Resumos
O experimento foi realizado com o objetivo de avaliar o efeito da idade sobre o potencial de degradação dos diferentes tecidos da lâmina foliar e do colmo de capim-braquiária (Brachiaria decumbens), capim-gordura (Melinis minutiflora) e capim-tifton 85 (Cynodon sp). Foram amostradas a 7ª (capim-braquiária e capim-gordura) e a 11ª (capim-tifton 85) lâminas foliares, no dia da exposição da lígula e 20 dias após. Por meio de observações ao microscópio foram estimadas a extensão da digestão in vitro dos tecidos da lâmina e do colmo e a redução na espessura da parede de células do esclerênquima do colmo. Lâminas foliares e segmentos de colmos jovens apresentaram maiores áreas digeridas. Permaneceram intactos os tecidos com células de parede espessada e lignificada, a bainha parenquimática dos feixes, o esclerênquima, o xilema e a epiderme do colmo. Tecidos com células de parede delgada, normalmente não-lignificada, o mesofilo, o floema e o parênquima, desapareceram completamente. O avanço na idade reduziu a digestão do mesofilo, em lâminas de capim-braquiária e capim-gordura, e do parênquima em colmos, principalmente de capim-gordura. A epiderme na lâmina foliar foi parcialmente digerida, independentemente da idade e da espécie. Embora aparentemente intactas, células esclerenquimáticas do colmo sofreram redução da espessura da parede com a incubação em líquido ruminal. A porcentagem de redução variou de 7 a 37% e a taxa de redução da espessura de 0,007 a 0,018 µm/h.
Brachiaria decumbens; Cynodon spp; espessura da parede celular; incubação em líquido de rúmen; Melinis minutiflora
A trial was carried out to evaluate the change in digestion of tropical grasses leaf and stem tissues with age. The grasses were signalgrass (Brachiaria decumbens), molassesgrass (Melinis minutiflora) and Tifton 85 bermudagrass (Cynodon sp). The 7th leaf of signalgrass and molassesgrass and the 11th leaf of bermudagrass were sampled by the time of their complete expansion (ligule exposure) and 20 days later. Segment of stem just below the sampled leaf was also sampled. Transversal sections of leaf and stem were examined at the microscope before and after rumen in vitro incubation. The degree of digestion of different leaf and stem tissues, as well as the cell wall thickness of stem sclerenchyma were estimated. Leaf blade and young stems showed larger digested areas. Stem tissues with thick and lignified cell wall, the parenchyma bundle sheath, the sclerenchyma, the xylem and the epidermis were not digested. Tissues with thin non-lignified cell wall, the mesophyll, the phloem and the parenchyma were completely digested. Molassesgrass and signalgrass leaves sampled 20 days after expansion had lower mesophyll digestion in relation to their recently expanded leaves. Similarly, molassesgrass stem parenchyma digestion decreased as plant aged. Regardless of age, leaf epidermis was only partially digested. Stem sclerenchyma cell wall showed percentage reduction in thickness from 7 to 37%, after in vitro incubation. The rate of reduction in thickness varied from 0.007 to 0.018 µm/h.
Brachiaria decumbens; Cynodon spp; cell wall thickness; ruminal incubation; Melinis minutiflora
FORRAGICULTURA
Degradação in vitro de tecidos da lâmina foliar e do colmo de gramíneas forrageiras tropicais, em função do estádio de desenvolvimento1 1 Parte da tese de Doutorado do 1 o autor apresentada à UFV.
