Abstract
The Dumbo rat possesses some characteristics that evoke several human syndromes, such as Treacher-Collins: shortness of the maxillary, zygomatic and mandibular bones, and low position of the ears. Knowing that many homeobox genes are candidates in craniofacial development, we investigated the involvement of the Msx1 and Dlx1 genes in the Dumbo phenotype with the aim of understanding their possible role in abnormal craniofacial morphogenesis and examining the possibility of using Dumbo rat as an experimental model for understanding abnormal craniofacial development. We studied the expression of these genes during craniofacial morphogenesis by RT-PCR method. We used Dumbo embryos at E12 and E14 and included the Wistar strain as a control. Semi-quantitative PCR analysis demonstrated that Msx1 and Dlx1 are expressed differently between Dumbo and Wistar rats, indicating that their low expression may underly the Dumbo phenotype.
Dumbo rat; Msx1; Dlx1; face; embryo; development
DEVELOPMENTAL GENETICS
SHORT COMMUNICATION
Expression of Msx1 and Dlx1 during Dumbo rat head development: correlation with morphological features
Suhair KaterjiI, Nathalie VanmuylderI; Michal SvobodaII; Marcel RoozeI; Stéphane LouryanI
ILaboratory of Anatomy and Embryology, Faculté de Médecine, Université Libre de Bruxelles, Brussels, Belgium
IILaboratory of Biological Chemistry, Faculté de Médecine, Université Libre de Bruxelles, Brussels, Belgium
Send correspondence to Send correspondence to: Stephane Louryan Laboratory of Anatomy Biomechanics and Organogenesis Faculté de Médecine Université Libre de Bruxelles route de Lennik 808 B 1070, Brussels E-mail: slouryan@ulb.ac.be
ABSTRACT
The Dumbo rat possesses some characteristics that evoke several human syndromes, such as Treacher-Collins: shortness of the maxillary, zygomatic and mandibular bones, and low position of the ears. Knowing that many homeobox genes are candidates in craniofacial development, we investigated the involvement of the Msx1 and Dlx1 genes in the Dumbo phenotype with the aim of understanding their possible role in abnormal craniofacial morphogenesis and examining the possibility of using Dumbo rat as an experimental model for understanding abnormal craniofacial development. We studied the expression of these genes during craniofacial morphogenesis by RT-PCR method. We used Dumbo embryos at E12 and E14 and included the Wistar strain as a control. Semi-quantitative PCR analysis demonstrated that Msx1 and Dlx1 are expressed differently between Dumbo and Wistar rats, indicating that their low expression may underly the Dumbo phenotype.
Key words: Dumbo rat, Msx1, Dlx1, face, embryo, development.
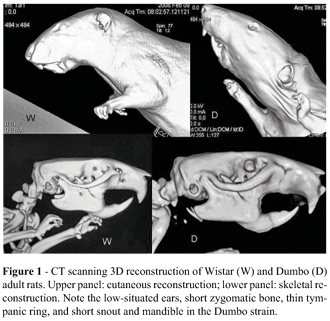
The "congenitally malformed" Dumbo rats seem to be the product of domestic breeding of rats of Wistar origin, probably in the USA, a few decades ago. They evoke comparisons with some human malformation syndromes, such as the Treacher-Collins, DiGeorge, and Nager syndromes, because of micrognathia, low position of the ears, and hypoplasia of the zygomatic, maxillary and mandibular bones (Figure1). This strain may constitute an experimental model for understanding abnormal craniofacial development.
Preliminary morphological and morphometic analysis indicated that the considerable differences between the craniofacial structures of Dumbo and Wistar rats might be due to genetic mutations in the Dumbo rat that were undetectable by chromosome mapping. Furthermore, the embryonic skulls of Dumbo rats displayed a delay bone growth. For these reasons, we selected the embryonic ages E12 and E14 (initiation of the chondrogenesis, beginning of ossification) for the present study.
Analysis of the embryonic development of Dumbo rats shows quantitative defects in structures derived from the first pharyngeal arch. These developmental defects are represented by disturbances in chondrogenesis and osteogenesis pathways, suggesting the involvement of the Msx1 and Dlx1 genes.
To compare genetic expression in Dumbo rats with the normal Wistar strain, we used RT-PCR to estimate the expression of Msx1 and Dlx1. As loading controls we used the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH), which is expressed at a constant level in different tissues, cells or experimental treatments (de Jonge et al., 2007). Since the use of multiple internal control genes has been recommended (Vandesompele et al., 2002), we used the nerve growth factor (NGF) encoding gene as second reference gene. NGF appears to be ubiquitously expressed in some craniofacial primordia during mouse development (Louryan et al., 1995).
