Resumos
O objetivo desta revisão foi discutir aspectos que envolvem a compreensão do metabolismo da vitamina K e os recentes avanços quanto ao papel da vitamina na nutrição humana. São abordadas questões sobre as fontes da vitamina, sua absorção, distribuição e metabolismo, recomendações dietéticas, avaliação do estado nutricional relacionado à vitamina K e ainda fatores que afetam sua concentração plasmática. Diante das novas evidências sobre a participação da vitamina K no metabolismo ósseo, pesquisas adicionais são necessárias a fim de identificar as possíveis conseqüências à saúde, a longo prazo, de deficiência subclínica desta vitamina. Pesquisadores e comitês futuros encontram um desafio no que se refere a investigar novos requerimentos nutricionais da vitamina.
vitamina K; filoquinona; disponibilidade biológica; metabolismo; nutrição
The aim of this study was to discuss the aspects that involve the understanding of vitamin K metabolism and recent advances of its role in human nutrition. The paper presents dietary sources, absorption, distribution in the body and metabolism of vitamin K, nutritional recommendations, evaluation of nutritional status related to the vitamin and factors influencing its plasmatic concentration. In the light of evidences about vitamin K role in bone metabolism, further studies are needed to identify the possible long-term consequences to health of subclinical vitamin K deficiency. A great challenge for researchers and further scientific committees will be certainly concerned with investigation of new nutritional requirements of the vitamin.
vitamin K; philloquinone; biological availability; metabolism; nutrition
REVISÃO
VITAMINA K: METABOLISMO E NUTRIÇÃO
VITAMIN K: METABOLISM AND NUTRITION
Sílvia Maria Custódio das DÔRES1 (1 ) Doutoranda do Curso de Fisiopatologia em Clínica Médica, área de Nutrição e Metabolismo, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: silvia_btu@zipmail.com.br (2 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: paiva@fmb.unesp.br (3 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. Correspondência para/ Correspondence to: A.O. CAMPANA. E-mail: alvaroc@fmb.unesp.br
Sérgio Alberto Rupp de PAIVA2 (1 ) Doutoranda do Curso de Fisiopatologia em Clínica Médica, área de Nutrição e Metabolismo, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: silvia_btu@zipmail.com.br (2 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: paiva@fmb.unesp.br (3 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. Correspondência para/ Correspondence to: A.O. CAMPANA. E-mail: alvaroc@fmb.unesp.br
Álvaro Oscar CAMPANA3 (1 ) Doutoranda do Curso de Fisiopatologia em Clínica Médica, área de Nutrição e Metabolismo, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: silvia_btu@zipmail.com.br (2 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. E-mail: paiva@fmb.unesp.br (3 ) Departamento de Clínica Médica, Faculdade de Medicina, Universidade Estadual Paulista, Distrito de Rubião Júnior, s/n., 18618-000, Botucatu, SP, Brasil. Correspondência para/ Correspondence to: A.O. CAMPANA. E-mail: alvaroc@fmb.unesp.br
RESUMO
O objetivo desta revisão foi discutir aspectos que envolvem a compreensão do metabolismo da vitamina K e os recentes avanços quanto ao papel da vitamina na nutrição humana. São abordadas questões sobre as fontes da vitamina, sua absorção, distribuição e metabolismo, recomendações dietéticas, avaliação do estado nutricional relacionado à vitamina K e ainda fatores que afetam sua concentração plasmática. Diante das novas evidências sobre a participação da vitamina K no metabolismo ósseo, pesquisas adicionais são necessárias a fim de identificar as possíveis conseqüências à saúde, a longo prazo, de deficiência subclínica desta vitamina. Pesquisadores e comitês futuros encontram um desafio no que se refere a investigar novos requerimentos nutricionais da vitamina.
Termos de indexação: vitamina K, filoquinona, disponibilidade biológica, metabolismo, nutrição.
ABSTRACT
The aim of this study was to discuss the aspects that involve the understanding of vitamin K metabolism and recent advances of its role in human nutrition. The paper presents dietary sources, absorption, distribution in the body and metabolism of vitamin K, nutritional recommendations, evaluation of nutritional status related to the vitamin and factors influencing its plasmatic concentration. In the light of evidences about vitamin K role in bone metabolism, further studies are needed to identify the possible long-term consequences to health of subclinical vitamin K deficiency. A great challenge for researchers and further scientific committees will be certainly concerned with investigation of new nutritional requirements of the vitamin.
Index terms: vitamin K, philloquinone, biological availability, metabolism, nutrition.
INTRODUÇÃO
A literatura nacional a respeito da vitamina K é bastante escassa, notadamente quanto a aspectos referentes ao seu metabolismo. Estudos anteriores sobre situações que induzem deficiência de vitamina K (Paiva et al, 1998) e a possível variação da ingestão dietética da vitamina, provocando oscilações do controle terapêutico da anticoagulação oral, motivaram o interesse do nosso grupo de Nutrição para o desenvolvimento de estudos concernentes ao metabolismo dessa vitamina.
Foram pesquisados os bancos de dados MEDLINE e LILACS. O objetivo deste trabalho foi discutir aspectos relacionados as fontes da vitamina, sua absorção, distribuição, recomendações dietéticas, fatores que afetam sua concentração plasmática e a avaliação do estado nutricional relacionado à vitamina K. Discutem-se ainda, os recentes avanços quanto ao papel da vitamina na nutrição humana.
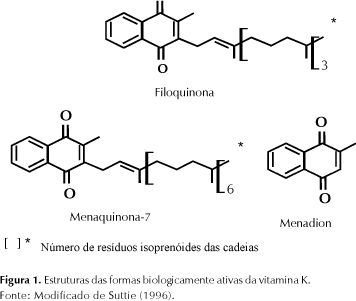
A vitamina K foi descoberta por Henrik Dam em 1929, como um fator anti-hemorrágico, capaz de restabelecer perturbações sangüíneas observadas em galinhas, alimentadas com dieta livre de gordura (Suttie, 1992). Em 1939, Dam na Dinamarca e Doisy em St. Louis isolaram a vitamina K1 da alfafa e determinaram sua exata estrutura: 2-metil-3-phytyl-1,4 naftoquinona. As formas naturais de vitamina K são a filoquinona e as menaquinonas. A vitamina K1, hoje chamada de filoquinona, é o único analógo da vitamina presente em plantas; é encontrada em hortaliças e óleos vegetais, os quais representam a fonte predominante da vitamina. A forma sintetizada por bactérias, as menaquinonas, originalmente chamadas de K2, foram subseqüentemente caracterizadas (Dowd et al., 1995). A família das menaquinonas constitui-se numa série de vitaminas designadas MK-n, onde o n representa o número de resíduos isoprenóides na cadeia lateral. As menaquinonas naturais variam de MK-4 a MK-13 (Vermeer et al., 1995). A menadiona, (2-metil-1,4 naftoquinona), é um composto sintético normalmente utilizado como fonte da vitamina para a alimentação animal (Suttie, 1996), (Figura 1).
Diversas investigações conduzidas de meados dos anos 60s a meados dos anos 70s, culminaram com a descoberta de fatores de coagulação (protrombina, fatores VII, IX, e X) que continham um aminoácido, o ácido gama carboxiglutâmico (Gla). Outras duas proteínas adicionais contendo Gla, a proteína S e C, atuam como inibidoras do mecanismo de coagulação (Suttie, 1992). A vitamina K atua como cofator essencial na reação de carboxilação de resíduos específicos de ácido glutâmico (Glu), levando à formação de Gla (Figura 2). A carboxilação capacita as proteínas de coagulação a se ligarem ao cálcio, permitindo assim a interação com os fosfolipídios das membranas de plaquetas e células endoteliais, o que, por sua vez, possibilita o processo de coagulação sangüínea normal. No início dos anos 90s foi isolada e caracterizada a enzima hepática carboxilase, envolvida na ativação dessas proteínas (Suttie, 1996).
Foram descobertos diversos grupos de proteínas dependentes de vitamina K, que não têm conexão com a coagulação sangüínea, mas estão implicados na homeostasia do cálcio. Proteínas contendo Gla são conhecidas por ocorrerem em um grande número de tecidos e órgãos como osso, rim, placenta, pâncreas, vesícula e pulmão. Na maioria dos tecidos, essas proteínas ainda não foram totalmente caracterizadas. É exceção a proteína do osso chamada osteocalcina ou proteína Gla do osso. É uma proteína de baixo peso molecular, com 3 resíduos de Gla, produzida por osteoblastos durante a formação da matriz óssea (Mijares et al., 1998). É uma das mais abundantes proteínas não colagenosas na matriz extracelular do osso; sua dosagem no sangue constitui importante marcador biológico da atividade osteoblástica. Existem atualmente diversas evidências de que a vitamina K é importante tanto no desenvolvimento precoce do esqueleto, quanto na manutenção do osso maduro sadio (Shearer, 1995). Uma segunda proteína isolada do osso e estruturalmente semelhante à osteocalcina, a proteína Gla da matriz, também é dependente de vitamina e seu papel fisiológico não é conhecido (Suttie, 1996). Nas placas ateroscleróticas, encontra-se outra proteína Gla denominada, Proteína Gla da placa, a qual parece evitar o endurecimento e mineralização da parede arterial (Gijbers et al., 1990).