In vitro digestion of leaf blade and stem tissues of tropical forage grasses according to stages of development
Domingos Sávio Campos PaciulloI; José Alberto GomideII; Eldo Antônio Monteiro da SilvaIII; Domingos Sávio QueirozIV; Carlos Augusto Miranda GomideV
IBolsista de Recém-Doutor do CNPq Embrapa Gado de Leite. E.mail: dscp@terra.com.br
IIPesquisador IA do CNPq. Departamento de Zootecnia - UFV. Viçosa-MG
IIIProfessor do Departamento de Biologia Vegetal - UFV. Viçosa-MG
IVPesquisador da EPAMIG - UFV. Viçosa - MG
VBolsista de Pós-doutorado da FAPESP - UNESP - Jaboticabal - SP
RESUMO
O experimento foi realizado com o objetivo de avaliar o efeito da idade sobre o potencial de degradação dos diferentes tecidos da lâmina foliar e do colmo de capim-braquiária (Brachiaria decumbens), capim-gordura (Melinis minutiflora) e capim-tifton 85 (Cynodon sp). Foram amostradas a 7ª (capim-braquiária e capim-gordura) e a 11ª (capim-tifton 85) lâminas foliares, no dia da exposição da lígula e 20 dias após. Por meio de observações ao microscópio foram estimadas a extensão da digestão in vitro dos tecidos da lâmina e do colmo e a redução na espessura da parede de células do esclerênquima do colmo. Lâminas foliares e segmentos de colmos jovens apresentaram maiores áreas digeridas. Permaneceram intactos os tecidos com células de parede espessada e lignificada, a bainha parenquimática dos feixes, o esclerênquima, o xilema e a epiderme do colmo. Tecidos com células de parede delgada, normalmente não-lignificada, o mesofilo, o floema e o parênquima, desapareceram completamente. O avanço na idade reduziu a digestão do mesofilo, em lâminas de capim-braquiária e capim-gordura, e do parênquima em colmos, principalmente de capim-gordura. A epiderme na lâmina foliar foi parcialmente digerida, independentemente da idade e da espécie. Embora aparentemente intactas, células esclerenquimáticas do colmo sofreram redução da espessura da parede com a incubação em líquido ruminal. A porcentagem de redução variou de 7 a 37% e a taxa de redução da espessura de 0,007 a 0,018 µm/h.
Palavras-chave:Brachiaria decumbens, Cynodon spp, espessura da parede celular, incubação em líquido de rúmen, Melinis minutiflora
ABSTRACT
A trial was carried out to evaluate the change in digestion of tropical grasses leaf and stem tissues with age. The grasses were signalgrass (Brachiaria decumbens), molassesgrass (Melinis minutiflora) and Tifton 85 bermudagrass (Cynodon sp). The 7th leaf of signalgrass and molassesgrass and the 11th leaf of bermudagrass were sampled by the time of their complete expansion (ligule exposure) and 20 days later. Segment of stem just below the sampled leaf was also sampled. Transversal sections of leaf and stem were examined at the microscope before and after rumen in vitro incubation. The degree of digestion of different leaf and stem tissues, as well as the cell wall thickness of stem sclerenchyma were estimated. Leaf blade and young stems showed larger digested areas. Stem tissues with thick and lignified cell wall, the parenchyma bundle sheath, the sclerenchyma, the xylem and the epidermis were not digested. Tissues with thin non-lignified cell wall, the mesophyll, the phloem and the parenchyma were completely digested. Molassesgrass and signalgrass leaves sampled 20 days after expansion had lower mesophyll digestion in relation to their recently expanded leaves. Similarly, molassesgrass stem parenchyma digestion decreased as plant aged. Regardless of age, leaf epidermis was only partially digested. Stem sclerenchyma cell wall showed percentage reduction in thickness from 7 to 37%, after in vitro incubation. The rate of reduction in thickness varied from 0.007 to 0.018 µm/h.
Key words:Brachiaria decumbens, Cynodon spp, cell wall thickness, ruminal incubation, Melinis minutiflora
Introdução
O potencial de digestão dos diferentes tecidos tem sido avaliado em seções transversais da lâmina foliar e do colmo, incubadas em líquido ruminal (Chesson et al., 1986; Wilson et al., 1991; Lempp et al., 1998). Os estudos mostram que os microorganismos colonizam praticamente todas as partículas que chegam ao rúmen. A digestão inicia-se pelas células do mesofilo e do floema (Hanna et al., 1973; Akin et al., 1973), as quais possuem apenas uma delgada parede primária não-lignificada. Estes tipos de células são facilmente fragmentadas em partículas pequenas, sendo rápida e completamente digeridas (Chesson et al., 1986).
A taxa de digestão das células do mesofilo condiciona o acesso dos microorganismos às células da bainha parenquimática dos feixes, uma vez que estas células estão envolvidas pelo mesofilo. Do ponto de vista nutricional, as células da bainha parenquimática são importantes, por apresentarem, no conteúdo celular, aproximadamente 50% da proteína foliar e alta proporção de amido. Todavia, em decorrência da elevada espessura de suas paredes, estas células apresentam, em geral, baixa taxa de digestão. Conseqüentemente, parte da proteína pode escapar da degradação ruminal naquelas células que não forem rompidas ou degradadas pelos microorganismos.