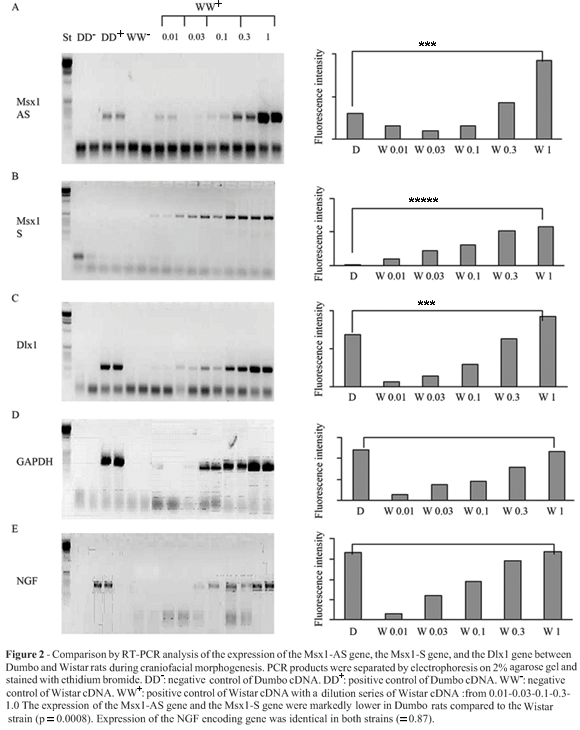
For the analyses on Dumbo rats, nine embryos at stages E12 and E14 were obtained from three different mothers, respectively. We also collected the same number of embryos for Wistar rats. Total RNA was extracted from small amounts of head tissue (20-100 mg) using the RNA NOW TC method (Texagen), according to the manufacturer's directions. The RNA pellet was dissolved in 50 µL of DEPC-treated water and RNA concentration was determined by spectrophotometry at 260 nm/ 280 nm using a Nanodrop ND1000 apparatus (Isogen). cDNA was synthesized by the Gene Amp RNA PCR kit (Applied Bio systems) using the enzyme MultiScibeRT (50U/µL): 1-2 µg of total RNA were transcribed in 20 µL of final volume of manufacture's buffer enriched with 2.5 µM random hexamers, 1 mM of dNTP and 20 units of RNAse inhibitor. Samples were incubated first at 25 °C for 10 min, then at 37 °C for 120 min. The reaction was stopped by the adition of 20 µL of 0.1 M EDTA and 30 µL water. The samples were heated to 94 °C for 2 min before storing at -20 °C. PCR reactions were set up in 20 µL, using the GoTaq PCR kit (Promega) with "Green buffer" and a final concentration of 200 µM dNTP, and 10 nM of each primer with 0.5 units of GoTaq DNA polymerase. Thermocycling wasperformed in MyCycler (BioRad), starting with a denaturation for 2 min at 94 °C, followed by cycles of 10 s at 94 °C, 20 s at 55-60 °C (depending on pair of primer used), 1 min at 72 °C. If not indicated otherwise, 35 amplifications cycles were done. After amplification, electrophoresis of 10 µL of each PCR product was performed on a 2% agarose gel with 0.5 µg/mL ethidium bromide, fragment size was estimated from a using 1 kb DNA ladder (Promega). To control for contamination of samples with genomic DNA, all PCR amplifications were carried out in parallel with a negative control of reverse transcription, i.e. with RNA samples submitted to reverse transcription but without MultiScibe Reverse Transcriptase. Semiquantitative RT-PCR estimates were validated using a standard curve dilution series of Wistar rat cDNA. Densitometries of amplicon fluorescence intensity were performed using VilberLourmat Bio1D software.
