Resumos
O estudo objetivou avaliar translocação orgânica, índices fisiológicos da análise de crescimento e rendimento do óleo essencial de Mentha piperita L. cultivada em solução nutritiva com variação dos níveis de N, P, K e Mg. Assim, foram avaliados os quatro tratamentos contendo 50% N, P, K, 25% Mg; 50% N, P, K, Mg; 65%N, 50%P, 25%K, 100% Mg e 100% N, P, K, Mg. A translocação orgânica foi avaliada por meio da determinação da razão de massa foliar (RMF) e da distribuição de massa seca para os diferentes órgãos. Os índices fisiológicos razão de área foliar (RAF), área foliar específica (AFE), taxa assimilatória líquida (TAL) e taxa de crescimento relativo (TCR) derivadas que compõem a análise de crescimento foram estimados pelo programa ANACRES, após ajuste exponencial quadrático da área foliar e massa seca de lâminas foliares e total da planta em relação ao tempo. O rendimento do óleo essencial, em porcentagem, foi calculado após extração da parte aérea por hidrodestilação. As plantas submetidas ao tratamento com nível completo de nutrientes (100%N/P/K/Mg) exportaram com menor eficiência o material orgânico a partir da folha e a RMF mostrou queda mais lenta, devido à retenção desse material por mais tempo no local de sua produção. Além disso, não apresentaram melhor produtividade e as curvas da TAL e TCR mostraram quedas mais lentas. As plantas submetidas ao tratamento com 65%N/50%P/25%K/100%Mg revelaram adequada exportação de matéria orgânica da folha para caule e pecíolos, conforme demonstra a RMF e a distribuição de massa seca para esses órgãos. Revelaram ainda a RAF mais elevada no inicio do desenvolvimento e mais baixa aos 94 DAT, indicando sombreamento das folhas como resultado de seu crescimento. A AFE dessas plantas mostrou menor variação durante o ciclo, refletindo espessura mais constante de suas folhas. A TAL e a TCR apresentaram curvas decrescentes, com quedas bem acentuadas em relação às demais plantas. Todos esses índices indicam melhor produtividade das plantas submetidas ao tratamento 65%N/50%P/25%K/100%Mg, que também apresentaram rendimento de óleo essencial, em média, maior durante o ciclo. Conclui-se que a redução de 35% de N, 50% de P, 75% de K (65%N/50%P/25%K/100%Mg) é indicada para o cultivo e extração de óleo essencial de M. piperita.
Mentha piperita; óleo essencial; nutrição mineral; hidroponia
This study aimed to evaluate organic translocation, physiologic indexes of growth analysis and essential oil yield of Mentha piperita L. cultivated in nutrient solution with varying levels of N, P, K and Mg. Thus, four treatments were evaluated: 50% N, P, K, 25% Mg; 50% N, P, K, Mg; 65% N, 50% P, 25% K, 100% Mg; and 100% N, P, K, Mg. Organic translocation was assessed through leaf mass ratio (LMR) and dry matter distribution to the different organs. The physiologic indexes leaf area ratio (LAR), specific leaf area (SLA), net assimilation rate (NAR) and relative growth rate (RGR), derived from growth analysis, were estimated using the software ANACRES, following the quadratic exponential fitting of leaf area and dry mass of leaves and total plant, relative to time. Essential oil yield was calculated as percentage after extraction from shoot using hydrodistillation. Plants subjected to the treatment with complete level of nutrients (100%N/P/K/Mg) less efficiently exported organic material from the leaf and LMR showed slower decrease due to the longer retention of such material in its production site. Besides, those plants did not present higher productivity and NAR and RGR curves showed slower decrease. Plants subjected to the treatment with 65%N/50%P/25%K/100%Mg had adequate organic matter exportation from the leaves to the stem and petioles, as demonstrated by LMR and dry matter distribution to these organs. Also in these plants, LAR was higher at the initial development and lower at 94 DAT, indicating leaf shading as a result of growth, and SLA showed less variation during the cycle, reflecting more homogeneous leaf thickness. NAR and RGR had decreasing curves, with marked falls relative to the remaining plants. All these indexes indicate higher productivity of plants subjected to the treatment with 65%N/50%P/25%K/100%Mg, which also had higher essential oil yield, on average, during the cycle. In conclusion, 35% reduction for N, 50% for P, 75% for K (65%N/50%P/25%K/100%Mg) is recommended for cultivation and essential oil extraction of M. piperita.
Mentha piperita; essential oil; mineral nutrition; hydroponics
Translocação orgânica, produtividade e rendimento de óleo essencial de Mentha piperita L. cultivada em solução nutritiva com variação dos níveis de N, P, K e Mg
Organic translocation, productivity and essential oil yield of Mentha piperita L. cultivated in nutrient solution with varying N, P, K and Mg levels
David, E.F.S.I, * * evelizedavid@yahoo.com.br ; Boaro, C.S.F.II
IFaculdade de Ciências Agronômicas, Departamento de Produção Vegetal/Horticultura, FCA-UNESP. Caixa Postal 237, CEP 18603-970, Botucatu-SP
IIInstituto de Biociências, Departamento de Botânica, Caixa Postal 510. UNESP-Botucatu-SP
RESUMO
O estudo objetivou avaliar translocação orgânica, índices fisiológicos da análise de crescimento e rendimento do óleo essencial de Mentha piperita L. cultivada em solução nutritiva com variação dos níveis de N, P, K e Mg. Assim, foram avaliados os quatro tratamentos contendo 50% N, P, K, 25% Mg; 50% N, P, K, Mg; 65%N, 50%P, 25%K, 100% Mg e 100% N, P, K, Mg. A translocação orgânica foi avaliada por meio da determinação da razão de massa foliar (RMF) e da distribuição de massa seca para os diferentes órgãos. Os índices fisiológicos razão de área foliar (RAF), área foliar específica (AFE), taxa assimilatória líquida (TAL) e taxa de crescimento relativo (TCR) derivadas que compõem a análise de crescimento foram estimados pelo programa ANACRES, após ajuste exponencial quadrático da área foliar e massa seca de lâminas foliares e total da planta em relação ao tempo. O rendimento do óleo essencial, em porcentagem, foi calculado após extração da parte aérea por hidrodestilação. As plantas submetidas ao tratamento com nível completo de nutrientes (100%N/P/K/Mg) exportaram com menor eficiência o material orgânico a partir da folha e a RMF mostrou queda mais lenta, devido à retenção desse material por mais tempo no local de sua produção. Além disso, não apresentaram melhor produtividade e as curvas da TAL e TCR mostraram quedas mais lentas. As plantas submetidas ao tratamento com 65%N/50%P/25%K/100%Mg revelaram adequada exportação de matéria orgânica da folha para caule e pecíolos, conforme demonstra a RMF e a distribuição de massa seca para esses órgãos. Revelaram ainda a RAF mais elevada no inicio do desenvolvimento e mais baixa aos 94 DAT, indicando sombreamento das folhas como resultado de seu crescimento. A AFE dessas plantas mostrou menor variação durante o ciclo, refletindo espessura mais constante de suas folhas. A TAL e a TCR apresentaram curvas decrescentes, com quedas bem acentuadas em relação às demais plantas. Todos esses índices indicam melhor produtividade das plantas submetidas ao tratamento 65%N/50%P/25%K/100%Mg, que também apresentaram rendimento de óleo essencial, em média, maior durante o ciclo. Conclui-se que a redução de 35% de N, 50% de P, 75% de K (65%N/50%P/25%K/100%Mg) é indicada para o cultivo e extração de óleo essencial de M. piperita.