Em recente publicação (Berkner, 2000) são citadas outras proteínas dependentes de vitamina K, como a proteína produzida pelo growth-arrest-specific gene 6 (gas 6), ligada ao controle de crescimento, as proteínas Gla ricas em prolina 1 e 2, com função potencial na transdução de sinais, e a proteína Z, cujas funções não são conhecidas. Para Berkener (2000), a própria enzima carboxilase pode ser considerada uma proteína dependente de vitamina K. Esta autora evidenciou a probabilidade da existência de outras proteínas dessa natureza, devido à grande distribuição tecidual da carboxilase.
FONTES ALIMENTARES / NUTRICIONAIS
A forma predominante de vitamina K nos alimentos é a filoquinona. A vitamina K é amplamente distribuída em alimentos de origem animal e vegetal, variando de 1 mg por 100 mL no leite a 400 mg por 100 g no espinafre e outras hortaliças (Olson, 1999). As carnes são fontes pobres em filoquinona. Elas provavelmente contribuem com menos que 5 mg por 100 g e, em muitos casos, menos do que 1 mg por 100 gramas de alimento. O fígado apresenta quantidades maiores, evidentemente por ser o maior órgão de estoque da vitamina (20-100 mg por 100 g), mas essa quantidade varia, dependendo da ingestão do animal antes do abate. Muitos peixes, cereais, e bebidas contêm pequenas mas mensuráveis quantidades de filoquinona (Shearer, 1995). Enquanto folhas de chá e grãos de café contêm apreciáveis quantidades de filoquinona, as infusões não são fontes importantes como se pensava (Booth et al., 1995a). Outros alimentos como frutas (polpa e suco), raízes e tubérculos são pobres em filoquinona (Booth & Suttie, 1998). Quanto ao leite, sabe-se que o teor de filoquinona do leite materno é menor do que o do leite de vaca: aproximadamente 1 a 3 mg por litro no leite humano contra 5 a 10 mg por litro no leite de vaca. A vitamina está concentrada na porção lipídica do leite e de laticínios ricos em gordura (Suttie, 1992).
Uma fonte importante de filoquinona é representada pelos óleos e pelas gorduras. As manteigas contêm aproximadamente 10 mg por 100 gramas, enquanto há grande variação nos óleos vegetais, sendo que os mais ricos, que contêm de 200-400 mg por 100 gramas, são os óleos de soja e oliva (Booth & Suttie, 1998). Vegetais de folhas verdes contêm maior teor de filoquinona e contribuem com 40-50% da ingestão total (Fenton et al., 1997). Os altos valores encontrados nesses vegetais confirmam a conhecida associação da filoquinona com tecidos capazes de realizar fotossíntese (Shearer et al., 1996). A distribuição de filoquinona nas plantas não é uniforme; maiores concentrações da vitamina são encontradas nas folhas externas quando comparadas às folhas mais internas. A casca das frutas e dos vegetais parece ter maiores concentrações da vitamina do que a polpa. Fatores como a estação do ano, o clima, local geográfico e a fertilização do solo afetam as concentrações de vitamina K1 nos alimentos (Booth et al., 1993).
Uma análise recente de mais de 260 alimentos freqüentemente consumidos nos EUA revelou que existem muitas fontes dietéticas de filoquinona que têm sido negligenciadas (Booth et al., 1995b). Alimentos que são pobres em filoquinona em sua forma crua, podem ter papel importante se forem processados, utilizando-se óleos ricos em filoquinona (Sadowski et al., 1996).
O feijão, importante alimento da dieta dos brasileiros, foi identificado como alimento que contribui significativamente para suprir as necessidades de vitamina K em 115 pacientes ambulatoriais de Hospital do interior do Estado de São Paulo, por meio de estudo longitudinal empregando o questionário de freqüência alimentar (Dôres et al., 2000). Ferland & Sadowski (1992) mostraram que a vitamina K1 contida nos óleos vegetais é estável ao calor e ao processamento, mas é rapidamente destruída pela luz fluorescente e natural. O óleo de canola perde 87% da vitamina após dois dias de exposição à luz do dia. Esses autores sugerem que a estocagem desses óleos, em embalagens opacas, preserva a vitamina, enquanto embalagens transparentes permitem que a iluminação ambiente reduza o conteúdo da mesma (Ferland & Sadowski, 1992). Langenberg et al. (1986), usando o método de Cromatografia Líquida de Alta Eficiência (CLAE), observaram que nem o cozimento, nem irradiações gama afetam o conteúdo de filoquinona de vegetais selecionados (ervilhas, espinafre e repolho); verificaram também que o conteúdo de filoquinona dos alimentos comercialmente disponíveis na forma de preparações vegetais secas e congeladas não difere daqueles conteúdos de vegetais frescos.
Óleos vegetais hidrogenados, ricos em filoquinona, são amplamente utilizados na indústria por suas características físicas e estabilidade oxidativa. Durante a hidrogenação, há conversão da filoquinona a 2-3 dihidrofiloquinona (dK) (Davidson et al., 1996). Alimentos preparados com alto conteúdo de gordura hidrogenada contêm 30-60 mg dK/100 g (Booth et al., 1996a). A importância fisiológica da dK depende de sua atividade biológica, que ainda não é conhecida; portanto, a influência da ingestão de alimentos com elevadas concentrações de óleos hidrogenados, no estado nutricional relacionado à vitamina K, não é conhecida até que novos estudos sejam desenvolvidos (Booth & Suttie, 1998).
Um dos maiores problemas quanto à obtenção dos teores de filoquinona da dieta, é aquele relacionado com as próprias tabelas de composição dos alimentos. Muitas tabelas são baseadas em avaliações biológicas que têm amplas margens de variação analítica. Esses valores de filoquinona ainda são encontrados na maioria das tabelas de composição comumente utilizadas (Sadowski et al., 1996). Esse problema está agora sendo resolvido através do uso de um método sensível e específico baseado em CLAE (Shearer, 1995). Tabelas americanas contendo a composição dos alimentos, exclusivamente geradas por CLAE, já podem ser encontradas para utilização rotineira (Tabela 1).
Menaquinonas como fonte nutricional
Embora haja um conhecimento crescente com relação ao teor de filoquinona nos alimentos, existe pouca informação sobre os alimentos fontes de menaquinonas. Sabe-se de longa data que fígados de diversas espécies animais são boas fontes de ampla variedade de menaquinonas, com cadeias laterais variando de MK-6 a MK-13 (Duello & Matschiner, 1970). Entretanto, análises quantitativas, por CLAE, sugerem que só o fígado de ruminantes, como vaca, contém concentrações suficientemente elevadas de algumas menaquinonas (MK-7, 11, 12, 13), que parecem ser de significância nutricional (variando de 10-20 mg por 100 gramas). Por outro lado, em função de o fígado ser um alimento de consumo esporádico pela maioria da população, o impacto dessas concentrações na nutrição humana parece ser pequeno. As concentrações de menaquinonas em outros órgãos animais (rim, coração e músculo) são muito baixas, nutricionalmente insignificantes (Shearer et al., 1996). Esses mesmos autores observaram, ainda, que vários queijos contêm 5-20 mg/100 g de MK-8 e 9. Quantidades limitadas de menaquinonas têm sido encontradas em produtos animais, como gema de ovo e manteiga (Hirauchi et al., 1989). Produtos fermentados, à base de soja, contêm quantidades substanciais de MK6 e MK8 e podem ser de importância nutricional para as populações consumidoras dessa classe de alimentos (Sakano et al., 1988).
Quanto à vitamina K sintetizada pelas bactérias, sabe-se que o intestino humano contém grandes quantidades de bactérias produtoras de menaquinonas; contudo, sua importância nutricional não é clara. A extensão e o mecanismo de absorção dessas menaquinonas, no intestino grosso, aparentemente é limitada, embora seja conhecido que o fígado humano apresente quantidades significativas dessa forma da vitamina (Suttie, 1995). Suttie (1996) relata que as menaquinonas são importantes na nutrição humana, porém contribuem relativamente pouco para suprir os requerimentos de vitamina K, ao contrário do que era previamente concebido.