The RT-PCR analyses revealed that the expression of the Msx1 sense (S) gene, the Msx1 antisense (AS) gene and of the Dlx1 gene in the craniofacial region of E12 and E14 embryos was markedly lower in Dumbo rats than in Wistar rats (Figure 2). A very large difference was observed for the Msx1sense (S) gene, which was almost undetectable in Dumbo rats. Using dilution curves of Wistar cDNA, we validated that in our conditions the fluorescence intensity of amplicons was directly related to the initial concentration of target DNA. Using dilutions curves, we estimated that the expression of the Msx1 sense (S) gene in the Dumbo rat was one hundred times lower than in the Wistar rat. The difference between Dumbo and Wistar rats is significant (p = 0.0008). Expression of the Msx1antisense gene and of the Dlx1 gene in the Dumbo rat were roughly threefold lower than in the Wistar rat. The differences between Dumbo and Wistar rats were significant (p = 0.0008).As expected, the two rat strains did not differ significantly in the expression of the control genes: for GAPDH gene (p = 1.00) and for NGF gene (p = 0.87).
During embryogenesis, cranial neural crest cells migrate into the presumptive mandibular, maxillary and zygomatic primordia, where they condense to form mesenchymal and precartilaginous blastemata before differentiating into osteoblasts. The osteoblasts synthesize bone matrix through intramembranous ossification (Couly et al., 1993; Hall and Miyake, 2000), whereas the ossicles of ear, derived from Meckel's and Reichert's cartilage form through endochondral ossification. Normal development requires mechanisms to ensure that bone morphology and growth are matched to those of the developing skull (Morriss-Kay, 1993).
The generation of different cell types from cranial neural crest (CNC) is regulated by genetic control, which is beginning to be elucidated, as a large number of candidate regulatory genes identified and mutations in these genes are being made. Some of these genes are homeobox genes. They are expressed in the early phases of development in a spatially and temporally restricted manner and have been implicated in the specification of particular domains of the head. Homeobox genes are a conserved ubiquitous superfamily of transcription factors found in all eukaryotes, with analogs in prokaryotes. In eukaryotic organisms, these genes generally regulate axis determination, segmental patterning, and tissue identity during development. The protein product of a homeobox gene contains a highly conserved homeodomain at the carboxyl end that includes a DNA binding helix-turn-helix motif. Homeobox proteins also contain a variable region composed of one or more domains involved in protein binding specificity and regulation (Qian et al., 1989; Kissinger et al., 1990)
The Msx and Dlx homeobox gene families are expressed in the pharyngeal arches, giving rise to craniofacial structures. The mandible, maxilla, zygoma and ear are derived from the first pharyngeal arch, which receives neural crest cells from the midbrain, namely the mesencephalon, and rhombomeres r1 and r2 (Tan and Morriss-Kay, 1985). The expression of the Msx and Dlx gene families in the cranial neural crest cells emigrating from the central nervous system continues in the craniofacial regions.
The Msx1 gene is strongly expressed in CNC (cranial neural crest) cells and plays a critical role in regulating epithelial-mesenchymal transitions during morphogenesis (Robert et al., 1989). Expression of Msx1 in the cranial neural crest continues during cell migration and colonization of the pharyngeal arches (Mackenzie et al., 1992). In the central nervous system, the expression of Msx1 is essential in the hindbrain and the rhombomeres. Dorsolateral expression of Msx1 continues in the brain during neurulation and becomes more lateral (Bendall and Abate-Shen, 2000).
Dlx1 is a member of the distal-less homeobox gene family. It is likely to be responsible for programming developmental events along the proximodistal and mediolateral dimensions of the pharyngeal arches (Qiu et al., 1995). Dlx genes establish intra-arch identity (Depew et al., 2005). Since the CNC contributing to the maxillary and mandibular components of the first arch is derived from the posterior midbrain and rhombomeres 1 and 2 (Osumi-Yamashita et al., 1994), candidate regulators of the Dlx genes must be expressed in this neuroepithelium.
Our findings may help to explain the delayed chondrogenesis and the late osseous growth of these regions in Dumbo rats in comparison with Wistar rats. Blin-Wakkach et al., (2001) demonstrated the existence of endogenous Msx1 antisense RNA (Msx1-AS RNA) in differentiated dental and craniofacial tissues of mice, rats, and humans. They also showed that this AS RNA can block Msx1 protein expression and that it exhibits a reverse temporospatial distribution pattern with Msx1 protein both in vivo and in vitro.
