Resumos
O estudo foi conduzido com o objetivo de identificar a composição botânica da dieta de bovinos criados extensivamente no Pantanal através da técnica microhistológica fecal, numa área característica da sub-região da Nhecolândia, Pantanal, no período de outubro/97 a setembro/99, e verificar a variação existente em função do mês, época, ano e estado fisiológico das vacas. Foram confeccionadas 726 lâminas, sendo cada lâmina uma unidade observacional. Com base na distribuição dos valores máximos de ocorrência da composição botânica de cada espécie/lâmina e nos quantis, as espécies foram classificadas em quatro grupos principais: 1- principal (máx >20%); 2- médio superior (12,3% <máx<=20,0%); 3- médio (6,7%<máx<=12,3%); 4-ocasional (máx<+6,7%). Estes grupos representaram 11; 14; 25 e 50% das espécies identificadas, respectivamente. Um delineamento de parcelas subdivididas no tempo distinto foi ajustado envolvendo cada efeito e suas interações com estado fisiológico e ano hidrológico. Para todos os grupos estudados não houve efeitos significativos de estado fisiológico das vacas e da interação estado fisiológico com época e ano sobre a composição média da dieta. Houve efeitos significativos de época e ano para todos os grupos de espécies, porém, a interação época x ano foi significativa somente para o grupo ocasional.
microhistologia fecal; pastagem nativa; gado de corte
This study aimed to identify the botanical composition of the diet of cattle extensively raised on native grasslands in the Pantanal wetland through the fecal microhystological technique, on a characteristic area of the subregion of Nhecolândia, from October/97 to September/99, and to verify the variation as affected by month, season, year and physiological condition of cows. A set of 726 microhystological slides was prepared, each slide being an observational unit. Based on distribution of maximum occurrence values of each species/slide and in the quantis, the species were classified into four main groups: 1- principal (max >20.0%); 2- medium superior (12.3% <max<=20.0%); 3- medium (6.7<max<=12.3%); 4-occasional (max<+6.7%). These groups represented 11%, 14%, 25% and 50% of the identified species, respectively. A split-plot in distinct time was adjusted envolving each effect (month, season) and its interactions with physiological condition and hydrological year. For all studied groups there were no significant effects of physiological condition of cows and no interaction of physiological condition with period and year on the mean composition of the diet. There were significant effects of season and year for all groups, however, the interaction season x year was significant only for the occasional group.
fecal microhystology; native grassland; beef cattle
Composição Botânica da Dieta de Bovinos em Pastagem Nativa na Sub-Região da Nhecolândia, Pantanal1 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT.
Sandra Aparecida Santos2 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT. , Ciniro Costa3 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT. , Geraldo da Silva e Souza4 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT. , Arnildo Pott5 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT. , Juliana Magalhães Alvarez6 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT. , Silvia Rodrigues Machado7 1 Parte da tese de Doutorado apresentada à FMVZ-UNESP, Campus de Botucatu, SP. Parcialmente financiada pelo FUNDECT.
RESUMO - O estudo foi conduzido com o objetivo de identificar a composição botânica da dieta de bovinos criados extensivamente no Pantanal através da técnica microhistológica fecal, numa área característica da sub-região da Nhecolândia, Pantanal, no período de outubro/97 a setembro/99, e verificar a variação existente em função do mês, época, ano e estado fisiológico das vacas. Foram confeccionadas 726 lâminas, sendo cada lâmina uma unidade observacional. Com base na distribuição dos valores máximos de ocorrência da composição botânica de cada espécie/lâmina e nos quantis, as espécies foram classificadas em quatro grupos principais: 1- principal (máx >20%); 2- médio superior (12,3% <máx<=20,0%); 3- médio (6,7%<máx<=12,3%); 4-ocasional (máx<+6,7%). Estes grupos representaram 11; 14; 25 e 50% das espécies identificadas, respectivamente. Um delineamento de parcelas subdivididas no tempo distinto foi ajustado envolvendo cada efeito e suas interações com estado fisiológico e ano hidrológico. Para todos os grupos estudados não houve efeitos significativos de estado fisiológico das vacas e da interação estado fisiológico com época e ano sobre a composição média da dieta. Houve efeitos significativos de época e ano para todos os grupos de espécies, porém, a interação época x ano foi significativa somente para o grupo ocasional.