Palavras-chave:Mentha piperita, óleo essencial, nutrição mineral, hidroponia
ABSTRACT
This study aimed to evaluate organic translocation, physiologic indexes of growth analysis and essential oil yield of Mentha piperita L. cultivated in nutrient solution with varying levels of N, P, K and Mg. Thus, four treatments were evaluated: 50% N, P, K, 25% Mg; 50% N, P, K, Mg; 65% N, 50% P, 25% K, 100% Mg; and 100% N, P, K, Mg. Organic translocation was assessed through leaf mass ratio (LMR) and dry matter distribution to the different organs. The physiologic indexes leaf area ratio (LAR), specific leaf area (SLA), net assimilation rate (NAR) and relative growth rate (RGR), derived from growth analysis, were estimated using the software ANACRES, following the quadratic exponential fitting of leaf area and dry mass of leaves and total plant, relative to time. Essential oil yield was calculated as percentage after extraction from shoot using hydrodistillation. Plants subjected to the treatment with complete level of nutrients (100%N/P/K/Mg) less efficiently exported organic material from the leaf and LMR showed slower decrease due to the longer retention of such material in its production site. Besides, those plants did not present higher productivity and NAR and RGR curves showed slower decrease. Plants subjected to the treatment with 65%N/50%P/25%K/100%Mg had adequate organic matter exportation from the leaves to the stem and petioles, as demonstrated by LMR and dry matter distribution to these organs. Also in these plants, LAR was higher at the initial development and lower at 94 DAT, indicating leaf shading as a result of growth, and SLA showed less variation during the cycle, reflecting more homogeneous leaf thickness. NAR and RGR had decreasing curves, with marked falls relative to the remaining plants. All these indexes indicate higher productivity of plants subjected to the treatment with 65%N/50%P/25%K/100%Mg, which also had higher essential oil yield, on average, during the cycle. In conclusion, 35% reduction for N, 50% for P, 75% for K (65%N/50%P/25%K/100%Mg) is recommended for cultivation and essential oil extraction of M. piperita.
Key words:Mentha piperita, essential oil, mineral nutrition, hydroponics
INTRODUÇÃO
Os esforços na busca do aumento de produção de óleos essenciais são de grande importância, principalmente quando se considera a dependência da indústria farmacêutica nacional. Atualmente, 90% dos óleos essenciais, destinados ao mercado brasileiro, são importados (Ming,1992). A Associação Brasileira da Indústria de Fitoterápicos estima que o setor movimente R$ 1 bilhão /ano no País (Abifito, 2002).
Considerando tal panorama nacional e também que o cultivo é uma das etapas que mais interfere na produção de matéria - prima vegetal de boa qualidade, com maior teor de óleo essencial, todos os estudos a ele relacionados podem melhorar a produção de compostos do metabolismo secundário de interesse econômico (David, 2004). É o que ocorre com as plantas medicinais e aromáticas, onde, além da biomassa vegetal, devem ser considerados os teores dos princípios ativos produzidos (Box, 1973). O avanço tecnológico e a crescente demanda do mercado consumidor por produtos de qualidade exigem, portanto, do setor agrícola entendimento dos aspectos agronômicos e fisiológicos (Habermann, 1999).
Apesar dessa relevância da área agronômica, as plantas medicinais e aromáticas ainda são pouco avaliadas em pesquisas científicas e várias espécies não têm recebido suficiente atenção sobre formas adequadas de propagação e tratos culturais, com possibilidade de obtenção de melhor rendimento de óleo. Menor número de estudos ainda é verificado no tocante à fisiologia da produção dessas espécies e o produtor acaba realizando empiricamente o manejo da cultura, o que pode resultar em produto de baixa qualidade (Furlan, 2000).
A produtividade das plantas é determinada pela quantidade de folhas, capacidade fotossintética de cada folha e disponibilidade de nutrientes, sendo esses, fatores chaves que definem o ganho de biomassa. O suprimento adequado de nutrientes minerais para as culturas é de fundamental importância para aumento da produtividade (Fageria et al., 1997). Tal importância está fundamentada no fato de que a nutrição mineral influencia direta e indiretamente o metabolismo do carbono devido a ação também no crescimento e morfogênese (Larcher, 2006). Milthorpe & Moorby (1974) referiram que a ausência, ou mesmo a insuficiência, de um nutriente interfere na produção final, embora muitas vezes não se conheça qual das variáveis fisiológicas intermediárias fica comprometida, afetando a produtividade. Muitos também são os estudos que caracterizam a importância do equilíbrio entre os elementos minerais para a nutrição das plantas, constatando por vezes efeitos antagônicos entre os mesmos. Desequilíbrios entre concentrações interferem na disponibilidade, absorção e translocação dos elementos essenciais na planta. Assim, além da disponibilidade de nutrientes, também deve ser considerada a razão elementar adequada entre eles para a obtenção de crescimento ótimo das plantas (Marenco et al., 2001).
