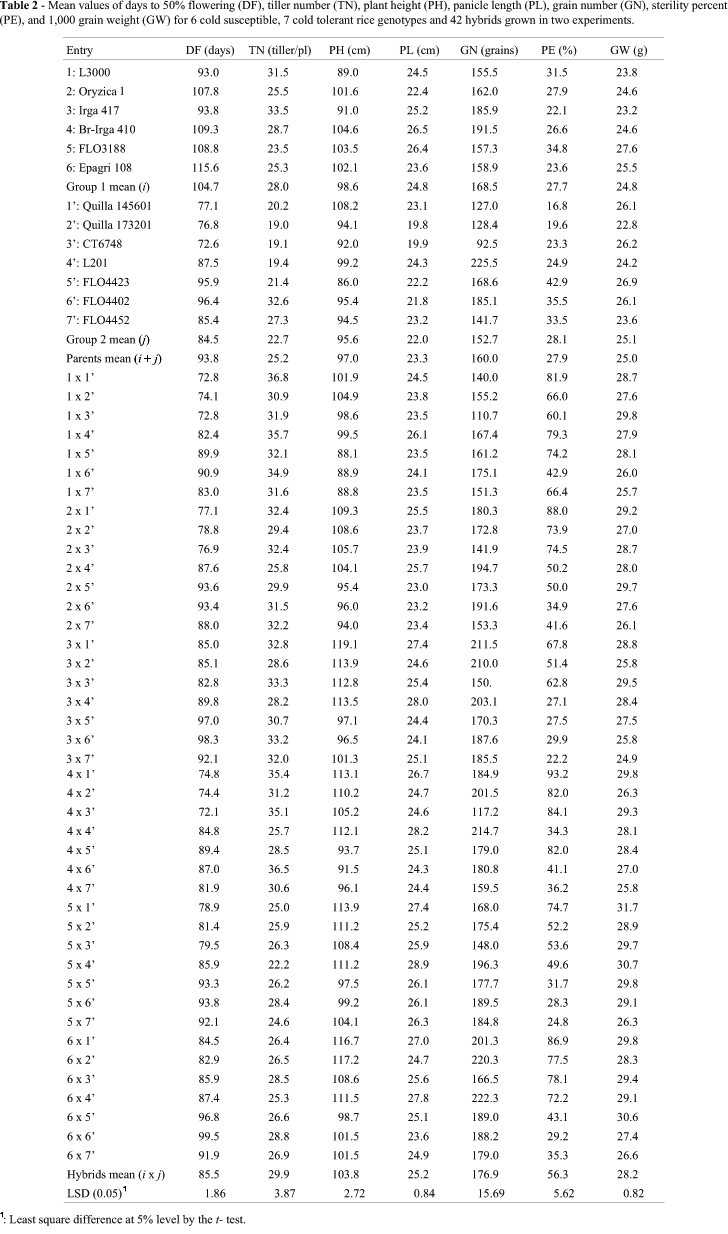
The two groups of parents were very different between and within them. The Group 1 and Group 2 contrast was highly significant for DF, TN, PH, PL and GN, indicating genotypic differences between the groups. In general, Group 2 parents were earlier, with lower tiller number and plant height, short panicles, and fewer grains by panicle. They were similar to Group 1 parents in sterility and 1,000 grain weight (Table 2). In group 1, L3000 and Irga 417 were earlier, shorter, and higher tillering; BR-Irga 410 had long panicles with more grains and FL03188 was more sterile with heavier grains. In group 2, typically japonica parents Quilla 145601, Quilla 173201, CT6748 and L201 were earlier and lower tillering; Quilla 145601 was the tallest parent; L201 had long panicles with more grains; the anther culture derivates FLAR lines (FLO4423, FLO4402 and FLO4452) were more sterile and FL04423 line had heavier grain weight (Table 2).
The contrast of parent vs. hybrid was highly significant for all characters, indicating the presence of mid-parent heterosis. In general the hybrids were earlier, with higher tiller number, taller, and had longer panicles, more grains per panicle, more sterility, and heavier grains than parents (Table 2). These results show the presence of genetic divergence between groups in the loci with dominance and directional dominance for all characters. The comparison between parent and hybrid means of all combinations showed the predominant direction of dominance in the groups of parents that were negative for days to flowering and positive for all other characters.