Numerosos estudos utilizando antibióticos a fim de induzir hipoprotrombinemia foram conduzidos, considerando que as menaquinonas contribuiriam significativamente para os requerimentos humanos de vitamina K. Entretanto, recentes pesquisas (Suttie, 1995) com o uso de dietas restritas em vitamina K, argumentam contra sua significância nutricional. As evidências de decréscimo da síntese de menaquinonas, na presença de tratamento com antibióticos, são mínimas; contudo, observou-se que alguns tipos de antibióticos, os que possuem uma cadeia lateral N-metiltiotetrazole (NMTT), têm a capacidade de inibir epoxi-redutase de vitamina K (Dowd et al., 1995). O resultado é um efeito similar aos cumarínicos, embora acentuadamente mais fraco e observado somente naqueles pacientes que apresentam estado nutricional comprometido quanto à vitamina K (Suttie, 1995).
Recentemente, um estudo com o objetivo de testar o grau de contribuição das menaquinonas, produzidas pela flora bacteriana, em relação aos requerimentos de vitamina K, foi conduzido em indivíduos utilizando Omeprazol (bloqueador de bomba de hidrogênio) e dieta restrita em filoquinona. Os resultados evidenciaram que o crescimento bacteriano, promovido pelo aumento do pH gástrico, resulta em síntese e absorção de menaquinonas, porém estas não foram produzidas em quantidades suficientes para restaurar o estado normal de vitamina K (Paiva et al., 1998).
Absorção, distribuição e metabolismo
A vitamina K da dieta é absorvida no intestino delgado, incorporada aos quilomícrons e transportada pelas vias linfáticas; requer bile e suco pancreático para máximo aproveitamento. A eficiência na absorção foi mensurada em 40-80%, dependendo do veículo no qual a vitamina é administrada e da circulação enterohepática. Quando a filoquinona é administrada em seres humanos, oralmente, em doses variando do nível fisiológico ao farmacológico, a vitamina aparece no plasma dentro de 20min, com pico em 2h; a seguir, declina exponencialmente a baixos valores, durante 48-72 horas, alcançando níveis de jejum de 1 a 2 nM ( 0,5¾1,0 ng/mL) (Olson, 1999). Lamon-Fava et al. (1998) observaram que as lipoproteínas ricas em triacilgliceróis são as principais carreadoras de filoquinona, transportando 83,0% da filoquinona plasmática, sendo as lipoproteínas de baixa e alta densidade (LDL e HDL) carreadoras menos importantes (7,1% e 6,6%, respectivamente).
Ao alcançar o fígado, a filoquinona é reduzida a hidronaftoquinona (KH2), que é o cofator ativo para a carboxilase (Davidson & Sadowski, 1997). O fígado tem um papel exclusivo na transformação metabólica que leva à excreção da vitamina K do organismo. A fração da vitamina excretada não é dependente da dose administrada. Assim, independentemente da dose administrada, por exemplo 1 mg ou 45 mg, aproximadamente 20% são excretados na urina dentro de 3 dias, enquanto 40-50% são excretados nas fezes, via sais biliares (Shearer et al., 1974). Esse extenso catabolismo da filoquinona pelo fígado explica o rápido turnover e a depleção das reservas hepáticas em pacientes com dieta pobre em filoquinona, observado por Usui et al. (1990). Parece provável, portanto, que aproximadamente 60-70% das quantidades de filoquinona, absorvidas em cada refeição, são definitivamente perdidas por excreção (Shearer et al., 1996), o que sugere que os estoques corporais de filoquinona são constantemente reabastecidos. Com relação aos estoques hepáticos, sabe-se que este inclui aproximadamente 90% de menaquinona e 10% de filoquinona. Os estoques de filoquinona são extremamente lábeis e sob condições de grave depleção nutricional podem reduzir-se a 25% de suas concentrações originais, após três dias (Usui et al., 1990). As concentrações hepáticas de filoquinona em adultos são de aproximadamente 5 ng/g (Shearer et al., 1988); porém, a relação entre os estoques hepáticos de vitamina K e os estoques corporais totais não são conhecidos (Shearer et al., 1996).
Estudo feito em tecidos humanos, retirados de cadáveres, mostrou que existem, no homem, padrões de distribuição tecidual de vitamina K comparáveis aos já observados em ratos. Elevados níveis de vitamina K1 foram encontrados no fígado, coração e pâncreas e níveis mais reduzidos no cérebro, rim e pulmão. A MK-4 foi encontrada na maioria dos tecidos, excedendo os níveis de K1 no cérebro e rim. Sabe-se pouco sobre a captação de vitamina K pelos tecidos; entretanto, o acúmulo de vitamina K no coração, cérebro e pâncreas sugere funções fisiológicas ainda desconhecidas da vitamina (Thijssen & Drittij-Reijnders, 1996).
Por ser o local de síntese de proteínas da coagulação dependentes de vitamina K, o fígado sempre é considerado o maior órgão de estoque das vitaminas K (Shearer, 1995); entretanto, o osso cortical contém tanta vitamina K quanto o fígado, podendo funcionar como um fornecedor de filoquinona (Hodges et al., 1993). Vale notar que o espectro das vitaminas K circulantes no plasma não reflete os estoques hepáticos (Shearer et al., 1996).
Pouco se sabe sobre o transporte e depuração da circulação das menaquinonas (Suttie, 1995), mas tem sido sugerido que a via de degradação metabólica seja similar à da filoquinona (Suttie, 1996).
Ciclo da vitamina K
Em essência, o ciclo de vitamina K, pode ser considerado uma via de recuperação da vitamina, presente em quantidades nanomolares no fígado e em outros tecidos (Olson, 1999).
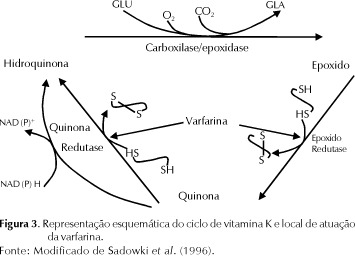
Sempre que um resíduo de glutamato (Glu) é carboxilado, a vitamina K é oxidada, dando origem à forma 2,3-epoxi (Kohlmeier et al., 1996). Esse metabólito é convertido novamente à sua forma ativa, pela ação da enzima microssomal, epoxi redutase de vitamina K e uma ou mais quinona redutases de vitamina K (Suttie et al., 1988). A ação da epoxi-redutase é inibida por cumarínicos como a varfarina, resultando, conseqüentemente, em indução de antagonismo à vitamina K (Suttie, 1992).
Aproximadamente 40 mmol de Gla são excretados diariamente (Ferland et al., 1993) e quantidades equimolares de vitamina K são oxidados. Os requerimentos diários, em contraste, não são maiores do que 0,2 mmol; é razoável assumir, portanto, que, em média, uma molécula de vitamina K é reciclada várias centenas de vezes (Kohlmeier et al., 1996). O resultado líquido do ciclo (Figura 3) é a conversão da epoxi-vitamina K em hidroquinona, que se torna disponível para novos processos de carboxilação (Olson, 1999).
Biodisponibilidade
Gijsbers et al. (1996) observaram que a biodisponibilidade da vitamina K é menor do que se imagina e depende da forma pela qual a vitamina é consumida. Em seu estudo, observaram que a filoquinona é prontamente absorvida a partir de um concentrado farmacêutico de vitamina K (Kanakion), atingindo o pico sangüíneo em 4 horas. Para a filoquinona do alimento, o pico é atingido mais lentamente, indicando que a absorção da vitamina nos vegetais é processo mais demorado, influenciado por fatores digestivos. Ocorre, ainda, variação interindividual com respeito às quantidades de vitamina K que podem ser extraídas dos vários alimentos, podendo a secreção de bile ter papel importante nessas diferenças. O autor observou, no mesmo estudo, que a biodisponibilidade de 1 mg de filoquinona, no espinafre, em seres humanos, foi de apenas 4%, comparado à filoquinona pura (Kanakion). Com adição de gordura (manteiga) ao espinafre, houve aumento de absorção para 13%. O efeito da gordura dá-se provavelmente pela estimulação da secreção de bile, que se sabe ser importante para absorção de compostos hidrofóbicos (Shearer et al., 1974).