Por outro lado, independente das dificuldades inerentes à complexidade que envolve o desenvolvimento, a análise de crescimento é o meio mais simples para avaliá-lo, inferindo a contribuição de diferentes processos fisiológicos sobre o comportamento vegetal (Causton & Venus, 1981). A referida análise descreve as condições morfo-fisiológicas da planta, em diferentes intervalos de tempo, permitindo acompanhar a dinâmica da produtividade, avaliada por variáveis fisiológicas e bioquímicas. É um método que pode ser utilizado para investigação de vários efeitos, entre eles a influência de práticas agronômicas sobre o crescimento (Magalhães, 1986) e baseia-se, fundamentalmente, no fato de que cerca de 90%, em média, da matéria seca acumulada pelas plantas ao longo do crescimento, resulta da atividade fotossintética. O restante é proveniente da absorção de nutrientes minerais, que embora quantitativamente de menor expressão, são indispensáveis ao desenvolvimento vegetal. Como o crescimento é avaliado por meio de variações em tamanho de algum aspecto da planta, geralmente morfológico, em função do acúmulo de material resultante da fotossíntese, esta passa a ser o aspecto fisiológico de maior importância para a análise de crescimento (Benincasa, 2003).
Com base no acima exposto o objetivo do presente estudo foi avaliar translocação orgânica, determinante da produtividade, índices fisiológicos a ela relacionados e rendimento de óleo essencial de plantas de Mentha piperita L., cultivadas em solução nutritiva com variação dos níveis de N, P, K e Mg.
MATERIAL E MÉTODO
O experimento foi conduzido em casa de vegetação do tipo "Paddy-Fan" do Departamento de Botânica do Instituto de Biociências da Universidade Estadual Paulista, Campus Botucatu - SP, entre os meses de março e junho. A temperatura e a umidade relativa na casa de vegetação foram mantidas, respectivamente, em torno de 27ºC e 70%.
As mudas de M. piperita foram obtidas a partir de plantas matrizes provenientes da Escola Superior de Agricultura Luís de Queiroz, ESALQ-USP. Foram propagadas em vasos e posteriormente em canteiros, para a obtenção da quantidade de material suficiente para a execução do experimento. Ramos aéreos dessas plantas foram selecionados e a partir deles, estacas apicais com 10 cm de comprimento e com os quatros primeiros pares de folhas foram confeccionadas. Essas estacas após serem desinfectadas por imersão rápida em solução de hipoclorito de sódio a 2% foram lavadas em água corrente e acondicionadas em solução de nitrato de potássio em concentração de 600 mg L-1 durante 10 dias, tempo necessário para o enraizamento (Soares & Sacramento, 2001).
A seguir, as estacas enraizadas foram transferidas para vasos com volume de 6 L contendo solução nutritiva nº 2 de Hoagland & Arnon (1950), modificada pela variação da concentração de N, P, K e Mg, que constituiu os diferentes tratamentos aos quais as plantas foram submetidas. Assim, o tratamento com 50% de N, P e K e 25% de Mg, (designado 50%N/P/K/25%Mg), continha, em mg L-1, 94,0 N/15,5 P/107,5 K/12,15 Mg, o tratamento com 50% de N, P, K e Mg (50%N/P/K/Mg), 94,0 N/15,5 P/107,5 K/24,3 Mg, o tratamento com 65% de N, 50% de P, 25% de K e 100% de Mg (65%N/50%P/25%K/100%Mg), 124,0 N/15,5 P/53,6 K/48,6 Mg e o tratamento com 100% de N, P, K e Mg (100%N/P/K/Mg), 189,0 N/31,0 P/214,5 K/48,6 Mg. O pH foi mantido entre 5,5 e 6,0 e a condutividade elétrica da solução, conforme as recomendações de Carmello (1992), entre 1,5 e 2,5 mS cm-1. A solução nutritiva, continuamente arejada, com soprador rotativo, foi renovada a cada duas semanas. Sempre que necessário, o volume de solução dos vasos foi completado com água destilada.
O delineamento experimental foi inteiramente casualizado, com quatro repetições, em esquema fatorial 4x4x5, ou seja, quatro tratamentos, quatro repetições com 1 planta por repetição e cinco épocas de colheita, realizadas aos 20, 35, 50, 65 e 85 dias após transplante (DAT), de modo a cobrir o ciclo do vegetal. Para o óleo essencial o delineamento foi inteiramente casualizado em esquema fatorial 4x4x4, ou seja, quatro tratamentos, quatro repetições com 1 planta por repetição com quatro épocas de colheitas, realizadas aos 69, 84, 98 e 113 DAT. Num total de 180 vasos, sendo 100 para a análise dos índices fisiológicos e 80 para o rendimento de óleo essencial.
Seguindo a metodologia utilizada por David et al. (2007), nas cinco colheitas, realizadas aos 20, 35, 50, 65 e 85 DAT, a planta de cada repetição, dos vários tratamentos foi separada em raízes, caules, pecíolos e lâminas foliares. Todos os órgãos, inclusive as lâminas foliares, após a determinação da área em dm2, com auxílio de integralizador de área foliar, modelo LI 3100 da LI-COR, foram acondicionados em sacos de papel etiquetados e colocados para secarem em estufa com circulação forçada de ar, em temperatura entre 60 e 70ºC, até obtenção de massa constante. Após a secagem completa, o material foi pesado em balança analítica Ohaus tipo Analytical Standard com sensibilidade de até 0,1 mg, para determinação da massa seca. Com base na área foliar e massas secas de lâminas foliares e totais das plantas a translocação orgânica foi estimada por meio da razão de massa foliar e da distribuição de massa seca para os diferentes órgãos.
A determinação da razão de massa foliar que é um índice que expressa a fração de matéria seca não exportada das folhas para o resto da planta pode refletir diferenças entre tratamentos quando o mesmo interfere na translocação orgânica. A maior ou menor exportação de material da folha pode ser característica genética, que está sob a influência de variáveis ambientais (Benincasa, 2003). Esta variável foi calculada pela relação entre massa seca da folha (MSF) e massa seca total (MST), ambas em gramas.

A distribuição de massa seca para os diferentes órgãos ou regiões de interesse é calculada em porcentagem de massa seca de cada órgão em relação à massa seca total, ao longo do ciclo ou nas fases de maior interesse. Esta variável tanto quanto a razão de massa foliar (RMF), também permite inferir a translocação orgânica (Benincasa, 2003), permitindo o conhecimento do órgão até onde chega o material orgânico que foi exportado da folha.