Tecidos como o esclerênquima e o xilema, formados por células de parede secundária espessada, são os que mais contribuem para a baixa qualidade da forragem (Akin, 1989; Wilson, 1993). Estes tecidos formam um sólido bloco multicelular no interior do rúmen, resultando em partículas de grande tamanho, que são pouco digeridas, em razão da lignificação e de problemas na acessibilidade dos microrganismos do rúmen à superfície da parede celular (Wilson & Mertens, 1995). Assim, os resíduos da digestão de gramíneas, independente do estádio vegetativo, contêm alta proporção de células esclerenquimáticas e xilema (Akin, 1989).
Akin (1989), compilando resultados de diferentes trabalhos sobre digestão de tecidos, sugeriu a divisão dos tecidos foliares de gramíneas C4 em rapidamente digestíveis (mesofilo e floema), lenta e parcialmente digestíveis (epiderme e células da bainha do feixe vascular) e indigestíveis (xilema e esclerênquima).
No colmo, Akin (1989) classificou a epiderme e o esclerênquima como indigestíveis, podendo-se incluir os feixes vasculares, exceto o floema. O parênquima imaturo e o floema são rápida e totalmente digestíveis, enquanto o aumento da idade reduz progressivamente a digestibilidade do parênquima.
Dos componentes químicos associados à parede celular, a lignina é o componente que, reconhecidamente, limita a digestão dos polissacarídeos da parede celular no rúmen (Jung & Deetz, 1993). Entretanto, em gramíneas, quando os microorganismos têm rápido acesso à superfície da parede celular, a digestão da parede secundária parece não ser comprometida somente pela lignificação. Wilson & Mertens (1995) sugeriram que a espessura da parede celular e o arranjo das células nos tecidos podem limitar a digestão da parede secundária, tanto quanto, ou até mais que a composição química da parede secundária.
De fato, Akin (1982) observou que a parede secundária lignificada das células esclerenquimáticas de gramíneas forrageiras apresentou considerável digestão no fluido ruminal, enquanto a lamela média e a parede primária permaneceram intactas. Wilson et al. (1991) verificaram diminuição de 54 a 85% na espessura da parede secundária das células esclerenquimáticas de três gramíneas de clima tropical, após 48 h de incubação em fluido ruminal. Outros resultados confirmaram a variável digestibilidade da parede secundária lignificada de células do esclerênquima e dos vasos de metaxilema, além da completa indigestibilidade da lamela média e da parede primária lignificadas (Wilson et al., 1993). Assim, apesar de aparentemente intactos, tecidos como o esclerênquima e o xilema podem sofrer digestão parcial, com variável redução da parede secundária.
De acordo com Wilson & Hatfield (1997), a taxa de digestão da parede de células do esclerênquima é, em média, de 0,025 µm/hora. Considerando que a espessura da parede dessas células varia de 1,30 a 3,74 mm (Moghaddam & Wilman, 1998; Wilson, 1993) e assumindo que os microorganismos têm imediato acesso à superfície da parede, conclui-se que a digestão não se completará durante o tempo de residência das partículas no rúmen. Pode-se deduzir que, quanto maior a espessura da parede celular, maior será o tempo necessário para sua completa digestão.
A importância da relativa inacessibilidade à parede secundária, resultante da estrutura física da parede celular, é reforçada pelos resultados de Grabber et al. (1992) e Wilson et al. (1993). Nesses estudos, diferentes tipos de tecidos isolados de folhas e colmos de gramíneas foram finamente moídos e incubados para digestão. Evidentemente, a maioria das características anatômicas limitantes à digestão originalmente encontradas nos tecidos foi destruída. Os resultados mostraram, surpreendentemente, elevada digestão de células esclerenquimáticas fortemente lignificadas, evidenciando que as restrições físicas exercem importante papel no processo de digestão da parede celular.
Os objetivos deste trabalho foram estimar o potencial de digestão dos diferentes tecidos e avaliar a espessura da parede de células do esclerênquima antes e após a incubação in vitro, em seções transversais da lâmina foliar e do colmo, em duas idades no perfilho das gramíneas forrageiras capim-braquiária, capim-gordura e capim-tifton 85.