Palavras-chave: microhistologia fecal, pastagem nativa, gado de corte
Botanical Composition of Cattle Diet on Rangelands in the Pantanal Wetland, Brazil
ABSTRACT - This study aimed to identify the botanical composition of the diet of cattle extensively raised on native grasslands in the Pantanal wetland through the fecal microhystological technique, on a characteristic area of the subregion of Nhecolândia, from October/97 to September/99, and to verify the variation as affected by month, season, year and physiological condition of cows. A set of 726 microhystological slides was prepared, each slide being an observational unit. Based on distribution of maximum occurrence values of each species/slide and in the quantis, the species were classified into four main groups: 1- principal (max >20.0%); 2- medium superior (12.3% <max<=20.0%); 3- medium (6.7<max<=12.3%); 4-occasional (max<+6.7%). These groups represented 11%, 14%, 25% and 50% of the identified species, respectively. A split-plot in distinct time was adjusted envolving each effect (month, season) and its interactions with physiological condition and hydrological year. For all studied groups there were no significant effects of physiological condition of cows and no interaction of physiological condition with period and year on the mean composition of the diet. There were significant effects of season and year for all groups, however, the interaction season x year was significant only for the occasional group.
Key Words: fecal microhystology, native grassland, beef cattle
Introdução
O Pantanal Mato-Grossense possui extensas áreas de campos naturais, favorecendo a atividade pastoril, razão pela qual, a região tem sua economia voltada principalmente para a exploração extensiva da pecuária de corte. A composição florística é variável em função dos gradientes hidrotopográficos (nível de inundação), tipo de solos e fatores antrópicos (intensidade de uso e manejo). Esta grande diversidade de ambientes e espécies forrageiras favorece a pecuária, por permitir maior seletividade de pastejo aos bovinos (Pott, 1997). Na última listagem efetuada por Pott & Pott (1999), a flora do Pantanal apresentou 1863 espécies, pertencentes a 774 gêneros e 136 famílias. As formações vegetacionais básicas são as arbóreas, as gramíneo-lenhosas (savanas), as campestres e as aquáticas, distribuídas em mosaico, nas diferentes unidades de paisagem da região.
Apesar dos recursos forrageiros constituírem a principal fonte de alimentos para os grandes herbívoros silvestres e domésticos do Pantanal, pouco se sabe como esses recursos são usados ao longo dos meses e dos anos. Como nas demais áreas de pastagens nativas, o principal desafio enfrentado pelos técnicos refere-se ao manejo destas áreas heterogêneas. Atualmente, tem-se a consciência de que somente através do conhecimento mais aprofundado da relação animal x planta e dos processos biológicos importantes, tais como a avaliação da composição botânica da dieta, será possível manejar adequadamente as pastagens nativas (Provenza, 1991), bem como avaliar o impacto dos herbívoros sobre a vegetação.
Existem várias técnicas e procedimentos usados para a avaliação da composição botânica da dieta de herbívoros, todos com vantagens e desvantagens (Holechek et al., 1982). Como a maior parte das metodologias apresenta fontes de erros inerentes (Duarte et al., 1992; Henley et al., 2001), a escolha depende do ecossistema e da espécie animal a ser estudada. A análise fecal é uma técnica amplamente usada nos estudos de dieta de grandes herbívoros criados extensivamente, pois abrange uma escala espacial e temporal mais ampla da dieta do que as outras técnicas (Norbury & Sanson, 1992). Apesar de a análise microhistológica fecal apresentar limitações na identificação de determinadas espécies forrageiras, decorrentes da digestão e discernibilidade diferenciada, McInnis & Vavra (1987) consideraram-na como a única metodologia prática para determinar a dieta de grandes herbívoros criados extensivamente.
Desta forma, o estudo foi conduzido com o objetivo de identificar a composição botânica mensal da dieta de vacas de cria através da técnica de microhistologia fecal durante dois anos hidrológicos, numa área característica da sub-região da Nhecolândia, Pantanal, MS.
Material e Métodos
Área de estudo
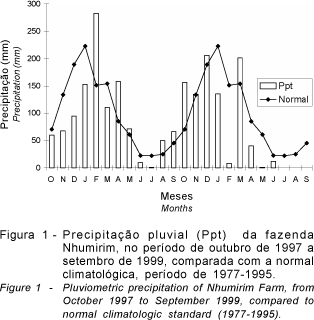
O estudo foi conduzido na fazenda Nhumirim, sub-região da Nhecolândia, Pantanal de Mato Grosso do Sul, numa área de pastagem nativa com 151 hectares (lat. 19°04'S, long. 56 36'W, alt. 98m), no período de outubro/97 a setembro/99, englobando dois anos hidrológicos. O clima da região é do tipo Aw de Koeppen: clima tropical, megatérmico, com inverno seco e chuvas no verão. Na Figura 1 é possível observar que a precipitação pluvial de 1997/98 esteve próxima da normal climatológica, o que não ocorreu no ano de 1998/99. Apesar da precipitação média acumulada de 1998/99 ser menor, ocorreu inundação parcial da área no mês de janeiro e esta não foi de longa duração devido ao baixo nível de precipitação ocorrido no mês de fevereiro (abaixo do normal). Considerou-se períodos trimestrais para a avaliação da dieta: outubro a dezembro, que corresponde ao início das chuvas; janeiro a março, que corresponde ao pico das chuvas; abril a junho, período de provável inundação da área, dependente da intensidade das chuvas dos meses anteriores; julho a setembro, que corresponde ao período de seca. Para tornar a análise dos resultados prática, o ano hidrológico 1997/98 foi considerado 'normal' e o ano hidrológico 1998/99 'atípico', baseado na normal climatológica da área (Soriano, 1999).