Os índices fisiológicos foram calculados, após o ajuste exponencial quadrático da área foliar e da massa seca de lâminas foliares e total das plantas em relação ao tempo, ou seja, idade das plantas. Os índices fisiológicos razão de área foliar (RAF), área foliar específica (AFE), taxa assimilatória líquida (TAL) e taxa de crescimento relativo (TCR) foram estimados pelo programa computacional ANACRES, de acordo com as especificações de Portes & Castro Junior (1991). A escolha da equação de ajuste se baseou nas recomendações dos autores que referem que para espécie de ciclo curto, como a menta, essa equação tem sido adequada, pois é uma das que melhor se ajusta aos conjuntos dos referidos dados, representando bem o crescimento dessa espécie. Assim, a massa seca total e de lâminas foliares e a área foliar, das plantas submetidas aos diferentes tratamentos foram ajustadas em função do tempo, considerando-se a versão lny= a + bt + ct2, para massa seca >0, segundo as equações:
MS(y) = a.e (bt + ct2) e AF(y1) = a1.e (b1t + c1t2)
Os coeficientes foram estimados por análise de regressão, após transformação das equações para a forma logarítmica.
A razão de área foliar, em dm2 g-1, expressa a área foliar útil para a fotossíntese (Benincasa, 2003) e foi obtida a partir dos valores instantâneos de área foliar (AF), responsável pela interceptação de energia luminosa e CO2, e massa seca total, resultado da fotossíntese, segundo a equação:

A área foliar específica, em dm2 g-1, reflete o inverso da espessura da folha (Benincasa, 2003) e foi obtida pela razão entre a área foliar (AF) e a massa seca de folhas (MSF).

A taxa assimilatória líquida, em g dm-2 dia-1, que expressa a taxa de fotossíntese líquida, em termos de massa seca produzida, em gramas, por decímetro quadrado de área foliar, por unidade de tempo (Portes & Castro Junior,1991) foi obtida pela equação:

A taxa de crescimento de uma planta ou de qualquer órgão da planta é uma função do tamanho inicial, isto é, o aumento em gramas, no caso de massa seca, está relacionado ao peso de massa seca no instante em que se inicia o período de observação (Portes & Castro Junior, 1991). Este índice, em g g-1 dia-1, foi calculado pela equação:

Para a determinação do rendimento de óleo essencial as plantas, após terem sido colhidas aos 69, 84, 98 e 113 DAT, foram pesadas e acondicionadas em sacos de papel e colocadas em estufas com circulação forçada de ar, em temperatura de 40ºC, por três dias, até atingir massa seca constante. De acordo com Silva & Casali (2000), a secagem visa impedir a deterioração do material por meio da redução do teor de água, atuando negativamente na ação das enzimas pela desidratação, permitindo assim, a conservação da planta por mais tempo. Além disso, eliminando-se a água, aumenta-se o percentual de princípios ativos em relação à massa.
No terceiro dia foi determinada a massa seca que foi hidrodestilada em aparelho do tipo Clevenger, no Laboratório de Óleos Essenciais do Departamento de Produção Vegetal da FCA - UNESP, Campus Botucatu. Foram colocados 30 gramas de massa seca de parte aérea de M. piperita em balão de fundo chato com capacidade de 2000 mL, adicionando-se água até a cobertura da amostra, que em seguida foi aquecida a temperatura constante, por uma hora e meia. Após a extração, o rendimento do óleo essencial foi calculado com base na diferença entre a massa final do frasco de coleta contendo óleo e a massa inicial, sem óleo. Como essa diferença correspondeu ao óleo extraído de 30 g de massa seca de parte aérea, estimou-se tal rendimento para 100 g de massa.
Os resultados da razão de massa foliar, distribuição de massa seca para os diferentes órgãos e rendimento de óleo essencial foram avaliados estatisticamente por meio de análise de variância do fatorial e de regressão, utilizando-se o nível de 5% de significância (Zar, 1996).
RESULTADO E DISCUSSÃO
De maneira geral, todos os tratamentos apresentaram redução da razão da massa foliar durante o ciclo de desenvolvimento da cultura (Figura 1a), resposta esperada uma vez que à medida que a planta se desenvolve menor é a fração de fotoassimilados retidos na folha, que são direcionados para os demais órgãos para garantir crescimento, havendo consequentemente decréscimo dessa razão, indicativa da translocação orgânica (Radford, 1967; Benincasa, 2003). Lugg & Sinclair (1980) demonstraram que a razão de massa foliar decresce nas fases de aumento da área foliar, ou seja, já na fase de desenvolvimento vegetativo. Assim, os resultados obtidos para esse índice no presente estudo, concordam com os relatados na literatura. Deve-se, no entanto, destacar o comportamento da razão de massa foliar apresentado pelas plantas cultivadas com solução completa 100%N/P/K/Mg, cujo decréscimo apresentou-se mais lento, indicando retenção de fotoassimilados nas folhas por mais tempo. Essa solução mostrou-se, portanto, excessiva para plantas de M. piperita, interferindo com a fase inicial do processo de translocação orgânica, determinante de sua produtividade.
As plantas cultivadas com os diferentes tratamentos não diferiram entre si e apresentaram tendência de decréscimo da distribuição da massa seca para raízes ao longo do ciclo (Figura 1b). Os resultados estão de acordo com as observações de que no início do ciclo de desenvolvimento as plantas investem em produção de raízes para poderem assimilar nutrientes, que são levados até as folhas onde ocorre síntese de fotoassimilados, que são transportadas para as regiões de crescimento, consumo e armazenamento (Radford, 1967; Moraes, 1986; Benincasa, 2003).
David et al. (2007), ao avaliarem M. piperita cultivada com solução nutritiva acrescida e decrescida em de 50% de fósforo, obtiveram, de modo geral, tendência de decréscimo da distribuição da massa seca de raízes ao longo do ciclo, exceção feita ao tratamento com solução nutritiva completa, com 31 mg L-1 de P, aos 80 dias após o transplante, época em que se verificou acentuada tendência de aumento da massa seca para as raízes. Esses resultados embora se refiram somente ao fósforo, indicam a possibilidade de interferência dos nutrientes no processo de translocação orgânica.