Recentemente, Garber et al. (1999) também estudaram a biodisponibilidade da filoquinona. Foi comparada a biodisponibilidade da vitamina na forma de suplemento (tablete com 500 µg/filoquinona) de acordo com as fontes alimentares. Foram observados os seguintes resultados:
- quando ingerida na forma pura, a absorção de filoquinona é seis vezes maior do que quando presente em algum alimento (ex: espinafre). A absorção de filoquinona do tablete é mais rápida, atingindo o pico de concentração sérica em 2 ou 3 horas;
- não foi observada a influência da cocção assim como do teor de gorduras das refeições quanto à biodisponibilidade de filoquinona;
- também não foi observada diferença significativa na absorção de filoquinona quando as ingestões de espinafre, brócolis e alface foram comparadas, sugerindo que a seleção desses alimentos específicos não tem influência na biodisponibilidade da vitamina K.
Os autores, entretanto, fizeram a ressalva de que é necessária a realização de estudo com amostragem ampliada, uma vez que seu trabalho foi efetuado com um número bastante reduzido de indivíduos.
Vitaminas K no sangue
A concentração de vitamina K no plasma, em jejum, de pessoas saudáveis é menor do que 1 ng/mL (2,2nmol/L) (Olson, 1994). A variação normal é de 0,4 a 2,2 nmol/L (0,2-1,0 ng/mL) com média de 1,1 nmol/L (Sadowski et al., 1989).
Avanços metodológicos recentes têm possibilitado mensurar rotineiramente as concentrações circulantes de filoquinona. Em estudo com 326 indivíduos, Sadowski et al. (1989) observaram uma variação normal de 0,29 a 2,64 nmol/L, com média geométrica de 0,87 nmol/L.
As concentrações de filoquinona são muito mais baixas do que as de outras vitaminas lipossolúveis. A concentração plasmática de filoquinona em jejum é 50 vezes menor do que a concentração da vitamina D, assim como é 2 mil vezes menor em relação ao retinol e 200 mil vezes menor em relação ao tocoferol. As concentrações plasmáticas pós-absortivas de filoquinona são notadamente elevadas; valores variando de 1 a 3 mmol/L podem ser observados. Os fatores que influenciam essas concentrações e sua relação com a ingestão dietética ainda não estão totalmente esclarecidos (Suttie, 1992).
Olson (1984) estimou o tamanho do pool corporal, como sendo de 0,22 mmol (100 mg) ou 3,1 nmol/kg de peso corporal. Esse pool de vitamina K é menor em relação à vitamina B12 e excepcionalmente baixo para uma vitamina lipossolúvel.
Fatores não dietéticos como idade, sexo, e/ou menopausa parecem afetar o metabolismo de vitamina K (Booth & Suttie, 1998).
Indivíduos com idade acima de 60 anos apresentam concentrações de filoquinona significativamente maiores em relação a adultos jovens (abaixo de 40 anos), independentemente da ingestão dietética (Ferland et al., 1993; Bach et al., 1996). Em estudo populacional, a menor concentração plasmática de filoquinona foi verificada durante a terceira década de vida, tanto para homens quanto para mulheres; após esse período, as concentrações aumentam e permanecem constantes (Sokoll & Sadowski, 1996). Sadowski et al. (1989) também observaram aumento da filoquinona plasmática em idosos, mais pronunciado em mulheres. As diferenças de concentração de filoquinona de acordo com a idade são influenciadas pelas concentrações de triacilgliceróis, que também aumentam com a idade. De fato, como a filoquinona é incorporada aos quilomícrons após absorção e é transportada para o fígado nas lipoproteínas, ricas em triacilgliceróis (Shearer, 1995), existe uma forte correlação positiva entre a filoquinona plasmática e as concentrações de triacilgliceróis (Kohlmeier et al., 1995). A relação entre filoquinona e triacilgliceróis no plasma (nmol de filoquinona/mmol de triacilgliceróis) é menor em idosos do que em jovens; isto tem sido interpretado ou como decorrente de uma menor absorção da vitamina ou como aumento de seus requerimentos ou, ainda, como, indicativo de menor ingestão de alimentos ricos em vitamina K (Sadowski et al., 1989).
A concentração de filoquinona plasmática é fortemente influenciada pelo polimorfismo genético da apolipoproteína E (apo E), sendo maior nos pacientes com variante apo E2, intermediária na apo E3 e menor na apo E4. Esse fato está associado ao ritmo de clareamento hepático dos quilomícrons remanescentes da circulação, que é menor para apo E2, mais rápido para apo E3 e mais rápido ainda para apo E4 (Shearer, 1995). A explicação desses achados fundamenta-se no fato de que as diferentes apolipoproteínas apresentam diferentes afinidades pelos receptores. Como conseqüência, há diferenças com relação à taxa de depuração plasmática dos quilomícrons, o que influencia a concentração de filoquinona (Vermeer et al., 1996).
Sadowski et al. (1989) observaram que o aumento do consumo de álcool resultou em progressiva redução nos níveis plasmáticos de filoquinona. A ingestão média de etanol dos indivíduos estudados foi de 16 g/dia; o consumo acima de 16 g/dia foi associado com decréscimo dos níveis de filoquinona circulantes, sendo mais pronunciado em idosos. Esses autores relataram ainda, que existe discreto aumento das concentrações de filoquinona séricas nos meses de verão, seguido por menores níveis no inverno. Uma possível explicação para esse fato seria a fonte dietética, uma vez que vários alimentos, ricos em filoquinona, sofrem ação sazonal.
AVALIAÇÃO NUTRICIONAL RELACIONADO À VITAMINA K
A avaliação individual do estado da vitamina K depende da aplicação apropriada de três procedimentos classicamente conhecidos: histórico, exame físico e exames laboratoriais.
Histórico e Exame físico
O histórico constitui-se em importante instrumento para detectar os indivíduos sob risco de desenvolver a deficiência vitamínica, que em geral são recém-natos, indivíduos com dietas pobres em vitamina K ou com má absorção intestinal. A história deve incluir questões sobre eventos hemorrágicos observados na boca, nariz, trato gastrointestinal (hematêmese, melena), rim (hematúria) e sob a pele (equimoses). Devem fazer parte do histórico do paciente dados acerca do uso de drogas, principalmente anticoagulantes cumarínicos. Complementarmente, se faz necessária a obtenção da história dietética abrangente, que inclui alimentos freqüentemente consumidos, recordatório de 24 horas e ocasionalmente registro alimentar de três dias (Olson, 1999).
No exame físico, buscam-se evidências de sinais hemorrágicos, manifestação cardinal da deficiência de vitamina K. Pode estar presente um só sinal ou mais dos seguintes: sangramento nasal ou oral, equimoses na virilha, garganta e nas pernas; sinais hemorrágicos sob unhas ou na conjuntiva; melena; hematúria e hematêmese. Palidez pode ser um sinal prévio de sangramento (Olson, 1999).
Avaliação laboratorial
Os métodos laboratoriais tradicionais para avaliação do estado nutricional relacionado à vitamina K são baseados em testes funcionais como tempo de protrombina (TP) e outros tempos de sangramento (Sokoll et al., 1997). Porém, sabe-se que o TP pode permanecer normal, mesmo quando a concentração de protrombina declina em 50% no plasma (Suttie, 1992) em conseqüência, este pode ser considerado um teste de baixa sensibilidade para detectar deficiências subclínicas de vitamina K.
A concentração da filoquinona plasmática não se correlaciona adequadamente com o estado nutricional da vitamina K, pois é dependente da ingestão recente da vitamina (nas 24 horas) (Booth & Suttie, 1998). Booth et al. (1997) mostraram que, entre todas as vitaminas lipossolúveis, somente a filoquinona apresentou correlação significativa entre a ingestão dietética e as concentrações plasmáticas em jejum. Em trabalho anterior, Kindberg & Suttie (1989) observaram em ratos que, sob condições de restrição de vitamina K dietética, as concentrações hepáticas de filoquinona mostram-se correlacionadas ao suprimento dietético da vitamina. Entretanto, somente quando o fígado contém vitamina K suficiente para garantir atividade ótima da carboxilase hepática, é que a filoquinona será liberada em quantidade suficiente pelo fígado de forma à elevar suas concentrações séricas. Conclui-se, portanto, que o nível sérico de filoquinona não se constitui em marcador ideal para a avaliação do estado nutricional relacionado à vitamina K.
Com a descoberta de novas funções da vitamina, principalmente no metabolismo ósseo, novos indicadores do estado nutricional, foram identificados e incluem: a excreção urinária de Gla, a proteína induzida pela deficiência ou antagonismo de vitamina K-Protein Induced by Vitamin K Absence or Antagonism (PIVKA) e a osteocalcina pouco carboxilada -undercarboxylated Osteocalcin (ucOc) (Shearer, 1995).