There was interaction between experiment and parents for all characters with the exception of days to flowering. This interaction was significant for group 1 parents for PH, PL, GN, and GW; and for group 2 parents for the PH, GN, PE and GW characters, suggesting that environmental conditions affected some characters in different ways in the two groups of parents. The contrast group 1 vs. group 2, interacted with experiments for all characters with exception of days to flowering. This interaction was complex (rank change) only for PE and GW.
The interaction of the experiment with the contrast parents vs. hybrids was non significant for PH, GN, PE and GW. The interactions with DF, TN, and PL were significant but non-complex, indicating that the general superiority of the hybrids over parents was consistent in the two experiments.
When cold susceptible and cold tolerant rice genotypes were crossed, considerable genetic variability was produced. Highly significant differences among hybrids were found for all characters (Table 1), indicating the existence of considerable genotypic variability among them. The diallel analysis indicated significance at the 1% level for the GCA group 1, GCA group 2 and SCA effects for all characters over experiments. These results indicate the existence of differences in the breeding values between parents of two groups and the presence of additive and non-additive gene effects for all characters in the study.
Estimates of the quadratic components associated with GCA and SCA effects are given in Table 3. An estimate of the relative importance of the additive and non-additive effects of genes is obtained from the ratio of the components for general combining ability of the two groups and the specific combining ability according to the ratio  (Baker, 1978). The values obtained show that for all characters, this proportion was higher than 0.75, indicating that additive effects are more important than non-additive effects for the characters studied.
(Baker, 1978). The values obtained show that for all characters, this proportion was higher than 0.75, indicating that additive effects are more important than non-additive effects for the characters studied.
Experiment by hybrid interaction was highly significant for all characters, indicating that the crosses behaved differently according to the experimental conditions. Similarly, interactions between experiments and GCA of group 2 and SCA were highly significant for all characters, although interaction with the GCA of group 1 was significant only for TN, PH, and GN, indicating that some estimates could change according to the planting date. However, when the sums of squares of these interaction components were compared with the total sums of squares the proportion was lower than 6%, indicating that it was of little importance compared with the main effects.
The GCA effect, which determines the average performance of a parent in crosses, and is an estimate of its breeding value was determined for each parent (Table 4). For days to flowering several parents adapted to sub-tropical conditions had negative significant effects (reduced days to 50% flowering in their hybrids) such as L3000 and BR-Irga 410 of group 1, and Quilla 145601, Quilla 173201 and CT6748 of group 2, so these parents could be useful donors for an early flowering trait. For tiller number, significant positive effects were observed for L3000, Irga 417 and BR-Irga 410 group 1 parents as well as for FL04402 and Quilla 145601 group 2 parents. Two group 1 parents (L3000 and BR-Irga 410) with positive effects for tiller number had significant effects for plant height (dwarfness), while in group 2 anther culture FLAR temperate program derived lines FL04423, FLO4402 and FL04452 showed negative effects for plant height. Lines FL03188, Irga 417, Epagri-108 and BR-Irga 410 were the best group 1 parents for the long panicle trait. Similarly, in the second group, L201 from California and Quilla 145601 had highly significant and positive effects for the long panicle trait. For number of grains per panicle, Epagri 108 (group 1) showed the highest positive effect and L201 (group 2) had one similar effect. Lines with certain introgression from the other background group were more compatible in inter-group crossing. Examples are Irga 417 and FL03188 of group 1, and L201, FL04423, FL04402, and FL04452 of group 2. Finally, for 1,000 grain weight the best parents were FL03188 from group 1 and Quilla 145601 from group 2.
The usefulness of a particular cross in exploiting heterosis is judged by specific combining ability effects. These values are show in Table 5. In the model followed, these effects indicate the interaction between specific alleles of parents involved in this cross. Combinations in the desired directions are useful in a pure line program, because of transgressive segregation in advanced generations. Examples include: 6x4' (Epagri 108 x L201) for DF, 1x4' (L3000 x L201) for TN, 4x6' (Br-Irga 410 x FLO4402) for PH, 1x6' (L3000 x FL04402) for PL, 3x1' (Irga 417 x Quilla 145601) for GN, 4x4' (Br-Irga 410 x L201) for PE, and 3x3' (Irga 417 x CT6748) and 6x5' (Epagri 108 x FL04423) for GW. However, no cross combined all values in a desirable direction, indicating the necessity of previous breeding of parents to combine all desirable SCA effects in a single hybrid. Besides, the trait sterility percent (PE) deserves special attention, since most of the hybrids presented a high degree of sterility, as indicated by the hybrid means and also by the components of means. These facts indicate the importance of choosing parents with wide compatibility in order to make the heterosis in rice useful.