Todos os tratamentos apresentaram aumento da distribuição de massa seca para caule mais pecíolos, mostrando que esses órgãos cresceram durante o período de avaliação, exceção feita às plantas nutridas com 100%N/P/K/Mg, que apresentaram redução aos 35 DAT, retornando o crescimento nas colheitas seguintes (Figura 1c). Tal comportamento pode ser justificado devido ao aumento de distribuição de massa seca para lâminas foliares nesse período (Figura 1d) em que as plantas deixaram de investir em caule mais pecíolos para investir em folhas.
Aos 20 DAT, as plantas nutridas com 50%N/P/K/Mg apresentaram distribuição de massa seca para caule e pecíolos menor que as plantas nutridas com 100%N/P/K/Mg, indicando que as primeiras apresentaram menor crescimento desses órgãos no inicio do ciclo de desenvolvimento o que talvez justifique sua menor produtividade. David et al. (2007) obtiveram, de modo geral, tendência de acréscimo da distribuição da massa seca para caule e pecíolos em plantas de M. piperita na maior parte de seu ciclo, nas plantas nutridas com solução completa, com 31 mg L-1 de P. Os resultados sugeriram que quanto maior a concentração de nutrientes, maior o estímulo para crescimento de caule e pecíolos. No entanto, não foram avaliadas as variações de relações entre nutrientes na solução nutritiva, como no presente estudo, em que resultados diferentes indicam que tais relações de fato interferem com o desenvolvimento vegetal.
As plantas apresentaram diminuição de distribuição de massa seca para lâminas foliares ao longo do ciclo (Figura 1d). As plantas cultivadas com 100%N/P/K/Mg, também apresentaram tal diminuição, que, no entanto, foi mais lenta em relação à dos outros tratamentos, indicando maior investimento dessas plantas em fotoassimilados para o desenvolvimento de lâminas foliares. Aos 35 DAT observou-se inclusive discreto aumento na distribuição de massa seca para lâminas foliares nas plantas nutridas com 100%N/P/K/Mg. Esse resultado está de acordo com os de David et al. (2007) que ao avaliarem M. piperita cultivada com solução nutritiva acrescida e decrescida de 50% de fósforo obtiveram, de modo geral, tendência de decréscimo da distribuição da massa seca para lâminas foliares ao longo do ciclo, exceção feita às plantas cultivadas em solução completa, aos 84 dias após o transplante, com acentuada tendência de aumento da massa seca para as lâminas foliares. Esses resultados demonstram para M. piperita estímulo de crescimento de folhas quando os nutrientes são elevados.
De maneira geral, as plantas apresentaram redução da RAF, ao longo do ciclo de desenvolvimento, com exceção das nutridas com 50%N/P/K/25%Mg, que apresentaram aumento desse índice aos 35 DAT e das nutridas com solução completa, ou seja, 100%N/P/K/Mg, que apresentaram aumento dos 20 aos 50 DAT, diminuindo a seguir (Figura 2a). Tal comportamento pode ter ocorrido devido ao maior investimento na produção de folhas para a captação de luz no início do desenvolvimento das plantas submetidas a esses tratamentos, o que pode ser observado na distribuição de massa seca para lâminas foliares (Figura 1d).

Esses resultados são concordantes com os registrados na literatura para a RAF, que expressa a área foliar útil para a fotossíntese e seu comportamento é de ser elevada no inicio do ciclo, período em que ocorre desenvolvimento de folhas para maior captação de luz, decrescendo a seguir devido à interferência de folhas superiores sobre as inferiores, caracterizando auto-sombreamento (Benincasa, 2003), diminuindo assim a área foliar útil para a realização da fotossíntese.
Os resultados observados para RAF estão de acordo também com os encontrados por Boaro (2001), Leal (2001), Valmorbida et al. (2007) e David et al. (2007), que independente de cultivarem espécies diferentes em solução nutritiva com variação de diferentes nutrientes, observaram RAF elevada no início do ciclo vegetativo, decrescendo devido à interferência de folhas superiores sobre as inferiores.
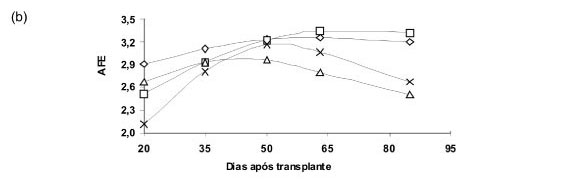
A área foliar específica aumentou, de maneira geral, durante todo o ciclo nas plantas cultivadas com 50% N/P/K/25%Mg e 50% N/P/K/Mg. Nas demais ocorreu aumento desse índice até a metade do período de avaliação, diminuindo a seguir. As plantas cultivadas com 65%N/50%P/25%K/100%Mg foram as que apresentaram a AFE mais constante durante o ciclo (Figura 2b). Segundo Benincasa (2003), no início do desenvolvimento, os valores da AFE podem ser maiores, revelando folhas pouco espessas, com pouca massa seca e área foliar. Com o desenvolvimento da cultura aumenta a área foliar e a massa seca das folhas, tendendo a queda dessa variável. Tal queda, portanto, no presente estudo ocorreu mais tarde nos tratamentos com 50% N/P/K/25%Mg e 50%N/P/K/Mg.
Os resultados observados concordam em parte com os de Valmorbida et al. (2007) que obtiveram aumento da AFE até os 63 DAT, quando cultivaram M.piperita com 100 e 50% de K. Leal (2001) estudou M. piperita cultivada em diferentes níveis de nitrogênio, que dentro de cada tratamento foram alterados a partir dos 42 dias após transplante e observou aumento da AFE nas plantas nutridas com os menores níveis de N, igual a 210 até 42 DAT e 105 mg L-1 a seguir (210/105 mg L-1) e 263/132 mg L-1 de N e diminuição naquelas com o maior nível, 315/158 mg L-1 de N até os 84, dias, demonstrando que doses elevadas de nitrogênio diminuem a AFE, ou seja, resulta em maior espessura de folhas. Tal comportamento pode ser esperado em plantas em que o nitrogênio estimula o crescimento vegetativo.
Esses resultados discordam dos encontrados por David et al. (2007) que observaram que, de maneira geral, a AFE em M. piperita cultivada com diferentes níveis de P diminuiu dos 20 aos 40 DAT, voltando a aumentar a seguir e sendo mais elevada aos 100 DAT. No entanto, os estudos citados acima avaliaram N, P e K separadamente, o que não aconteceu no presente estudo, onde todos os elementos foram avaliados em conjunto e a interação entre eles pode explicar algumas divergências de comportamento que as plantas apresentaram.