Msx1-S is expressed strongly in the proliferative progenitor cells of dental mesenchyme and bone, and it is down-regulated in terminally differentiated tissues (Robert et al., 1989; Mackenzie et al., 1991; Houzelstein et al., 1997). By contrast, an inverse distribution of the Msx1-AS RNA was shown by Blin-Wakkach et al. (2001). These authors showed that when the AS transcript is more abundant, Msx1 protein is undetectable, and conversely, overexpression of the sense RNA results in production of Msx1 protein. They next demonstrated that the balance between the levels of the two Msx1 RNAs (sense/antisense) is related to the expression of Msx1 protein and that this ratio is very important in the control of terminal differentiation of the skeleton. They also demonstrated that the Msx1-AS RNA is involved in a cross talk between the Msx-Dlx pathways. Forest-Potts and Sadler (1997) highlighted that antisense attenuation of Msx1 during early stages of neurulation led to hypoplasia of the maxillary and mandibular bones, and to abnormalities in the neural tube. When cultured mouse embryos were injected with Msx1-AS oligodeoxynucleotides, expression of Msx1 protein was disrupted and craniofacial abnormalities ensued. Msx1 was shown to down-regulate the master gene of osteoblastic determination, Cbfa1, a strongly indication that the ratio between Msx1-S and Msx1-AS RNA is a key factor in cell differentiation and phenotypic expression in mineralized tissues (Blin-Wakkach et al., 2001). Because the expression patterns of the Msx genes are closely related to the development of neural crest cells in several species, the failure of early craniofacial development could be due to aberrant CNC cells induction or migration. Han et al., (2007) reported that the Msx1 gene is specifically required for osteogenesis in the cranial neural crest lineage. They showed that differentiation of the mesenchyme and establishment of certain craniofacial structures was defective in Msx1-/-mice. They also showed that the failure of CNC-derived mesenchymal cells to express Runx2 and Osterix in the absence of Msx1 may prevent osteogenic differentiation. Runx2 is an essential transcription factor controlling osteoblast differentiation. Null mutation of Runx2 leads to a complete lack of ossification in both neural crest and mesoderm derived bones (Komori et al., 1997).
Targeted null mutation of Msx1 results in multiple craniofacial abnormalities involving a defect in mandibular bone development. In humans, mutations in the Msx1 gene have been implicated in tooth agenesis (Padanilam et al., 1992; Hu et al., 1998) and cleft palate (Van Den Boogaard et al., 2000), and the phenotype was proposed to be related to a dose effect of Msx1 protein (Hu et al., 1998). Interestingly, Msx1 down-regulation is associated with the terminal differentiation of several cell types, such as cartilage (Mackenzie et al., 1991; Coelho et al., 1993; Mina et al., 1995) and muscle (Houzelstein et al., 1999).
Our data indicate that expression of the Dlx1 gene at the E12 and E14 stages during craniofacial development is weaker in the Dumbo rat than in Wistar rat. The reduced expression of the Dlx1 gene in Dumbo rats might be implicated in the malformed genesis of the head in these rats. Depew et al. (2002, 2005) showed that Dlx-mutant mice exhibit severe craniofacial deformities, including cleft palate, and dysmorphic middle ear and jawbones. Dlx-mutant mice show delayed ossification of dermal bones (Merlo et al., 2000) resembling the defects caused by inactivation of one copy of Cbfa1 (Otto et al., 1997). It seems that both Msx1 and Dlx1 have a direct or indirect relation with Cbfa1.
Kim et al., (1998) showed that Fgfr2 expression was reduced in the craniofacial structures of Msx1-/- mouse embryos. There is evidence that FGF signalling is involved in calvarial development. In calvarial culture, FGF4 accelerates ossification. FGF2 can rescue the compromised osteogenitor proliferation of Tgfr2 conditional knockout mice (Sasaki et al., 2006). Robel et al., (1995) showed that FGF2 increased Dlx1 expression and that this effect was gene-specific, dose-dependent, and temporally regulated, with larger effects at earlier stages of development. This interaction between FGF2 and Dlx1 may be important for the regulation of the antero-posterior pattern in craniofacial development. Zhang et al., (1997) showed that some of the defects in Msx1-/- mice may be aggravated or rescued by controlling certain Dlx genes. The essential condition for this regulation to occur is that the two genes be expressed in the same cells at the same time.
In conclusion, we found that the Msx1 and Dlx1 genes are expressed differently during head development of Dumbo and Wistar rats, with a reduction of expression in the Dumbo strain. This suggests that the Dumbo rat could be a suitable experimental model for understanding abnormal craniofacial development. This rat reflects the relation between some homeobox genes and the craniofacial abnormalities. The search for other concomitant events related to craniofacial abnormalities will be necessary, such as studying apoptosis and the involvement of other genes in the Dumbo phenotype. Confirmation of our findings alsom requires studying the expression of the implicated genes by in situ hybridization and by investigating the expression of Msx1 protein by Western blot analysis.