O Gla é normalmente excretado na urina, por adultos, em taxa de 40 mmol por dia. Essa substância é liberada durante o catabolismo das proteínas dependentes de vitamina K e aparece na urina sem alterações (Olson, 1999). Na vigência de deficiência vitamínica, as proteínas dependentes de vitamina K são sintetizadas com um número reduzido de resíduos de Gla, o que condiciona menor excreção urinária de Gla nesse período. A excreção de Gla portanto, pode ser medida como um índice do catabolismo dessas proteínas.
Da mesma forma, durante períodos de deficiência de vitamina K, várias proteínas que contêm Gla, entram na circulação na forma total ou parcialmente descarboxilada, sendo, portanto, importantes indicadores do estado nutricional relacionado à vitamina K (Ferland et al., 1993). As proteínas mais freqüentemente citadas em estudos são a protrombina descarboxilada (PIVKA II) e a osteocalcina descarboxilada (ucOc). Vermeer et al. (1995) relataram que, à medida que a ingestão de filoquinona decresce, a osteocalcina circulante parece ser a primeira proteína Gla a aparecer no plasma na forma descarboxilada; portanto, os autores consideram a ucOc o marcador mais sensível do estado nutricional relativo à vitamina K. A PIVKA II é medida no plasma por anticorpos específicos; em pessoas saudáveis, sua concentração é próxima a zero, enquanto que, em pessoas com deficiência de vitamina K, os valores podem aumentar em até 30% de toda a protrombina (Olson, 1999).
A sensibilidade de diversos indicadores é avaliada quando o estado de vitamina K é alterado tanto por dietas restritivas como pela administração de drogas antivitamina K (varfarina). Alison et al. (1987) mantiveram 33 indivíduos em dieta líquida padronizada contendo aproximadamente 5 mg de vitamina K por dia, durante 13 dias. As concentrações de filoquinona plasmática caíram em média 70% e a PIVKA II elevou-se em 67%. Essas alterações ocorreram tanto em controles como em indivíduos tomando antibióticos. Suttie et al. (1988) estudaram 10 indivíduos nos quais a ingestão de vitamina K foi reduzida em média de 82 mg para menos de 40 mg por dia, por restrição voluntária da ingestão de vegetais e saladas verdes. Em três semanas, a filoquinona plasmática caiu 50% e houve aumento significativo da PIVKA II e decréscimo da excreção de Gla urinário. Ferland et al. (1993) estudaram 32 indivíduos em unidade metabólica; após um período de consumo de dieta com aproximadamente 80 mg de filoquinona/dia, os participantes do estudo foram colocados em dieta contendo aproximadamente 10 mg /dia. Após 13 dias, a excreção de Gla caiu e PIVKA aumentou. Uma reposição gradual de vitamina K com 45 mg promoveu elevação dos níveis de Gla urinário para os valores basais; entretanto, os níveis de TP não se alteraram nesse estudo.
Sokoll & Sadowski (1996) mostraram também que, num grupo de 263 indivíduos saudáveis, a ucOc parece refletir o estado nutricional de vitamina K, porque existe uma associação negativa significante entre a porcentagem de ucOc e a filoquinona plasmática. Os indivíduos com os menores valores de filoquinona plasmática apresentaram, em média, maiores concentrações de ucOc e vice-versa. Além disso, as concentrações de ucOc se correlacionam positivamente com as concentrações de PIVKA II. Esses mesmos autores mostraram que existe influência da idade, sexo e menopausa nos indicadores, como filoquinona plasmática, Gla urinário, PIVKA II, osteocalcina sérica, e ucOc; entretanto, este fato parece não constituir impedimento para sua utilização na avaliação do estado da vitamina K.
O antagonismo à vitamina K, produzido pela administração de doses baixas (minidoses) do anticoagulante varfarina, também é usado para avaliar a sensibilidade de vários marcadores de vitamina K. Foi mostrado previamente que o TP permaneceu em níveis normais enquanto a porcentagem de osteocalcina descarboxilada (ucOc) aumentou em média 170% em relação aos níveis basais de 9 indivíduos, após receberem 1 mg de varfarina/dia por 7 dias. Observou-se, após reposição com 5 mg de filoquinona/dia, durante 2 dias, decréscimo nas concentrações de ucOc, abaixo das concentrações basais (Sokoll et al., 1995).
Bach et al. (1996) estudaram dois grupos de indivíduos, nove idosos e nove jovens. Após período pré-tratamento, onde todos consumiam suas dietas habituais (em torno de 1 mg/Kg/dia de filoquinona nos jovens e aproximadamente o dobro nos idosos), foram administradas minidoses de varfarina (1mg/dia), durante 14 dias. Esse período foi sucedido pelo fornecimento de 1 mg de filoquinona/dia, por 5 dias, para reversão da deficiência provocada de vitamina K. Foi notada a elevação das concentrações de PIVKA II, mas não decréscimo significativo do Gla urinário. A alteração mais importante foi o aumento das concentrações de ucOc em até três vezes, em relação ao período pré-tratamento, nos indivíduos jovens e em até seis vezes nos idosos. As concentrações de ucOc, durante a suplementação com vitamina K, foram menores não só em relação ao último dia do uso da varfarina, como também em relação ao período pré-tratamento. Esses autores sugerem que o antagonismo à vitamina K, resultante da inibição da enzima epoxi redutase, pode produzir efeito diverso na produção de proteínas por diferentes tecidos, sendo a ucOc o indicador mais sensível do referido estado de antagonismo à vitamina K, resultante de tratamento com varfarina. Deve-se destacar portanto, que o comportamento de alguns marcadores nutricionais na deficiência de vitamina K são distintos daqueles observados na situação de antagonismo à vitamina. Observa-se na deficiência alterações como: redução dos níveis séricos de filoquinona e aumento da PIVKA II (Paiva et al., 1998), enquanto que no estado de antagonismo, a alteração mais preponderante é a elevação acentuada da ucOc e aumento da epoxi-filoquinona no soro (Bach et al., 1996, Camilo et al., 1998).
DEFICIÊNCIA DE VITAMINA K
Diversos fatores, já discutidos, protegem os adultos da deficiência de vitamina K, como: a distribuição ampla de vitamina K nos alimentos, o ciclo endógeno da vitamina e a própria flora intestinal. Serão discutidos a seguir, as principais manifestações de deficiência e suas possíveis causas.
- Deficiência Subclínica: é geralmente aceito que a ingestão diária de 1 mg/Kg de peso corporal é necessária para garantir a carboxilação dos vários fatores de coagulação. Porém, a definição de deficiência de vitamina K depende fortemente do marcador a ser utilizado na avaliação. Vermeer & Hamulyák (1991) consideram o estado de deficiência como aquele onde pelo menos uma proteína-Gla encontra-se descarboxilada, podendo ser revertida pela administração extra de vitamina K. A partir daí e considerando que a carboxilação completa das proteínas-Gla ósseas requer maiores quantidades de vitamina K, parte substancial da população pode ser considerada bioquimicamente deficiente em vitamina K (deficiência subclínica da vitamina).
- Manifestação hemorrágica: a hemostasia normal depende de interações entre vasos sangüíneos, os elementos figurados do sangue e as proteínas da coagulação sangüínea. Estados de deficiência de vitamina K com hipoprotrombinemia podem produzir o prolongamento do TP e estão associados a um risco aumentado de hemorragias (Guerra & Rosenfeld, 1995).
- Osteoporose: a deficiência dietética de vitamina K e seu antagonismo podem provocar a descarboxilação parcial ou total da osteocalcina, importante proteína da matriz óssea, já citada. A concentração circulante de osteocalcina tem sido apontada como indicador de risco de fratura de quadril (Szulc et al., 1996). Alguns estudos avaliaram diretamente o estado de vitamina K em indivíduos osteoporóticos. Hodges et al. (1993) observaram níveis reduzidos de filoquinona e menaquinonas no plasma e no osso de mulheres idosas com fraturas de quadril. Knapen et al. (1989) compararam as concentrações de osteocalcina de mulheres em pré e pós-menopausa. Foi observado que mulheres em pós-menopausa apresentavam osteocalcina pouco-carboxilada em torno de 40% do total, em relação às mulheres em pré-menopausa. As mulheres em pós-menopausa responderam à suplementação com filoquinona, com aumento da osteocalcina total carboxilada, decréscimo na excreção de cálcio urinário e de hidroxiprolina. Mais recentemente, Booth et al. (2000) concluíram, estudando homens e mulheres idosas, participantes do estudo Framingham, que a ingestão baixa de vitamina K está associada ao aumento da incidência de fraturas de quadril; contudo, não foi observada associação com baixa densidade mineral óssea. Apesar das limitações observadas e discutidas nesses estudos epidemiológicos, eles parecem concordantes no sentido de reconhecer o papel da vitamina K no retardo da perda óssea em pessoas idosas (Olson, 2000).