A taxa assimilatória liquida diminuiu durante o ciclo de desenvolvimento da espécie em todos os tratamentos (Figura 2c). Esses resultados concordam com os observados por Valmorbida et al. (2007); Leal (2001); Payne et al. (1991); Milthorpe & Moorby (1974) e Watson (1965). Milthorpe & Moorby (1974) referem que a diminuição da taxa assimilatória liquida com a idade das plantas se deve ao sombreamento de folhas inferiores. Segundo Watson (1965), esse índice expressa o balanço entre fotossíntese e respiração, sendo, portanto, mais influenciado pelas condições climáticas, do que pelo potencial genético do vegetal. O autor ainda refere que, área foliar específica e taxa assimilatória liquida são inversamente proporcionais, pois à medida que a área foliar específica aumenta, ocorre redução na taxa assimilatória líquida, devido ao efeito do auto-sombreamento.
No presente estudo, as plantas cultivadas com 100%N/P/K/Mg apresentaram TAL com declínio mais lento ao longo do ciclo, em relação aos outros tratamentos. As plantas que apresentaram declínio mais acentuado durante todo o ciclo de desenvolvimento foram nutridas com 50%N/P/K/Mg e com 65%/N50%P/25%K/100%Mg (Figura 2c). Esse comportamento indica maior eficiência fotossintética e esse índice expressa a taxa de fotossíntese líquida, em termos de massa seca produzida (Portes & Castro Junior, 1991). Os resultados observados no presente estudo são concordantes com os registros de Boaro (2001) ao avaliar as observações de Watson (1965) e Xavier (1976). Esses autores referem à existência de evidências da influência de níveis elevados de nutrientes sobre a TAL, sugerindo a diminuição, quando baixas concentrações de nutrientes são utilizadas. Ainda referem ser pouco provável o incremento da produção pelo aumento da TAL. Ao realizarem análise de crescimento de quatro cultivares de feijão, verificaram que, em geral, a taxa assimilatória líquida era maior antes dos 20 dias após a emergência, a partir dos quais declinava com a idade da planta.
Pode-se observar que a TCR diminuiu em todos os tratamentos ao longo do ciclo de desenvolvimento da cultura (Figura 2d). Benincasa (2003) registrou que nem sempre se verificam diferenças tão evidentes entre tratamentos e, em alguns casos, não se consegue mesmo detectar causas de diferenças de produção pela análise de crescimento, o que parece ter ocorrido com a TCR no presente estudo, em especial com as plantas cultivadas com 50%N/P/K/Mg e 65 %N/50%P/25%K/100%Mg, que apresentaram curvas com declínio mais rápido indicando crescimento mais acelerado, ou seja, apresentaram as taxas mais elevadas, no início e mais baixas, no final.
Em todos os tratamentos, de acordo com as observações registradas por Boaro (2001), verifica-se que a taxa de crescimento relativo apresentou efeito maior da taxa assimilatória liquida, mostrando comportamento muito mais semelhante a ela, do que em relação a razão de área foliar. Esses resultados estão de acordo com os de Magalhães & Montojos (1971), para cultivares de feijoeiros em que a taxa assimilatória líquida e a taxa de crescimento relativo apresentaram correlação positiva. Deve ser ressaltado que, embora as comparações envolvam espécies diferentes, o comportamento dos índices fisiológicos que refletem a produtividade vegetal apresenta-se igual.
As plantas submetidas a 65%N/50%P/25%K/100%Mg apresentaram, em média maior rendimento de óleo essencial (Figura 3). Deve ainda ser destacada a tendência de produção mais precoce de óleo essencial ao longo do ciclo dessas plantas, em relação às nutridas com solução completa. Estas além, de apresentarem o ponto de máximo rendimento mais tardio não atingiram os níveis de rendimento apresentados pelas plantas nutridas com 65%N/50%P/25%K/100%Mg. Esses resultados concordam com os observados por Valmorbida et al. (2006) e David et al. (2006), de maior rendimento de óleo essencial em M. piperita cultivada em solução nutritiva com redução de 75% de K e 50% de P, respectivamente. No presente estudo, no tratamento com 65%N/50%P/25%K/100%Mg, as plantas foram cultivadas com as mesmas reduções de P e K dos estudos acima citados.
Os óleos voláteis são substâncias encontradas em várias partes das plantas (Simões & Spitzer, 1999), formados por terpenóides (mono e sesquiterpenos) e/ou lignóides (Alil-e propenilfenóis e cumarinas) (Gottileb, 1985). Os terpenos, ou terpenóides são produzidos no metabolismo secundário, ou seja, são sintetizados a partir de compostos formados no metabolismo primário. Os produtos do metabolismo secundário, embora não necessariamente essenciais para o organismo produtor, garantem vantagens para a sua sobrevivência e para a perpetuação da espécie, em seu ecossistema. A defesa contra herbívoros e microorganismos, a proteção contra raios UV, a atração de polinizadores ou animais dispersores de sementes são alguns exemplos (Wink, 1990 apud Santos, 2000; Taiz, 2004). Segundo Maia (1998), a produção e, consequentemente os teores das substâncias presentes no óleo, são muito dependentes de fatores ambientais. Por isto, plantas desenvolvidas sob diferentes condições podem conter óleos com características diferentes.
No presente estudo, as plantas nutridas com níveis de N, P e K respectivamente 35%, 50% e 75% menores em relação àqueles estabelecidos para a solução completa (T3) foram as que apresentaram maior rendimento de óleo (Figura 3). Esses resultados talvez possam ser explicados pelos níveis superestimados dos macronutrientes proporcionados pela solução nutritiva completa, conforme referem Ruiz (1998) e Zeiger (2004), excessivos no presente estudo para a menta.