Discussion
This study encountered considerable sterility in crosses between cold susceptible and cold tolerant rice genotypes. The hybrid mean for sterility was 56.3% with variation from 24.8% to 93.0%. These results coincide with the sterility typical in indica by japonica crosses (Oka and Morishima, 1997). Interestingly, some parents with introgression from either indica or japonica, such as Irga 417, FL03188, FL04423, FL04402 and FL04452 were generally less sterile in their inter-group crosses, showing more compatibility. Virmani et al. (2003) pointed out that indica and japonica crosses result in hybrid sterility despite their high heterosis for various agronomic characters but there are some crosses with normal spikelet fertility, in this case one or both parents posses genes for wide compatibility. These findings suggest that there is Latin-American rice germplasm having the wide compatibility character, useful for wide hybridizations and hybrid rice production.
Significant mid-parent heterosis was found for all characters in crosses between cold susceptible and cold tolerant rice lines. These results agree with the literature showing the superiority of rice hybrids in indica/japonica crosses. Virmani (1989) reported estimates of mid-parent heterosis for several agronomic characters based on an extensive review. Recently, Yuan (2003) mentioned that rice heterosis follow a general trend and was highest in indica/japonica crosses. Because heterosis indicates dominance, supplying superior rice hybrids to farmers is a valuable strategy to exploit the non-additive portion of the genetic variance. As allelic dispersion also contributes to heterosis, study of it in relation to rice heterosis and inbreeding depression could provide new ways for rice improvement.
In our present study, genetic analysis by the Griffing IV method adapted for a partial diallel revealed the presence of additive and non-additive effects of the genes in the inheritance of agronomic characters in rice. Calculation of the relative importance of the additive and non-additive effects showed that the relative proportion of the sum of quadratic components associated with the additive effects was higher than 0.75 for all characters. This suggests that in the crosses studied additive effects were more important than non-additive effects in the inheritance of days to 50% flowering, tiller number, plant height, panicle length, grains per panicle, sterility percent and grain weight. Similar findings were reported by Yonezawa (1997) who recognized the existence of some dominance effects while concluding that the major component of genotypic variance in rice is additive. Gravois and McNew (1993) using USA germplasm and a factorial design showed that for yield, plant height, panicle number, panicle length and number of primary branches in the panicle the GCA effects were more important than the SCA effects as they explained 70%, 89%, 60% and 85% of the genotypic variance. Singh and Kumar (2004) in a line x tester analysis found the prevalence of additive variance for days to flowering, plant height, effective tillers, and grain yield, although line by tester contrast was also significant, indicating the importance of additive and non-additive effects. Verma and Srivastava (2004) also founded that CGA and SCA effects were highly significant for days to 50% flowering, plant height, number of productive tiller, panicle length, number of spikelets per panicle and 100 grain weight, indicating the relevance of both additive and non-additive effects in the inheritance of these traits as well as the greater importance of non-additive gene action.
The variation observed in the breeding values of the two groups of parents show that in Latin American irrigated rice germplasm there exist considerable additive genetic variation to be exploited in breeding to produce inbred lines for sub-tropical conditions. Because of the importance of additive effects, the dispersion of positive alleles for all characters within and between groups, and the difficulty in using immediately exotic germplasm, breeding methods such as recurrent selection that emphasize GCA and is an efficient method of systematically recombining many genes could be used in rice breeding programs in order to combine high yield potential and cold tolerance for use in temperate Southern America regions.
These experiments were conducted in a median altitude tropical location and the performance of parents and hybrids could change in other environments. In fact, the occurrence of genotype by environment interactions indicates that the performance of the genotypes could differ according to the environmental conditions. However, the temperatures observed in this study are not very different from the summer temperatures in the southern cone. Furthermore, many important varieties such as Irga 409, BR-Irga 410 and El Paso 144 have been obtained by local selection from segregant populations developed at CIAT. Thus, besides increasing information on the inheritance of traits related to cold tolerance in rice, these results will be useful for rice breeding programs in the southern cone.