Entre as principais causas de deficiência de vitamina K destacam-se:
- Inadequação Dietética: embora a deficiência primária de vitamina K seja rara na população saudável, pode ocorrer naqueles indivíduos que apresentam baixa ingestão da vitamina associado ao uso de determinados medicamentos (Suttie, 1992).
- Doença Hemorrágica do recém-nato: a doença hemorrágica dos recém-natos é uma síndrome bem reconhecida, relacionada à deficiência de vitamina K (Lane & Hathaway, 1985). Fatores como imaturidade hepática, luz intestinal estéril e baixo conteúdo de vitamina K no leite materno são contribuintes para a deficiência da vitamina nessa população (Andrew et al., 1987; Canfield & Hopkinson, 1989). Fórmulas comerciais para lactentes são atualmente suplementadas com a vitamina K e a Academia Americana de Pediatria recomenda a administração intramuscular de filoquinona ao nascimento, como profilaxia rotineira (Suttie, 1996).
- Medicamentos: além dos medicamentos cumarínicos (drogas anticoagulantes), alguns antibióticos, como já discutido anteriormente, por exemplo as cefalosporinas, podem inibir diretamente a enzima epóxido-redutase hepática, causando antagonismo à vitamina (Olson, 1999).
- Nutrição Parenteral Total (NPT): a deficiência de vitamina K tem sido observada em indivíduos submetidos à NPT durante longos períodos (Carlin & Walker, 1991).
- Alterações da absorção intestinal: Síndrome de má absorção e obstrução biliar também são conhecidas e possíveis causas de deficiência de vitamina K, que respondem à suplementação vitamínica (Olson, 1999).
- Megadoses de vitaminas A e E: megadoses de vitaminas liposolúveis A e E antagonizam a vitamina K. Tem sido reconhecido desde 1944 que hipervitaminose A no rato leva à hipoprotrombinemia que pode ser revertida pela administração de vitamina K. Acredita-se que a vitamina A reduza a absorção da vitamina K (Olson, 1999). Com relação à vitamina E, há referência à potencialização da atividade da varfarina associada à administração de doses elevadas desta vitamina, acima de 1200UI (Anonymous, 1982).
Recomendações dietéticas
Os estudos quantitativos para determinação dos requerimentos médios de vitamina K são poucos. Além disso, a interpretação dos achados é difícil devido a vários fatores que incluem o baixo requerimento total do nutriente, dificuldade de dosar precisamente a vitamina nos alimentos e nos tecidos, a quantidade notável da vitamina em diversos alimentos e a imprecisão do método habitualmente empregado para avaliar o estado de vitamina K, o tempo de protrombina (TP) simples (Suttie, 1992).
As recomendações dietéticas (Recommended Dietary Allowances - RDA) para a vitamina K foram estabelecidas pela Food and Nutrition Board para crianças, homens e mulheres adultos pela primeira vez na sua décima edição (1989), tal como exposto no Tabela 2. As recomendações são definidas como o nível de ingestão do nutriente que, com base no conhecimento científico, se julga como sendo adequado para alcançar as necessidades de todas as pessoas sadias (Olson, 1999). Entretanto, até o momento, os requerimentos humanos de vitamina K são baseados somente na sua clássica função na coagulação. A ingestão usual de vitamina K em países ocidentais é estimada em 150-500 mg diariamente (Olson, 1994), quantidade essa bem acima do requerimento dietético estabelecido. Booth et al. (1996b) realizaram estudo para estimar o consumo de vitamina K da dieta da população americana. Foi observado que, de 14 grupos separados por sexo e idade, o único que apresentou ingestão abaixo da RDA foi o grupo de adultos jovens de ambos os sexos. Todos os demais grupos consumiam quantidades que atingiam as recomendações, porém abaixo de 90 mg /dia. Em contraste, a ingestão estimada de vitamina K1, nas fórmulas infantis, foi cerca de seis vezes maior que as quantidades recomendadas pela RDA. Foi descrito, ainda, no mesmo trabalho, consumo médio mais elevado de vitamina K1 no grupo mais idoso, tanto em homens quanto em mulheres, embora dentro das recomendações estabelecidas.
Nos bancos de dados pesquisados não foram encontrados trabalhos em que a ingestão de vitamina K na população brasileira tenha sido avaliada. Nosso laboratório estimou a ingestão de vitamina K numa população de 115 pacientes (62 homens e 53 mulheres) acompanhados em ambulatório especializado. Observou-se ingestão média de vitamina K (por kg/peso) acima da RDA, tanto nos grupos de homens quanto de mulheres, quanto nos grupos com idade ³ e < do que 65 anos. Em 23% desses pacientes houve uma ingestão abaixo dos valores recomendados pela RDA de 1989, ou seja, abaixo de 1 mg/kg de peso corporal (Dôres et al., 2000).
Tem sido reconhecido que as necessidades de vitamina K podem ser suficientes para manter normal a coagulação sangüínea, porém são sub-ótimas para o osso (Kohlmeier et al., 1996). Um estudo conduzido em 1997, em unidade metabólica, com nove pacientes mostrou que dieta suplementada com vitamina K (420 mg/dia) produziu declínio nas concentrações de ucOc, dentro de período de 5 dias, em média de 41%. Este fato sugere que ingestões "normais" de vitamina K, na população americana (100 mg/dia), não são suficientes para promover a máxima carboxilação de todas as proteínas dependentes de vitamina K (Sokoll et al., 1997). Se, como argumentado por diversos autores, as necessidades de vitamina K para função óssea são maiores, o grande desafio para pesquisadores e comitês futuros será determinar se essa demanda "extra" pode ser quantificada precisamente (Shearer et al., 1996).
CONSIDERAÇÕES FINAIS
Analisando os estudos relatados nesta revisão, podemos fazer as seguintes considerações:
- O conhecimento do mecanismo exato da função da vitamina K possibilitou a descoberta do seu papel no metabolismo ósseo, além da sua clássica função na coagulação sanguínea. A atuação da vitamina em tecidos extra hepáticos não é considerada na recomendação dietética da RDA de 1989.
- As menaquinonas têm distribuição na dieta mais restrita do que a filoquinona. Um grande questionamento, entretanto, refere-se à utilização do pool de menaquinonas produzidas pela flora intestinal.
- Os grupos de alimentos que mais contribuem para suprir as necessidades de vitamina K são os grupos dos óleos e gorduras e das hortaliças, sendo juntos, responsáveis por até 80% da ingestão total. O feijão foi identificado como importante fonte alimentar de vitamina K na dieta de um grupo de pacientes brasileiros.
- A biodisponibilidade da vitamina K é influenciada por fatores nutricionais e digestivos e a adição de gordura à dieta total favorece a biodisponibilidade da vitamina.
- As concentrações plasmáticas de vitamina K, em jejum, são extremamente baixas e fatores como sexo, idade e polimorfismo genético da apolipoproteína E, afetam o metabolismo da vitamina K
- A relação entre a ingestão dietética de vitamina K e o estado nutricional, relacionado à vitamina, tem sido inadequadamente estudada em função da limitada quantidade de tabelas de composição de alimentos disponível. Atualmente, a utilização de cromatografia (CLAE) vem facilitando determinações rotineiras de vitamina K; contudo, ainda não se dispõe de tabelas com a dosagem de filoquinona de alimentos brasileiros.
- O antagonismo à vitamina K produzido pela administração de drogas cumarínicas comparado à inadequação dietética, parece afetar mais intensamente os marcadores nutricionais da vitamina, em especial, a osteocalcina descarboxilada.
- Atualmente, dispomos de novos e melhores indicadores do estado nutricional, relacionado à vitamina, como o estado de carboxilação das proteínas dependentes da vitamina K.
- Quanto à ingestão na população norte-americana, esta mostra-se suficiente para atingir os requerimentos de acordo com a RDA de 1989, mas não para permitir a máxima carboxilação das proteínas dependentes da vitamina K. Assim, a deficiência subclínica de vitamina K parece ser uma realidade para parte da população dos Estados Unidos da América. Vale ressaltar que a definição de deficiência de vitamina K depende fortemente do marcador avaliado.
- Os dados da literatura aqui analisados ainda não são conclusivos a respeito da quantidade ideal de vitamina K para a saúde humana; entretanto, apontam para a necessidade da realização de novos estudos, com o propósito de revisão imediata das atuais recomendações.