A influência da nutrição mineral na produção de metabólitos secundários apresenta resultados contraditórios na literatura. Sinha & Singh (1982), ao avaliarem Mentha arvensis L. em solução nutritiva de Hoagland & Arnon modificada, verificaram que a deficiência de potássio aumentou a porcentagem do conteúdo de óleo essencial, mas diminuiu consideravelmente o total de óleo essencial por planta. Mairapetyan et al. (1999), quando estudaram a otimização das relações N:P:K em M. piperita, em cultivo hidropônico, concluíram que a menta requer maior suprimento de fósforo para o máximo acúmulo de óleo essencial. Leal (2001), trabalhando com M. piperita, em solução nutritiva nº 2 de Hoagland & Arnon (1950) com diferentes níveis de nitrogênio, concluiu que níveis de nitrogênio maiores que o recomendado na solução completa, diminuíram a produção e a qualidade do óleo essencial. Valmorbida et al. (2006) também estudando M. piperita, em solução nutritiva nº 2 de Hoagland & Arnon (1950) verificaram que o maior nível de potássio utilizado, igual a 234 mg L-1, não resultou em maior rendimento de óleo essencial. Os autores observaram ainda que para o melhor rendimento e qualidade de óleo essencial, a M. piperita deve ser cultivada com redução de 50 a 75% da quantidade de potássio proposta para a solução completa de Hoagland & Arnon (1950). Da mesma forma, David et al. (2006) concluíram que o menor nível de P, igual a 15,5 mg L-1, não foi deficiente para a M.piperita, condição em que as plantas apresentaram a maior produção de óleo essencial.
Embora a TAL e TCR não tenham mostrado diferenças marcantes entre os tratamentos, a RAF e a AFE evidenciaram o efeito da variação dos níveis dos nutrientes no crescimento vegetal. Assim, as plantas submetidas ao tratamento com o nível completo de nutrientes (100%N/P/K/Mg) exportaram assimilados com menor eficiência a partir da folha e a RMF mostrou queda mais lenta, devido à retenção desse material por mais tempo no centro de produção, as folhas. Além disso, não apresentaram melhor produtividade e as curvas da TAL e TCR mostraram quedas mais lentas. Esses níveis foram excessivos para o desempenho máximo das plantas em produtividade.
No entanto, as plantas submetidas ao tratamento com 65%N/50%P/25%K/100%Mg revelaram adequada exportação de assimilados da folha para caule e pecíolos, conforme demonstram a RMF e a distribuição de massa seca para esses órgãos. Revelaram ainda a RAF mais elevada no início do desenvolvimento e mais baixa aos 94 DAT, indicando sombreamento das folhas como resultado do crescimento. A AFE dessas plantas mostrou menor variação durante o ciclo, refletindo espessura mais constante das folhas. A TAL e a TCR apresentaram curvas decrescentes, com quedas bem acentuadas em relação às demais plantas. Todos esses índices indicam melhor produtividade das plantas submetidas ao tratamento 65%N/50%P/25%K/100%Mg. Da mesma forma, o rendimento de óleo essencial foi, em média, durante o ciclo, maior nessas plantas.
Conclui-se, portanto, que a solução completa (100%N/P/K/Mg) comprometeu a produtividade e o rendimento de óleo essencial de M. piperita L. Assim, sugere-se o cultivo da espécie com 65% de N, 50% de P, 25% de K e 100% de Mg e a colheita aos 83 dias após transplante, para obtenção de melhor rendimento de óleo essencial.
Recebido para publicação em 22/02/2008
Aceito para publicação em 13/05/2009
- ABIFITO - Associação Brasileira da Indústria Fitoterápica. Uma legislação justa para os produtos de origem natural 2002. Disponível em: <http://www.abifito.com.br>. Acesso em: 12 Jun. 2003.
- BENINCASA, M.M.P. Análise de crescimento de plantas: noções básicas. 2.ed. Jaboticabal: Fundação Nacional de Ensino e Pesquisa, 2003. 41p.
- BOARO, C.S.F. Desenvolvimento de feijoeiros (Phaseolus vulgaris) em solução nutritiva com variação dos níveis de Mg e da relação entre macronutrientes, durante o ciclo 2001. 172p. Tese (Livre Docência em Fisiologia Vegetal) - Instituto de Biociências, Universidade Estadual Paulista, Botucatu.
- BOX, M.M. Cultivo de plantas medicinalis 2.ed. Madrid: Publicaciones de Extension Agrária, 1973. 490p.
- CARMELO, Q.A.C. Saturação por bases e relações entre K, Ca e Mg no solo na nutrição potássica do milho (Zea mays L.) cv. Piranão 1992. 105p. Tese (Doutorado em Solos e Nutrição de Plantas) - Escola Superior de Agricultura de Luiz de Queiroz, Piracicaba.
- CAUSTON, D.R.; VENUS, J.C. The biometry of plant growth London: Edward Arnold, 1981. 307p.
- DAVID, E.F.S. Níveis de fósforo no desenvolvimento e produção de óleo essencial de Mentha piperita L. cultivada em solução nutritiva 2004. 159p. Dissertação (Mestrado em Agronomia/Horticultura) - Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu.
- DAVID, E.F.S.; BOARO, C.S.F.; MARQUES, M.O.M. Rendimento e composição do óleo essencial de Mentha piperita L., cultivada em solução nutritiva com diferentes níveis de fósforo. Revista Brasileira de Plantas Medicinais, v.4, n.8, p.183-8, 2006.
- DAVID, E.F.S.; BOARO, C.S.F.; MISCHAN, M.M. Desenvolvimento e rendimento de óleo essencial de menta (Mentha x piperita L.) cultivada em solução nutritiva com diferentes níveis de fósforo. Biotemas, v.20, n.2, p.5-26, 2007.
- FAGERIA, N.K.; BALIGAR, V.C.; JONES, C.A. Field Crops and mineral nutrition. In: ______. Growth and mineral nutrition of field crops 2.ed. New York: Marcel Dekker, 1997, p.1-10.
- FURLAN, M.R. Efeito da adubação com N-P2O5-K2O sobre a biomassa, o rendimento e a composição do óleo essencial de Ocimum basilicum L. cultivar Genovese 2000. 172p. Tese (Doutorado em Agronomia)- Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu.
- GOTTLIEB, O.R. Evolução e função de óleos essenciais. In: SIMPÓSIO DE ÓLEOS ESSENCIAIS, 1., 1985, São Paulo. Anais ... São Paulo: Universidade de São Paulo, 1985. p.175-91.
- HABERMANN, G. Trocas gasosas e relações hídricas em laranjeira-doce (Citrus sinensis L. Osbeck cv. Pêra) com clorose variegada dos citros (CVC) 1999. 79p. Dissertação (Mestrado em Ciências Biológicas-Botânica) - Instituto de Biociências, Universidade Estadual Paulista, Botucatu.
- HOAGLAND, D.R.; ARNON, D.I. The water: culture method for growing plants without soil. Berkeley: California Agricultural Experiment Station, 1950. 32p.