Material e Métodos
As gramíneas forrageiras capim-gordura (Melinis minutiflora), capim-tifton 85 (Cynodon sp) e capim-braquiária (Brachiaria decumbens) foram cultivadas em área do Departamento de Zootecnia da Universidade Federal de Viçosa, em Viçosa, Minas Gerais. Viçosa localiza-se a 20º45' de latitude sul e 42º54' de latitude oeste, a uma altitude média de 650 m. Seu clima é do tipo Cwb (classificação Köppen), clima tropical de altitude, com verões quentes e chuvosos.
Após corte de uniformização, realizado em novembro/98 com cutelo, a altura de cinco centímetros acima do solo, foi feita adubação em cobertura com sulfato de amônio (60 kg/ha de N), cloreto de potássio (60 kg/ha de K2O) e superfosfato simples (80 kg/ha de P2O5). Durante os meses de dezembro/98 e janeiro/99 foram colhidos perfilhos para análises químicas (Paciullo et al., 2001) e anatômicas (Paciullo et al., 2002).
Em fevereiro/99 foram selecionados e identificados com anéis de mesma coloração dois grupos de 10 perfilhos em cada parcela. O critério para orientar na colheita dos perfilhos baseou-se na idade da folha. Os perfilhos foram vistoriados periodicamente para se determinar o momento da colheita. No primeiro grupo de dez perfilhos coletaram-se a 7ª (capim-braquiária e capim-gordura) e a 11ª (capim-tifton 85) folhas, da base para o topo do perfilho, no momento da completa expansão (idade 0), caracterizado pelo aparecimento da lígula. No segundo grupo de perfilhos, foram retiradas folhas dos mesmos níveis de inserção, porém 20 dias após o aparecimento da lígula (idade 20).
Foi amostrado o segmento de colmo localizado imediatamente abaixo da folha colhida. No momento da amostragem, os segmentos de colmo apresentavam seus tecidos diferenciados. Por isso, aqueles associados às folhas recém-expandidas foram chamados de colmos jovens e os associados às folhas com 20 dias de idade, de colmos em idade avançada.
No momento pré-determinado (dia do aparecimento da lígula ou 20 dias após o aparecimento da lígula), os perfilhos foram colhidos para obtenção das lâminas foliares e dos segmentos de colmo de interesse às análises de digestão dos tecidos. Após colhidos, os perfilhos foram congelados em nitrogênio líquido e, em seguida, armazenados em freezer à temperatura de -80ºC, para posteriores análises de digestão dos tecidos.
Os perfilhos retirados do freezer permaneceram por uma noite em geladeira. Após descongelamento, foram dissecados em lâmina e segmento de colmo (mesmo procedimento utilizado para análises anatômicas). Para avaliação da digestão dos tecidos, seguiu-se metodologia descrita por Akin (1982), com modificações propostas por Wilson et al. (1991). Da porção mediana da lâmina e do segmento de colmo de dois perfilhos de cada tratamento foram retirados segmentos de aproximadamente 2 cm, que, em seguida, foram seccionados transversalmente em micrótomo de mão a aproximadamente 100 mm. Três seções de cada segmento foram montadas em fita adesiva de face dupla, previamente aderida à lâmina de microscópio. Até que todas as lâminas fossem montadas com os cortes transversais, estas permaneceram em recipiente com água destilada. Em seguida, foram colocadas em tubos contendo uma mistura líquido ruminal:solução tampão de McDougall, na proporção de 1:1.
Outro grupo de três seções retiradas das mesmas frações destinadas à incubação foi também montado em lâminas de microscópio com fita adesiva de face dupla, porém não foi incubado em líquido de rúmen. Estas seções foram imediatamente observadas com auxílio do microscópio de luz e do Software de Análise de Imagens, modelo Image Pro Plus versão 1.3, para a determinação da espessura da parede das células do esclerênquima (ESC). Foram realizadas 20 medidas por seção. A técnica utilizada não permitiu obtenção de imagens de ótima qualidade para avaliação dos cortes transversais das lâminas foliares, motivo pelo qual apenas foi possível medição da espessura da parede das células do ESC do colmo. Por outro lado, a metodologia apresenta algumas vantagens como as citadas por Engels (1996): rapidez no preparo e na comparação das amostras, possibilidade de avaliar grande número de seções, antes e após a digestão em líquido de rúmen, e a manutenção do arranjo anatômico dos tecidos não-digeridos após a incubação, visto que, as seções da planta não são manuseadas após aderidas à fita.