Recebido para publicação em 16 de agosto e aceito em 23 de novembro de 2000.
- ALLISON, P.M., MUMMAH-SCHENDEL, L.L., KINDBERG, C.G., HARMS, C.S., BANG, N.U., SUTTIE, J.W. Effects of a vitamin K-deficient diet and antibiotics in normal human volunteers. Journal of Laboratory Clinical Medicine, St. Louis, v.110, n.2, p.180-188, 1987.
- ANDREW, M., PAES, B., MILNER, R. Development of the human coagulation system in the full-term infant. Blood, Duluth, v.70, n.1, p.165-172, 1987.
- ANONYMOUS. Vitamin K, vitamin E and the coumarin drugs. Nutrition Reviews, New York, v. 40, n.6, p.180-182, 1982.
- BACH, A.U., ANDERSON, S.A., FOLEY, A.L., WILLIAMS, E.C. , SUTTIE, J.W. Assessment of vitamin K status in human subjects administered "minidose" warfarin. American Journal of Clinical Nutrition, Bethesda, v.64, n.6, p.894-902, 1996.
- BERKNER, K.L. The vitamin K-dependent carboxylase. Journal of Nutrition, Bethesda, v.130, n.8, p.1877-1880, 2000.
- BOOTH, S.L., SADOWSKI, J.A., WEIHRAUCH, J.L., FERLAND, G. Viatmin K1 (phylloquinone) content of foods: a provisional table. Journal of Food Composition and Analysis, Blacksburg, v.6, p.109-120, 1993.
- BOOTH, S.L., MADABUSHI, H.T., DAVIDSON, K.W. , SADOWSKI, J.A. Tea and coffee brews are not dietary sources of vitamin K-1 (phylloquinone). Journal of the American Dietetic Association, Chicago, v.95, n.1, p.82-83, 1995a.
- BOOTH, S.L., SOKOLL, L.J., O'BRIEN, M.E., TUCKER, K., DAWSON-HUGHES, B., SADOWSKI, J.A. Assessment of dietary phylloquinone intake and vitamin K status in postmenopausal women. European Journal of Clinical Nutrition, London, v.49, n.11, p.832-841, 1995b.
- BOOTH, S.L., PENNINGTON, J.A., SADOWSKI, J.A. Dihydro-vitamin K1: primary food sources and estimated dietary intakes in the American diet. Lipids, Champaign, v.31, n.7, p.715-720, 1996a.
- BOOTH, S.L., PENNINGTON, J.A., SADOWSKI, J.A. Food sources and dietary intakes of vitamin K-1 (phylloquinone) in the American diet: data from the FDA Total Diet Study. Journal of the American Dietetic Association, Chicago, v.96, n.2, p.149-154, 1996b.
- BOOTH, S.L., TUCKER, K.L., McKEOWN, N.M., DAVIDSON, K.W., DALLAL, G.E., SADOWSKI, J.A. Relationships between dietary intakes and fasting plasma concentrations of fat-soluble vitamins in humans. Journal of Nutrition, Bethesda, v.127, n.4, p.587-592, 1997.
- BOOTH, S.L. , SUTTIE, J.W. Dietary intake and adequacy of vitamin K. Journal of Nutrition, Bethesda, v.128, n.5, p.785-788, 1998.
- BOOTH, S.L., TUCKER, K.L., CHEN, H., HANNAN, M.T., GAGNON, D.R., CUPPLES, L.A., WILSON, P.W.F., ORDOVAS, J., SCHAEFER, E.J., DAWSON-HUGHES, B., KIEL, D.P. Dietary vitamin K intakes are associated with hip fracture but not with bone mineral density in elderly men and women. American Journal of Clinical Nutrition, Bethesda, v.71, n.5, p.1201-1208, 2000.
- CAMILO, M.E., PAIVA, S.A.R., O'BRIEN, M., BOOTH, S.L., DAVIDSON, K.W., SOKOLL, L., SADOWSKI, J.A., RUSSELL, M.R. The interaction between vitamin K nutriture and warfarin administration in patients with bacterial overgrowth due to atrophic gastritis. The Journal of Clinical Nutrition Health and Aging, v.2, n.2, p.716-721, 1998.
- CANFIELD, L.M., HOPKINSON, J.M. State of the art vitamin K in human milk. Journal of Pediatric Gastroenterology and Nutrition, New York, v.8, n.4, p.430-441, 1989.
- CARLIN, A., WALKER, W.A. Rapid development of vitamin K deficiency in an adolescent boy receiving total parenteral nutrition following bone narrow transplantation. Nutrition Reviews, New York, v.49, n.6, p.179-183, 1991.
- DAVIDSON, K.W., BOOTH, S.L., DOLNIKOWSKI, G.G., SADOWSKI, J.A. Conversion of vitamim K1 to 2',3'-dihydrovitamin K1 during the hydrogenation of vegetable oils. Journal of Agricultural and Food Chemistry, Washington DC, v.44, p.980-983, 1996.
- DAVIDSON, K.W., SADOWSKI, J.A. Determination of vitamin K compounds in plasma or serum by HPLC using post colum chemical reduction and fluorometric detection. In: McCORNICK, D.B., SUTTIE, J., WAGNER, C. (Ed.). Vitamins and coenzymes: methods in enzymology. San Diego : Academic Press, 1997. p.408-421.
- DÔRES, S.M.C., FREITAS Jr. I.F., PAIVA, S.A.R., MAFFEI, F.H.A., CAMPANA, A.O. Estimativa da ingestão usual e fontes dietéticas de vitamina K1 em pacientes ambulatoriais. In: CONGRESSO LATINO-AMERICANO DE NUTRIÇÃO, 12., 2000, Buenos Aires. Anais.. Buenos Aires, 2000. p.EN-33.
- DOWD, P., HERSHLINE, R., HAM, S.W., NAGANATHAN, S. The mechanism of action of vitamin K. Annual Review of Nutrition, Palo Alto, v.15, p.419-440, 1995.
- DUELLO, T.J., MATSCHINER, J.T. Identification of phylloquinone in horse liver. Archives of Biochemistry and Biophysics, New York, v.138, n.2, p.640-645, 1970.
- FENTON, S.T., PRICE, R.J., BOLTON-SMITH, C., HARRINGTON, D., SHEARER, M.J. Nutrient sources of phylloquinone in Scottish men and women. Proceedings of the Nutrition Society, London, v.56, p.301, 1997.
- FERLAND, G., SADOWSKI, J.A. The vitamin K1 (phylloquinone) content of edible oils: effects of heating and light exposure. Journal of Agricultural and Food Chemistry, Washington DC, v.40, p.1869-1873, 1992.
- FERLAND, G., SADOWSKI, J.A, O'BRIEN, M.E. Dietary induced subclinical vitamin K deficiency in normal human subjects. Journal of Clinical Investigation, New York, v.91, n.4, p.1761-1768, 1993.
- GABER, A.K., BINKLEY, N.C., KRUEGER, D.C., SUTTIE, J.W. Comparison of phylloquinone bioavailability from food sources or a supplement in human subjects. Journal of Nutrition, Bethesda, v.129, n.6, p.1201-1203, 1999.
- GIJSBER, B.L.; VAN HAARLEN L.J.M; EBBERINK, R.H; VERMEER, C. Caracterization of a Gla containing protein from calcified human atherosclerotic plaques. Artherosclerosis, Dallas, v.10, n.6, p.991-995, 1990.
- GIJSBERS, B.L., JIE, K.S. , VERMEER, C. Effect of food composition on vitamin K absorption in human volunteers. British Journal of Nutrition, London, v.76, n.2, p.223-229, 1996.
- GUERRA, C.C.C., ROSENFELD, L.G.M. Coagulação e anticoagulantes. In: MAFFEI, F.H.A., LASTÓRIA, S., YOSHIDA, W.B., ROLLO. Doenças vasculares periféricas [s.l] : Médica Científica, 1995. p.47-65.
- HIRAUCHI, K., SAKANO, T., NOTSUMOTO, S., NAGAOKA, T., MORIMOTO, A., FUJIMOTO, K., MASUDA, S. , SUZUKI, Y. Measurement of K vitamins in animal tissues by high-performance liquid chromatography with fluorimetric detection. Journal of Chromatography, Amsterdam, v.497, p.131-137, 1989.
- HODGES, S.J., BEJUI, J., LECLERCQ, M., DELMAS, P.D. Detection and measurement of vitamins K1 and K2 in human cortical and trabecular bone. Journal of Bone Mineral Research, New York, v.8, n.8, p.1005-1008, 1993.