Acknowledgments
We are grateful to the National Rice Foundation from Venezuela (FUNDARROZ) for the scholarship to EA Torres, to the Dr. PR Jennings for reading and correcting the text, to the Latin American Fund for Irrigated Rice (FLAR) and The International Center for Tropical Agriculture (CIAT) for grants and help to support this work and to FLAR employees for collaboration in field work.
Received: September 12, 2006; Accepted: November 29, 2006.
Senior Editor: Ernesto Paterniani
- Baker RJ (1978) Issues in diallel analysis. Crop Sci 18:533-536.
- Comstock RE and Robinson HF (1952) Estimation of average dominance of genes. In: Gowen JW (ed) Heterosis. Iowa State College Press, Ames, pp 494-516.
- Gardner CO and Eberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22:439-452.
- Geraldi IO and Miranda Filho JB (1988) Adapted models for the analysis of combining ability of varieties in partial diallel crosses. Braz J Genet 11:419-430.
- Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Aust J Biol Sci 9:463-493.
- Gravois K and McNew R (1993) Combining ability and heterosis in U.S southern long grain rice. Crop Sci 33:83-86.
- Ikehashi H and Araki H (1986) Genetics of F1 sterility in remote crosses of rice. In: Proceedings of the International Rice Genetics Symposium. The International Rice Research Institute (IRRI), Manila, pp 119-130.
- Kobayashi S, Fukuta Y, Sato T, Osaka M and Khush GS (2003) Molecular marker dissection of rice (Oryza sativa L.) plant architecture under temperate and tropical climates. Theor Appl Genet 107:1350-1356.
- Mackill D and Lei X (1997) Genetic variation for traits related to temperate adaptation of rice cultivars. Crop Sci 37:1340-1346.
- Miranda Filho JB and Geraldi IO (1984) An adapted model for the analysis of partial diallel crosses. Braz J Genet 7:677-688.
- Oka HK and Morishima H (1997) Wild and cultivated rice. In: Matsuo T, Fusuhara Y, Kikuchi F and Yamaguchi H (eds) The Science of the Rice Plant. V. 3: Genetics. Food and Agriculture Policy Research Center, Tokyo, pp 88-111.
- Oliveira AC, Morais AR, Souza Junior CL and Gomes EE (1987) Analysis of partial diallel crosses repeated over locations. Braz J Genet 10:517-533.
- Sarkarung SA (1991) Simplified Crossing Method for Rice Breeding: A Manual. Centro Internacional de Agricultura Tropical, Cali, 32 pp.
- SAS Institute (1999) SAS Language Guide for Personal Computers (Release 8.0 edition). SAS Inst., Cary.
- Singh NK and Kumar A (2004) Combining ability analysis to identify suitable parents for heterotic rice hybrid breeding. International Rice Research Notes 29:21-22.
- Verma OP and Srivastava HK (2004) Genetic component and combining ability analyses in relation to heterosis for yield and associated traits using three diverse rice growing ecosystems. Field Crops Res 88:91-102.
- Virmani S (1989) Heterosis and Hybrid Rice Breeding. Spring Verlag and IRRI, Manila, 189 pp (Monographs on Theor Appl Genet, 22).
- Virmani SS, Sun ZX, Mou TM, Ali AJ and Mao CX (2003) Two-line hybrid rice breeding manual. The International Rice Research Institute, Los Baños, 88 pp.
- Wu HX and Matheson AC (2000) Analysis of half diallel mating system with missing crosses: Theory and SAS program for testing and estimating GCA and SCA fixed effects. Silvae Genet 49:130-137.
- Yonezawa K (1997) Yield components. In: Matsuo T, Fusuhara Y, Kikuchi F and Yamaguchi H (eds) The Science of the Rice Plant. V. 3: Genetics. Food and Agriculture Policy Research Center, Tokyo, pp 400-412.
- Yuan LP (2003) Recent progress in breeding super hybrid rice in China. In: Virmani S, Mao C and Hardy B (eds) Hybrid Rice for Food Security Poverty Alleviation and Environmental Protection. The International Rice Research Institute, Manila, pp 3-6.
- Zhang Y and Kang MS (1997) DIALLEL-SAS: A SAS program for Griffing's diallel analyses. Agronomy J 89:176-182.
Send correspondence to:
Publication Dates
-
Publication in this collection
30 Aug 2007 -
Date of issue
2007
History
-
Received
12 Sept 2006 -
Accepted
29 Nov 2006