Após 46 horas de incubação, as lâminas foram retiradas dos tubos e lavadas cuidadosamente com água destilada, sendo cobertas com lamínulas e examinadas ao microscópio. Com o uso do sistema analisador de imagens, foram medidas as espessuras da parede das células do ESC do colmo, com objetivo de se detectar possível redução na espessura, decorrente da digestão.
Para estimativa do desaparecimento dos tecidos, as imagens dos tecidos da folha e do colmo foram registradas antes e depois da incubação, copiando-as para a memória do computador e posteriormente para um disco flexível de 3,5".
Resultados e Discussão
No estudo de digestão de tecidos, foi feita análise qualitativa do desaparecimento dos tecidos da lâmina foliar e do colmo. A utilização da técnica da fita de face dupla aderida à lâmina de microscópio não permitiu obtenção de imagens de ótima qualidade para avaliação das lâminas foliares, dificultando, às vezes, a observação do desaparecimento dos tecidos. Em geral, lâminas recém-expandidas apresentaram maiores áreas digeridas. Permaneceram intactos o ESC, a bainha parenquimática dos feixes (BPF), e o xilema (XIL), enquanto o mesofilo (MES) e o floema (FLO) desapareceram totalmente, independente da espécie. A epiderme (EPI) das três espécies apresentou digestão parcial. A digestão do MES de lâminas aos 20 dias de idade de capim-braquiária e capim-gordura não foi completa. Em espécies de Panicum maximum cultivadas no período seco do ano, Lempp et al. (1998) observaram resíduos de células do MES após incubação por 72 horas. Por meio de testes histoquímicos, os autores detectaram, em algumas regiões do mesofilo, a presença de compostos fenólicos. Este fato também foi constatado por Akin (1989). Neste estudo, é provável que compostos fenólicos tenham sido depositados na parede das células do MES, com o avanço do desenvolvimento, reduzindo a taxa de digestão de lâminas com 20 dias de idade. Por outro lado, em lâminas de capim-tifton 85, a idade não alterou o desaparecimento das células do MES, que, juntamente com o FLO, foi completamente digerido. A EPI sofreu digestão parcial e o XIL, a BPF e o ESC permaneceram intactos.
Células com parede delgada e não-lignificada, como as do FLO e as do MES, geralmente apresentam completa digestão após período de incubação de 12 a 24 horas (Akin et al., 1983; Hartest et al., 1983), apesar de que, algumas vezes, a digestão não se completa após incubação por 48 horas (Twidwell et al., 1989; Wilson et al., 1991; Lempp et al., 1998). Para a BPF e a EPI, a digestão é parcial após período de incubação entre 24 e 48 horas (Akin et al., 1983; Bohn et al., 1988; Wilson et al., 1991), embora alguns autores tenham encontrado desaparecimento total das células da epiderme após incubação por 24 h (Akin & Burdick, 1975; Akin et al., 1983). Os tecidos que apresentam células com paredes espessadas e lignificadas, como o XIL e o ESC, são praticamente indigestíveis, permanecendo aparentemente intactos após longo tempo de incubação em líquido ruminal (Wilson et al., 1991; Brito et al., 1999). As variações encontradas para digestão de um mesmo tecido são decorrentes da espécie, do estádio de desenvolvimento, de fatores de ambiente e da própria metodologia utilizada para avaliação da digestão.
Segundo Lempp et al. (1997), um dos méritos da observação da seção transversal do resíduo da digestão in vitro é permitir verificar a importância da estrutura "girder" no destacamento dos diferentes tecidos e, conseqüentemente, a exposição dos mesmos aos microorganismos ruminais. Por observação visual, foi constatado, neste trabalho, que esta estrutura é totalmente resistente à digestão, mesmo em lâminas foliares recém-expandidas. De fato, espécies com menor freqüência de estrutura "girder", na lâmina foliar, apresentam maior taxa de fragmentação dos tecidos, quando comparadas àquelas com elevada freqüência (Ehlke & Casler, 1985). Este fato evidencia a importância do uso desta característica em programas de melhoramento de forrageiras, que visam a obtenção de espécies com mais alta digestibilidade (Wilson, 1997).