Os solos da área de estudo foram classificados como Areia Quartzosa nas unidades de paisagem localizadas nas cotas mais elevadas do mesorelevo (floresta semidecídua, cerradão e campo-cerrado), enquanto que nas demais unidades de paisagem, os solos foram classificados como Areia Quartzosa Hidromórfica e Podzol Hidromórfico (Carvalho Filho et al., 2000).
A área de estudo é denominada de 'Pantanal alto', pois corresponde às áreas que sofrem inundação principalmente de origem pluvial, embora parte da área estudada seja parcialmente influenciada pela água de origem fluvial, decorrente do transbordamento dos rios. No Pantanal alto, a inundação normalmente atinge somente as áreas mais baixas do mesorelevo, o que possibilita a permanência do rebanho durante o ano todo. A área de estudo apresenta unidades de paisagem (fitofisionomias) características da sub-região da Nhecolândia. Estas foram classificadas com base nos estudos de Comastri-Filho (1984) e Pott et al. (1997) com as seguintes denominações: 1) floresta semidecídua, regionalmente conhecida como mata. Na parte interna a vegetação é esparsa, com predominância de acuri (Scheelea phalerata), ambiente geralmente usado para abrigo e refúgio dos bovinos, enquanto que as bordas apresentam muitas espécies como diversas trepadeiras, destacando-se 'cipó-de-arraia-liso' (Cissus erosa), 'cipó-dágua' (Arrabidaea sp), 'cipó-prata' (Mascagnia benthamiana (Gris.) Anderson) e 'buta' (Cissampelus pareira L.). No extrato inferior predomina o 'gravateiro' (Bromelia balansae Mez); 2) savana florestada, regionalmente conhecida como cerradão, que se caracteriza pela presença de espécies arbóreas como 'jatobá' (Hymenaea courbaril), 'coroa-de-frade' (Mouriri elliptica Mart.), 'gonçalo' (Astronium fraxinifolium Schott) e trepadeiras como 'japecanga' (Smilax fluminensis); 3) savana arborizada, regionalmente conhecida como campo cerrado, que compreende uma área de transição entre campo limpo e cerradão, com presença de espécies herbáceas (Elyonurus muticus, Axonopus purpusii, Mesosetum chaseae - grama-do-cerrado) e espécies de cerrado (Curatella americana - lixeira, Byrsonima orbignyana - canjiqueira, Annona dioica - ariticum); 4) savana gramíneo-lenhosa, regionalmente conhecida como campo limpo, com predominância de gramíneas como o capim mimoso (Axonopus purpusii), capim rabo-de-burro (Andropogon bicornis) e capim vermelho (Andropogon hypogynus); 5) savana gramíneo-lenhosa, regionalmente conhecida como caronal, com predominância do capim carona (Elyonurus muticus); 6) lagoas permanentes, regionalmente conhecidas como baías, em cujas bordas destacam o lodo (Eleocharis minima), Cyperus brevifolius, Pontederia parviflora e grama-do-carandazal (Panicum laxum); 7) lagoas temporárias que podem secar totalmente no período seco. As principais espécies encontradas são P. parviflora, Diodia kuntzei, capim-de-capivara (Hymenachne amplexicaulis), capim mimosinho (Reimarochloa brasiliensis) e Panicum laxum; 8) vazantes e baixadas. Destas unidades de paisagem, apenas as matas, cerradão, campo-cerrado e caronal geralmente não sofrem inundação.
Acompanhando o esquema de manejo da fazenda, realizou-se queima controlada de alguns locais com 'macegas' ('patches' com gramíneas cespitosas maduras), geralmente presentes nas áreas de campo limpo e caronal. Estas queimas localizadas foram efetuadas em locais diferentes nos meses de outubro, abril, maio e julho do ano de 1997/98 e em outubro e novembro do ano de 1998/99.