Received: August 6, 2008; Accepted: December 16, 2008.
Ricardo Guelerman P. Ramos
- Bendall AJ and Abate-Shen C (2000) Roles for Msx and Dlx homeoproteins in vertebrate development. Gene 247:17-31.
- Blin-Wakkach C, Lezot F, Ghoul-Mazgar S, Hotton D, Monteiro S, Teillaud C, Pibouin L,Orestes-Cardoso S, Papagerakis P, Macdougall M, et al. (2001) Endogenous Msx1 antisense transcript: In vivo and in vitro evidences, structure, and potential involvement in skeleton development in mammals. Proc Natl Acad Sci USA 98:7336-7341.
- Coelho CN, Upholt WB and Kosher RA (1993) The expression pattern of the chicken homeobox-containing gene GHox-7 in developing polydactylous limb buds suggests its involvement in apical ectodermal ridge-directed outgrowth of limb mesoderm and in programmed cell death. Differentiation 52:129-137.
- Couly GF, Coltey PM and Le Douarin NM (1993) The triple origin of skull in higher vertebrates: A study in quail-chick chimeras. Development 117:409-429.
- de Jonge HJ, Fehrmann RS, de Bont ES, Hofstra RM, Gerbens F, Kamps WA, de Vries EG, van der Zee AG, te Meerman GJ and ter Elst A (2007) Evidence based selection of housekeeping genes. PLoS ONE 2:e898.
- Depew MJ, Lufkin T and Rubenstein JLR (2002) Specification of jaw subdivision by Dlx genes.Science 298:381-384.
- Depew MJ, Simpson CA, Morasso M and Rubenstein JL (2005) Reassessing the Dlx code: The genetic regulation of branchial arch skeletal pattern and development. J Anat 207:501-561.
- Dupont E, Canu MH, Stevens L and Falempin M (2005) Effects of a 14-day period of hindpaw sensory restriction on mRNA and protein levels of NGF and BDNF in the hindpaw primary somatosensory cortex. Brain Res Mol Brain Res 133 78-86.
- Forest-Potts L and Sadler TW (1997) Disruption of Msx-1 and MSX-2 reveals roles for these genes in craniofacial, eye, and axial development. Dev Dyn 209:70-84.
- Hall BK and Miyake T (2000) Craniofacial development of avian and rodent embryos. Meth Mol Biol 135:127-137.
- Han J, Ishii M, Bringas Jr P, Maas RL, Maxson Jr RE and Chai Y (2007) Concerted action ofMsx1 and Msx2 in regulating cranial neural crest cell differentiation during frontal bone development. Mech Dev 124:729-745.
- Houzelstein D, Cohen A, Buckingham ME and Robert B (1997) Insertional mutation of the mouse Msx1 homeobox gene by an nlacZ reporter gene. Mech Dev 65:123-133.
- Houzelstein D, Auda-Boucher G, Chéraud Y, Rouaud T, Blanc I, Tajbakhsh S, Buckingham ME, Fontaine-Pérus J and Robert B (1999) The homeobox gene Msx1 is expressed in a subset of somites, and in muscle progenitor cells migrating into the forelimb. Development 126:2689-2701.
- Hu G, Vastardis H, Bendall AJ, Wang Z, Logan M, Zhang H, Nelson C, Stein S, Greenfield N, Seidman CE, et al. (1998) Haploinsufficiency of MSX1: A mechanism for selective tooth agenesis. Mol Cell Biol 18:6044-6051.
- Kim HJ, Rice DP, Kettunen PJ and Thesleff I (1998) FGF-, BMP- and Shh-mediated signalling pathways in the regulation of cranial suture morphogenesis and calvarial bone development. Development125:1241-1251.
- Kissinger CR, Liu BS, Martin-Blanco E, Kornberg TB and Pabo CO (1990) Crystal structure of an engrailed homeodomain-DNA complex at 2.8 A resolution: A framework forunderstanding homeodomain-DNA interactions. Cell 63:579-590.
- Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al. (1997) Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 89:755-764.
- Louryan S, Biermans J and Flemal F (1995) Nerve growth factor in the developing craniofacial region of the mouse embryo. Eur J Morphol 33:415-419.