As células do parênquima (PAR) e do FLO de colmos jovens foram totalmente digeridas, independente da espécie. A EPI, o XIL e o ESC permaneceram aparentemente intactos (Figura 1). O desenvolvimento foi fator preponderante de alterações anatômicas e digestivas do colmo. Assim, o decréscimo na proporção de PAR, associado aos aumentos em área do ESC e da espessura da parede celular, com o avanço do desenvolvimento (Paciullo et al., 2002), resultou em menor digestão das seções de colmo, notadamente em capim-gordura. De acordo com Paciullo et al. (2002), aumento mais expressivo em área de ESC, com o desenvolvimento, e maior proporção de XIL foram observados no colmo de capim-gordura. Em colmos maduros, a EPI, o ESC, o XIL, além das células de PAR próximas ao ESC, permaneceram não-digeridos. O decréscimo na digestão do PAR, com a idade do colmo, pode ser atribuído à progressiva deposição de compostos fenólicos na parede destas células (Akin et al., 1984; Akin, 1989).
Estimativas da digestão baseadas na técnica usada neste trabalho são fundamentadas no desaparecimento de células ou tecidos. No presente estudo, a digestão foi apenas parcial, não chegando a comprometer a integridade do tecido esclerenquimático, mesmo em colmos jovens, quando ainda não havia ocorrido deposição de lignina, e a parede secundária não estava totalmente formada. Por outro lado, apesar deste tecido manter sua integridade após a incubação, observou-se redução da espessura da parede secundária, em razão de sua digestão parcial. A extensão de digestão da parede celular do esclerênquima é ilustrada na Figura 2, comparando células idênticas em seções não-incubadas e incubadas por 46 horas. A espessura da parede secundária sofreu redução, que variou de 7 a 37,6%, enquanto as taxas de digestão variaram de 0,007 a 0,018 µm/h, conforme a espécie e a idade da gramínea (Tabela 1). Outros autores relataram diminuição da espessura da parede celular após incubação em líquido ruminal, com taxas variando de 0,015 a 0,030 µm/h (Wilson et al., 1991; Akin & Hartley, 1992), dependendo da espécie, do tempo de digestão, da metodologia de avaliação, entre outros fatores.
O aumento da idade do colmo não concorreu para a redução na taxa de digestão da parede celular. Pelo contrário, as taxas foram semelhantes em capim-braquiária e capim-tifton 85 e aumentaram de 0,010 para 0,018 µm/h em capim-gordura (Tabela 1), com o avanço da idade. Este fato sugere que o aumento da lignificação na parede secundária, com o desenvolvimento do colmo, não influiu na taxa de digestão. Relatos da literatura mostram que a parede secundária sofre diferente grau de digestão (Wilson et al., 1991; Akin & Hartley, 1992), enquanto a lamela média e a parede primária lignificadas permanecem intactas após três semanas de digestão (Engels & Schurmans, 1992; Wilson et al., 1993). Este fato decorre das diferenças em composição da lignina presente nas diversas camadas da parede celular.
Assim, a lamela média e a parede primária apresentam lignina rica em unidades do tipo guaiacil, mais ramificada e estreitamente associada aos polissacarídeos e, conseqüentemente, mais resistente à digestão que a lignina rica em unidades do tipo siringil, predominantemente encontrada na parede secundária (Jung & Deetz, 1993).
Com a maior taxa de digestão (0,018 µm/h) e o tempo de incubação de 46 horas, estima-se que a máxima redução da parede celular foi de 0,83 µm, ainda assim, substancialmente inferior à menor espessura observada (1,27 µm) (Tabela 1). Portanto, mesmo na ausência da lignificação, como em colmos jovens, a digestão da parede celular não se completou durante o tempo de incubação em líquido ruminal. Este fato sugere que, em células com parede espessa (acima de 1 µm), as limitações à digestão se originam, principalmente, de problemas estruturais, conforme a hipótese de Wilson & Mertens (1995) e Wilson & Hatfield (1997) de que a baixa acessibilidade à parede secundária, resultante da sua elevada espessura, reduz a extensão da digestão da parede celular pelos microrganismos ruminais. Por essa razão, a digestão de células com parede espessada não se completa durante o tempo de permanência das partículas no rúmen, sendo os principais resíduos de gramíneas encontrados nas fezes.