- LARCHER, W. Ecofisiologia vegetal São Carlos: RiMa, 2006. 550p.
- LEAL, F.P. Desenvolvimento, produção e composição de óleo essencial da Mentha piperita L., cultivada em solução nutritiva com diferentes níveis de nitrogênio 2001. 148p. Dissertação (Mestrado em Agronomia-Horticultura) - Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu.
- LUGG, D.G.; SINCLAIR, T.R. Seasonal changes in morfology and anatomy of field grown soybean leaves. Crop Scince, v.20, p.191-6, 1980.
- MAGALHÃES, A.C.N. Análise quantitativa de crescimento. In: FERRI, M.G. Fisiologia vegetal São Paulo: EDUSP, 1986. v.1, p.331-50.
- MAGALHÃES, A.C.; MONTOJOS, J.C. Effect of solar radiation on the growth parameters and yield of two varieties of common beans (Phaseolus vulgaris L.) Turrialba, v.21, p.165-8, 1971.
- MAIA, N.B. Efeito da nutrição mineral na qualidade do óleo essencial da menta (Mentha arvensis L.) cultivada em solução nutritiva. In: MING. L.C. et al. (Coords.). Plantas medicinais, aromáticas e condimentares: avanços na pesquisa agronômica. Botucatu: Faculdade de Ciências Agronômicas, Universidade Estadual Paulista. v.2, p.81-95. 1998
- MAIRAPETYAN, S.K. et al. Otimization of the N:P:K ratio in the nutrient medium of some soilless aromatic and medicinal plants. Acta Horticulturae, n.502, p.29-32, 1999.
- MARENCO, R.A.; GONÇALVES, J.F.C.; VIEIRA, G. Photosynthesis and leaf nutrient contents in Ochroma pyramidae (Bombacaceae). Photosynthetica, v.39, n.4, p.539-43, 2001.
- MILTHORPE, F.L.; MOORBY, J. Some aspects of overall growth and its modification. In: ______. An introduction to crop physiology London: Cambridge University Press, 1974. cap.9, p.152-79.
- MING, L.C. Influência de diferentes níveis de adubação orgânica na produção de biomassa e teor de óleo essencial de Lippia alba (Mill.), N.E.Br.-Verbenaceae 1992. 206p. Dissertação (Mestrado em Ciências - Área de Concentração Botânica) - Universidade Federal do Paraná, Curitiba.
- MORAES, M.A. Estudo de algumas alterações no crescimento do feijoeiro (Phaseolus vulgares L. cv. Carioca), causadas por diferentes níveis de boro 1986. 106p. Dissertação (Mestrado em Botânica) - Instituto de Biociências, Universidade Estadual Paulista, Botucatu.
- PAYNE, W.A. et al. Pearl millet growth as affected by phosphorus and water. Agronomic Journal, v.83, p.942-8, 1991.
- PORTES, T.A.; CASTRO JUNIOR, L.G. Análise de crescimento de plantas: Um programa computacional auxiliar. Revista Brasileira de Fisiologia Vegetal, v.3, p.53-60, 1991.
- RADFORD, P.S. Growth analysis formula; their use and abuse. Crop Science, v.7, p.171-5, 1967.
- RUIZ, H. As relações molares de macronutrientes em tecidos vegetais como base para formulação de soluções nutritivas. Ceres, v.44, n.255, p.533-46, 1997.
- SANTOS, R.I. Metabolismo básico e origem dos metabólitos secundários. In: SIMÕES. C.M.O. et al. Farmacognosia: da planta ao medicamento. 2.ed. Porto Alegre/Florianópolis: Ed. UFRGS/UFSC, 2000. p.394-412.
- SAS. The SAS System-Release 8.2. SAS Institute Inc., Cary, NC, USA. 1999-2001.
- SILVA, F.; CASALI, V.W.D. Plantas medicinais e aromáticas: pós-colheita e óleos essenciais. Viçosa: Arte Livros, 2000. 135p.
- SIMÕES, C.M.O.; SPITZER, V. Óleos voláteis. In: SIMÕES, C.M.O. et al. Farmacognosia: da planta ao medicamento. 2.ed. Porto Alegre: Universidade Federal do Rio Grande do Sul, Universidade Federal de Santa Catarina, 1999. p.394-412.
- SINHA, N.C.; SINGH, J.N. Influence of potassium deficiency on phosphorus metabolism, respiration, foliage growth and essential oil content. Plant and soil, v.66, n.1, p.283-90, 1982.
- SOARES, A.; SACRAMENTO, L.V.S. Desempenho de Mentha sp. quanto a formação de raízes adventícias em função do substrato. In: JORNADA PAULISTA DE PLANTAS MEDICINAIS, 5., 2001, Botucatu. ANAIS... Botucatu: Universidade Estadual Paulista, Botucatu.
- TAIZ, L. Metabólitos secundários e defesa vegetal. In:_______. Fisiologia vegetal 3.ed. Porto Alegre: Artmed, 2004. cap.13, p.309-32.
- VALMORBIDA, J. et al. Rendimento e composição química de óleos essenciais de Mentha piperita L. cultivada em solução nutritiva com diferentes concentrações de potássio. Revista Brasileira de Plantas Medicinais, v.8, p.56-61, 2006.
- VALMORBIDA, J. et al. Crescimento de Mentha piperita L. cultivada em solução nutritiva com diferentes doses de potássio. Revista Brasileira de Plantas Medicinais, v.9, p.27-31, 2007.
- WATSON, D.J. The physiological basis of variation in yield. Advances in Agronomy, v.5, p.343-8, 1965.
- XAVIER, F.E. Análise de crescimento de quatro cultivares de feijão (Phaseolus vulgaris L.), em duas épocas de plantio, em Viçosa, Minas Gerais 1976. 26p. Dissertação (Mestrado em Fisiologia Vegetal) - Universidade Federal de Viçosa, Viçosa.
- ZAR, J.H. Biostatistical analysis 2.ed. Englewood Cliffs: Prentice - Hall International Editions, 1986. 718p.
- ZEIGER, E. Nutrição mineral. In:_______. Fisiologia vegetal 3.ed. Porto Alegre: Artmed, 2004. cap.5, p.95-105.
Datas de Publicação
-
Publicação nesta coleção
25 Fev 2011 -
Data do Fascículo
2009
Histórico
-
Recebido
22 Fev 2008 -
Aceito
13 Maio 2009