- KINDBERG, C.G., SUTTIE, J.W. Effect of various intakes of phylloquinone on signs of vitamin K deficiency and serum and liver phylloquinone concentrations in the rat. Journal of Nutrition, Bethesda, v.119, n.2, p.175-180, 1989.
- KNAPEN, M.J., HAMULYÁK, K., VERMEER, M.H.J. The effect of vitamin K supplementation on circulating osteocalcin (bone Gla protein) and urinary calcium excretion. Annual Internal Medicine, v.111, n.12, p.1001-1005, 1989.
- KOHLMEIER, M., SALOMON, A., SAUPE, J., SHEARER, M.J. Transport of vitamin K to bone in humans. Journal of Nutrition, Bethesda, v.126, n.4, p.1192S-1196S, 1996. Supplement.
- KOHLMEIER, M., SAUPE, J., DROSSEL, H.J., SHEARER, M.J. Variation of phylloquinone (vitamin K1) concentrations in hemodialysis patients. Thrombosis Haemostasis, Stuttgart, v.74, n.5, p.1252-1254, 1995.
- LAMON-FAVA, S., SADOWSKI, J.A., DAVIDSON, K.W., O'BRIEN, M.E., McNAMARA, J.R., SCHAEFER, E.J. Plasma lipoproteins as carriers of phylloquinone (vitamin K1) in humans. American Journal of Clinical Nutrition, Bethesda, v.67, n.6, p.1226-1231, 1998.
- LANE, PA., HATHAWAY, W.E. Vitamin K in infancy. Journal of Pediatrics, St. Louis, v.106, n.3, p.351-359, 1985.
- LANGENBERG, J.P., TJADEN, U.R., De VOGEL, E.M., LANGERAK, D.I. Determination of vitamin K1 in raw and processed vegetables using reversed phase HPLC with electrofluorometric detection. Acta Alimentaria, v.15, n.3, p.187-198, 1986.
- MIJARES, M.E., NAGY, E., GUERRERO, B., AROCHA-PIÑANGO, C.L. La vitamina K: bioquímica, función y deficiencias. Revisión. Investigation Clinics, v.39, n.3, p.213-229, 1998.
- OLSON, R.E. The function and metabolism of vitamin K. Annual Review of Nutrition, Palo Alto, v.4, p.281-337, 1984.
- OLSON, R.E. Vitamin K. In: SHILS, M.E., A. OLSON, J.A., SHIKE, M. Modern nutrition in health and disease [s.l] : Lea & Febiger, 1994. p.342-358.
- OLSON, R.E. Vitamin K. In: SHILS, M.E., OLSON, J.A., SHIKE, M., ROSS, A.C. Modern nutrition in health and disease. Baltimore : Williams & Wilkins, 1999. p.363-380.
- OLSON, R.E. Osteoporosis and vitamin K intake. American Journal of Clinic Nutrition, Bethesda, v.71, p.1031-1032, 2000.
- PAIVA, S.A., SEPE, T.E., BOOTH, S.L., CAMILO, M.E., O'BRIEN, M.E., DAVIDSON, K.W., SADOWSKI, J.A., RUSSELL, R.M. Interaction between vitamin K nutriture and bacterial overgrowth in hypochlorhydria induced by omeprazole. American Journal of Clinical Nutrition, Bethesda, v.68, n.3, p.699-704, 1998.
- SADOWSKI, J.A., HOOD, S.J., DALLAL, G.E., GARRY, P.J. Phylloquinone in plasma from elderly and young adults: factors influencing its concentration. American Journal of Clinical Nutrition, Bethesda, v.50, n.1, p.100-108, 1989.
- SADOWSKI, J.A., BOOTH, S.L., MANN, K.M., MALHOTRA, O., BOVILL, E.G. Structure and mechanism of activation of vitamin K antagonist. In: POLLER, L., HIRSH, J. Oral anticoagulants New York : Arnold, 1996.
- SAKANO, T., NOTSUMOTO, M., NAGAOKA, T., MORIMOTO, A., FUJIMOTO, K., MASUDA, S., SUZUKI, Y., HIRAUCHI, K. Measuraments of K vitamins by highperformance liquid chromatography with fluorometric detection. Vitamins, Kyoto, v.62, p.393-398, 1988.
- SHEARER, M.J., McBURNEY, A., BARKHAN, P. Studies on the absorption and metabolism of phylloquinone (vitamin K1) in man. Vitamins and Hormones, New York, v.32, p.513-542, 1974.
- SHEARER, M.J., BECHTOLD, H., ANDRASSY, K., KODERISCH, J., McCARTHY, P.T., TRENK, D., JAHNCHEN, E., RITZ, E. Mechanism of cephalosporin-induced hypoprothrombinemia: relation to cephalosporin side chain, vitamin K metabolism, and vitamin K status. Journal of Clinical Pharmacology, Hagerstown, v.28, n.1, p.88-95, 1988.
- SHEARER, M.J. Vitamin K. Lancet, London, v.345, n.8944, p.229-234, 1995.
- SHEARER, M.J., BACH, A., KOHLMEIER, M. Chemistry, nutritional sources, tissue distribution and metabolism of vitamin K with special reference to bone health. Journal of Nutrition, Bethesda, v.126, n.4, p.1181S-1186S, 1996.
- SOKOLL, L., O'BRIEN, M., CAMILO, M., SADOWSKI, J. Undercarboxylated osteocalcin and development of a method to determine vitamin K status. Clinical Chemistry, Washington DC, v.41, n.8, p.1121-1128, 1995.
- SOKOLL, L.J., BOOTH, S.L., O'BRIEN, M.E., DAVIDSON, K.W., TSAIOUN, K.I., SADOWSKI, J.A. Changes in serum osteocalcin, plasma phylloquinone and urinary gamma- carboxyglutamic acid in response to altered intakes of dietary phylloquinone in human subjects. American Journal of Clinical Nutrition, Bethesda, v.65, n.3, p.779-784, 1997.
- SOKOLL, L.J., SADOWSKI, J.A. Comparison of biochemical indexes for assessing vitamin K nutritional status in a healthy adult population. American Journal of Clinical Nutrition, Bethesda, v.63, n.4, p.566-573, 1996.
- SUTTIE, J.W. Vitamin K and human nutritrion. Journal of the American Dietetic Association, Chicago, v.92, n.5, p.585-590, 1992.
- SUTTIE, J.W. The importance of menaquinones in human nutrition. Annual Review of Nutrition, Palo Alto, v.15, p.399-417, 1995.
- SUTTIE, J.W. Vitamin K. In: ZIEGLER, E.E., FILER Jr. L.J. Present knowledge in nutrition. Washington : ILSI Press, 1996. p.137-145.
- SUTTIE, J.W., MUMMAH-SCHENDEL, L.L., SHAH, D.V., LYLE, B.J., GREGER, J.L. Vitamin K deficiency from dietary vitamin K restriction in humans. American Journal of Clinical Nutrition, Bethesda, v.47, n.3, p.475-480, 1988.
- SZULC, P., CHAPUY, M.C., MEUNIER, P.J., DELMAS, P.D. Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture: a three year follow-up study. Bone, v.18, n.5, p.719-724, 1996.
- THIJSSEN, H.H., DRITTIJ-REIJNDERS, M.J. Vitamin K status in human tissues: tissue-specific accumulation of phylloquinone and menaquinone-4. British Journal of Nutrition, London, v.75, n.1, p.121-127, 1996.
- USUI, Y., TANIMURA, H., NISHIMURA, N., KOBAYASHI, N., OKANOUE, T., OZAWA, K. Vitamin K concentrations in the plasma and liver of surgical patients. American Journal of Clinical Nutrition, Bethesda, v.51, n.5, p.846-852, 1990.
- VERMEER, C., GIJSBERS, B.L., CRACIUN, A.M., GROENEN-VAN DOOREN, M.M., KNAPEN, M.H. Effects of vitamin K on bone mass and bone metabolism. Journal of Nutrition, Bethesda, v.126, n.4, p.1187S-1191S, 1996. Supplement.
- VERMEER, C., HAMULYAK, K. Pathophysiology of vitamin K-deficiency and oral anticoagulants. Thrombosis Haemostasis, Stuttgart, v.66, n.1, p.153-159, 1991.
- VERMEER, C., JIE, K.S., KNAPEN, M.H. Role of vitamin K in bone metabolism. Annual Review of Nutrition, Palo Alto, v.15, p.1-22, 1995.
Datas de Publicação
-
Publicação nesta coleção
15 Fev 2002 -
Data do Fascículo
Dez 2001
Histórico
-
Aceito
23 Nov 2000 -
Recebido
16 Ago 2000