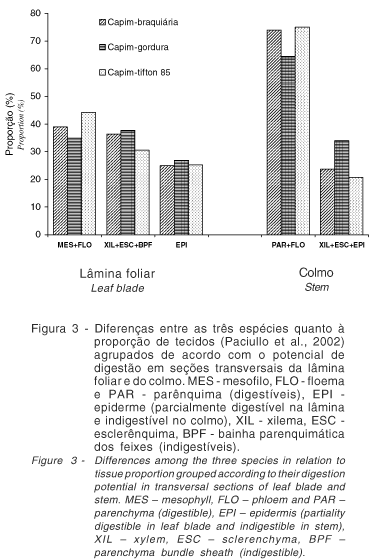
Uma análise conjunta dos dados de proporção (Paciullo et al., 2002) e de digestão de tecidos, permite inferir que, no geral, o capim-tifton 85 apresentou características anatômicas mais compatíveis com as de uma gramínea de melhor valor nutritivo (Figura 3). Isso porque, no capim-tifton 85, as mais baixas proporções de tecidos pobremente digeridos na lâmina foliar e no colmo, associadas às elevadas proporções de MES e PAR, refletiram em maiores área digeridas, mesmo em lâminas e colmos em estádio de desenvolvimento mais avançado. Por outro lado, as elevadas proporções de XIL, BPF e ESC, em capim-gordura, indicam que esta espécie apresenta características anatômicas mais típicas de forrageira de valor nutritivo mais baixo. O capim-braquiária, embora tenha apresentado proporção de tecidos intermediária às duas outras espécies, mostrou mais alta espessura da parede celular, independente do estádio de desenvolvimento. Tal característica certamente influiu negativamente na digestão das seções transversais.
Conclusões
As células do mesofilo, do parênquima e do floema foram as únicas a sofrerem completa digestão in vitro.
O avanço em idade reduziu a digestão do mesofilo, em capim-braquiária e capim-gordura, e do parênquima, nas três espécies.
Apesar da manutenção da integridade do esclerênquima do colmo, as células deste tecido apresentaram variada redução na espessura da parede celular, o que evidencia sua parcial digestão.
Literatura citada
Recebido em: 25/05/01
Aceito em: 11/12/01
- AKIN, D.E. Section to slide technique for study of forage anatomy and digestion. Crop Science, v.22, p.444-446, 1982.
- AKIN, D.E. Histological and physical factors affecting digestibility of forages. Agronomy of Journal, v.81, n.1, p.17-25, 1989.
- AKIN, D.E.; BURDICK, D. Percentage of tissue types in tropical and temperate grass leaf blades and degradation of tissues by rumen microorganisms. Crop Science, v.15, n.5, p.661-668, 1975.
- AKIN, D.E.; HARTLEY, R.D. UV absorption microspectrophotometry and digestibility of cell types of bermudagrass internodes at different stages of maturity. Journal of Science Food Agriculture, v.59, p.437-447, 1992.
- AKIN, D.E.; BROWN, R.H.; RIGSBY, L.L. Digestion of stem tissues in Panicum species. Crop Science, v.24, n.4, p.769-773, 1984.
- AKIN, D.E.; WILSON, J.R.; WINDHAM, W.R. Site and rate of tissue digestion in leaves of C3, C4, and C3/C4 intermediate Panicum species. Crop Science, v.23, n.1, p.147-155, 1983.
- AKIN, D.E.; AMOS, H.E.; BARTON, F.E. et al. Rumen microbial degradation of grass tissue by scanning electron microscopy. Agronomy of Journal, v.65, n.5, p.825-888, 1973.
- BOHN, P.J.; BROWN, R.H.; AKIN, D.E. "In vitro" dry matter digestibility, leaf anatomy, and fiber concentration of a hybrid between C3 and C3-C4 Panicum species. Crop Science, v,28, p.332-336, 1988.
- BRITO, C.J.F.A.; RODELLA, R.A.; DESCHAMPS, F.C. et al. Anatomia quantitativa e degradação "in vitro" de tecidos em cultivares de capim-elefante (Pennisetum purpureum Schumach). Revista Brasileira de Zootecnia, v.28, n.2, p.223-229, 1999.
- CHESSON, A.; STEWART, C.S.; DALGARNO, K. et al. Degradation of isolated grass mesophyll, epidermis and fibre cell wall in the rumen and by cellulolitic rumen bacteria in axemic culture. Journal Applied Bacteriology, v.60, n.4, p.327-336, 1986.
- EHLKE, N.J.; CASLER, M.D. Anatomical characteristics of smooth bromegrass clones selected for in vitro dry matter digestibility. Crop Science, v.25, p.513-517, 1985.