- Mackenzie A, Leeming GL, Jowett AK, Ferguson MW and Sharpe PT (1991) The homeobox gene Hox 7.1 has specific regional and temporal expression patterns during early murine craniofacial embryogenesis, especially tooth development in vivo and in vitroDevelopment 111:269-285.
- Mackenzie A, Ferguson MW and Sharpe PT (1992) Expression patterns of the homeobox gene, Hox-8, in the mouse embryo suggest a role in specifying tooth initiation and shape. Development 115:403-420.
- Merlo GR, Zerega B, Paleari L, Trombino S, Mantero S and Levi G (2000) Multiple functions of Dlx genes. Int J Dev Biol 44:619-626.
- Mina M, Gluhak J, Upholt WB, Kollar EJ and Rogers B (1995) Experimental analysis of Msx-1 and Msx-2 gene expression during chick mandibular morphogenesis. Dev Dyn 202:195-214.
- Morriss-Kay G (1993) Retinoic acid and craniofacial development: Molecules andmorphogenesis. Bioessays 15:1-5.
- Osumi-Yamashita N, Ninomiya Y, Doi H and Eto K (1994) The contribution of both forebrain midbrain crest cell to the mesenchyme in the frontonasal mass of mouse embryos. Dev Biol 164:409-419.
- Otto F, Thornell AP, Crompton T, Denzel A, Gilmour KC, Rosewell IR, Stamp GWH, Beddington RSP, Mundlos S, Olsen BR, et al. (1997) Cbfa, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differetiation and bone development. Cell 89:75-771.
- Padanilam BJ, Stadler HS, Mills KA, McLeod LB, Solursh M, Lee B, Ramirez F, Buetow KH and Murray JC (1992) Characterization of the human HOX 7 cDNA and identification of polymorphic markers. Hum Mol Genet 1:407-410.
- Pibouin-Fragner L, Nadra R, Fernandes I, Berdal A and Menuelle P (2005). Régulation hormonale du gène Msx1 chez le rat: Caractérisation et expression de l'ARN Msx1 antisens au cours du développement craniofacial. Cahiers de l'ADF 18-19:50-56.
- Price M, Lemaistre M, Pischetola M, Di Lauro R and Duboule D (1991) A mouse gene related to Distal-less shows a restricted expression in the developing forebrain. Nature 351:748-751.
- Qian YQ, Billeter M, Otting G, Müller M, Gehring WJ and Wüthrich K (1989) The structure of the Antennapedia homeodomain determined by NMR spectroscopy in solution: Comparison with prokaryotic repressors. Cell 59:573-580.
- Qiu M, Bulfone A, Martinez S, Meneses JJ, Shimamura K, Pedersen RA and Rubenstein JLR (1995) Role of Dlx-2 in head development and evolution: Null mutation of Dlx-2 results in abnormal morphogenesis of proximal first and second branchial arch derivatives and abnormal differentiation in the forebrain. Genes Dev 9:2523-2538.
- Robel L, Ding M, James AJ, Lin X, Simeone A, Leckman JF and Vaccarino FM (1995) Fibroblast growth factor 2 increases Otx2 expression in precursor cells from mammalian telencephalon. J Neurosci 15:7879-91.
- Robert B, Sassoon D, Jacq B, Gehring W and Buckingham M (1989) Hox-7, a mouse homeobox gene with a novel pattern of expression during embryogenesis. EMBO J 8:91-100.
- Sasaki T, Ito Y, Bringas P Jr, Chou S, Urata MM, Slavkin H and Chai Y (2006) TGFbeta-mediated FGF signaling is crucial for regulating cranial neural crest cell proliferation during frontal bone development. Development 133:371-381.
- Tan SS and Morriss-Kay G (1985) The development and distribution of the cranial neural crest in the rat embryo. Cell Tissue Res 240:403-16.
- Van Den Boogaard MJ, Dorland M, Beemer FA and van Amstel HK (2000) MSX1 mutation is associated with orofacial clefting and tooth agenesis in humans. Nat Genet 24:342-3.
- Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A and Speleman F (2002) Accurate normalization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genes. Genome Biol 3:e34.
- Zhang H, Hu G, Wang H, Sciavolino P, Iler N, Shen MM and Abate-Shen C (1997) Heterodimerization of Msx and Dlx homeoproteins results in functional antagonism. Mol Cell Biol 17:2920-2932.
Send correspondence to:
Publication Dates
-
Publication in this collection
01 May 2009 -
Date of issue
2009
History
-
Accepted
16 Dec 2008 -
Received
06 Aug 2008