- ENGELS, F.M. Developments in application of light and scanning electron microscopy techniques for cell wall degradation studies. Netherlands Journal of Agriculture Science, v.44, p.357-373, 1996.
- ENGELS, F.M.; SCHUURMANS, J.L.L. Relationship between structural development of cell walls and degradation of tissues in maize stems. Journal of Science Food Agriculture, v.59, p.45-51, 1992.
- GRABBER, J.H.; JUNG, G.A.; ABRAMS, S.M. et al. Digestion kinetics of parenchyma and sclerenchyma cell walls isolated from orchardgrass and switchgrass. Crop Science, v.32, p.806-810, 1992.
- HANNA, W.W.; MONSON, W.G.; BURTON, G.W. Histological examination of fresh forages leaves after "in vitro" digestion. Crop Science, v.13, n.1, p.98-102, 1973.
- HARTEST, A.A.; OWENSBY, C.E.; HARBERS, L.H. Rumen microbial degradation of indiangrass and big bluestem leaf blades. Journal of Animal Science, v.57, n.6, p.1626-1636, 1983.
- JUNG, H.G.; DEETZ, D.A. Cell wall lignification and degradability. In: JUNG, H.G. et al. (Eds.) Forage cell wall structure and digestibility Madison: American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, 1993. p.315-346.
- LEMPP, B.; EZEQUIEL, J.M.B.; SANTOS, J.M. et al. Observação da estrutura girder na taxa de digestão dos tecidos em lâminas de Panicum maximum Jacq. cv. aruana e vencedor. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 34., 1997, Juiz de Fora. Anais... Juiz de Fora: Sociedade Brasileira de Zootecnia, 1997. p.12-14.
- LEMPP, B.; EZEQUIEL, J.M.B.; SANTOS, J.M. et al. Observação da taxa de digestão das células de mesofilo de duas cultivares de Panicum maximum Jacq., águas e seca. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998. p.269-71.
- MOGHADDAM, P.R.; WILMAN, D. Cell wall thickness and cell dimensions in plant parts of eight forage species. Journal of Agriculture Science, v.131, p.59-67, 1998.
- PACIULLO, D.S.C.; GOMIDE, J.A.; QUEIROZ, D.S. et al. Composição química e digestibilidade "in vitro" de lâminas foliares e colmos de gramíneas forrageiras, em função do nível de inserção no perfilho, da idade e da estação de crescimento. Revista Brasileira de Zootecnia, v.30, n.3, p.964-974, 2001 (Suplemento 1).
- PACIULLO, D.S.C.; GOMIDE, J.A.; SILVA, E.A.M. et al. Características anatômicas da lâmina foliar e do colmo de gramíneas forrageiras tropicais, em função do nível de inserção no perfilho, da idade e da estação de crescimento. Revista Brasileira de Zootecnia, v.31, n.2, p.890-899, 2002. (suplemento)
- TWIDWELL, E.K.; JOHNSON, K.D.; BRACKER, C.E. et al. Plant tissue degradation measurement using image analysis. Agronomy of Journal, v.81, n.5, p.837-840, 1989.
- WILSON, J.R. Organization of forage plant tissues. In: JUNG, H.G.; BUXTON, D.R.; HATFIELD, R.D. et al. (Eds.) Forage cell wall structure and digestibility Madison: American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, 1993. p.1-32.
- WILSON, J.R.; HATFIELD, R.D. Structural and chemical changes of cell wall types during stem development: consequences for fibre degradation by rumen microflora. Australian Journal of Agriculture Research, v.48, p.165-180, 1997.
- WILSON, J.R.; MERTENS, D.R. Cell wall accessibility and cell structure limitations to microbial digestion of forage. Crop Science, v.35, n.1, p.251-259, 1995.
- WILSON, J.R.; DEINUM, B.; ENGELS, F.M. Temperature effects on anatomy and digestibility of leaf and stem of tropical and temperate forage species. Netherlands Journal of Agriculture Science, v.39, n.1, p.31-48, 1991.
- WILSON, J.R.; MERTENS, D.R.; HATFIELD, R.D. Isolates of cell types from sorghum stems: digestion, cell wall and anatomical characteristics. Journal of Science Food Agriculture, v.63, p.407-417, 1993.
Datas de Publicação
-
Publicação nesta coleção
08 Set 2004 -
Data do Fascículo
Abr 2002
Histórico
-
Aceito
11 Dez 2001 -
Recebido
25 Maio 2001