Identificação da dieta
Os animais utilizados formaram um rebanho de 46 vacas de cria Nelore, do qual foram selecionadas para observação dentro do rebanho, vacas em função do seu estágio fisiológico: dez vacas com bezerro ao pé, cinco vacas vazias, cinco vacas prenhes e cinco novilhas. Procurou-se manter ao máximo o rebanho já existente na área, sendo introduzidas apenas cinco novilhas. Os animais foram mantidos em pastejo contínuo, numa lotação de leve a média. O rebanho foi levado ao curral mensalmente, onde os animais selecionados foram identificados com tinta colorida em ambos os lados do costado, visando facilitar a observação no campo e verificar se as vacas ainda estavam no mesmo estágio fisiológico, bem como renovar a identificação.
Durante o período de estudo, montou-se uma coleção de referência das principais espécies forrageiras presentes na área, um pré-requisito para análise fecal. Das 286 espécies forrageiras catalogadas na área, foi colhido material fresco de 120 plantas, principalmente folhas. A escolha das espécies usadas para compor a coleção baseou-se na abundância e naquelas que se constituem em potencial de alimento para os bovinos (observados diretamente no decorrer do estudo). O material foi estocado em álcool 70°, do qual foram preparadas duas lâminas para cada espécie forrageira. Para o preparo das lâminas utilizou-se o método de Scott & Dahl (1980) com algumas adaptações. As amostras de cada espécie foram cortadas com o auxílio de uma tesoura, lavadas e batidas no liquidificador com a adição de água até a completa fragmentação. O material foi coado em peneira tipo ABNT 140 e lavado com água corrente. Uma pequena alíquota deste material foi colocada no centro de uma lâmina histológica, cujo material foi uniformemente espalhado, e em seguida, adicionada a solução clareadora de Hertwig. Após o aquecimento e revolvimento do material, foi adiconada a solução de montagem de Hoyer. A composição de ambas as soluções foi descrita por Middeleton & Rojas (1994).
As estruturas epidérmicas de cada espécie forrageira foram analisadas com o auxílio de um microscópio de luz, desenhadas e fotografadas, com base nos seguintes caracteres de identificação ou diagnóstico: forma das células curtas (corpos de sílica) no caso das gramíneas, forma das células longas, tipos de tricomas/drusas/espículas, tipos de estômatos, forma das células subsidiárias dos estômatos (Metcalf, 1960; Metcalf & Chalk, 1979).
Com base nos caracteres da epiderme foliar, foi elaborado um guia de identificação, cujas principais espécies estão descritas, a seguir:
Família GRAMINEAE (POACEAE)
Células da epiderme linearmente arranjadas, paralelas às nervuras e ao eixo da folha; estômatos em fileira na zona intercostal; células guardas em forma de osso.
Andropogon hypogynus Hack. (capim-vermelho): corpos de sílica na zona costal na forma de haltere ou algo nodular, dispostos em longas fileiras. Células de sílica na zona intercostal principalmente na forma crenada e isoladas, mas às vezes aparecem aos pares na forma crenada e de cruz. Células subsidiárias arredondadas, na forma de cúpula ou triangular. Células longas com paredes finas e levemente sinuosas. Sobre a superfície adaxial, presença de micropelos (tipo panicóide) entre as células longas e algumas espículas entre as células de sílica sobre as nervuras.
Axonopus purpusii (Mez) Chase (mimoso): corpos de sílica na zona costal na forma de cruz e haltere ou alternados. Células de sílica na zona intercostal em pares na forma de meia lua e crenada ou isoladas. Células longas da zona intercostal com ondulações regulares. Células subsidiárias dos estômatos na forma triangular. Presença de micropelos entre as células longas, cuja parte distal é pouco visível. Macropelos retos e unicelulares. Presença de muitas espículas entre as células longas na superfície adaxial da folha.
Hymenachne amplexicaulis (Rudge) Nees (capim-de-capivara): corpos de sílica na zona costal na forma de cruz ou haltere, dispostos em longas fileiras. Células de sílica na zona intercostal principalmente pareadas, na forma de cruz ou tipo oriza (forma de oito) e crenadas, e isoladas na forma crenada. Células subsidiárias levemente triangulares. Células longas da zona intercostal alongadas com paredes finas e irregularmente sinuosas. Alguns macropelos unicelulares. Sobre a supefície adaxial, presença de espículas.
Leersia hexandra- Presença de câmaras de ar ou espaço livre entre as células de sílica da zona costal. Corpos de sílica apresentam a forma de oito (tipo oriza). Na zona intercostal, as células de sílica apresentam-se isoladas ou em pares, de formato variado. Células subsidiárias dos estômatos na forma triangular ou arredondadas. Células longas sinuosas e presença de macropelos unicelulares. Presença de espículas grandes, de bases infladas e presença de pequenas papilas de formato variável, presente entre as células longas.
Mesosetum chaseae Luces (grama-do-cerrado): corpos de sílica na zona costal e intercostal principalmente pareados, na forma de cruz e crenada. Células subsidiárias dos estômatos na forma de cúpula. Células longas da zona intercostal com paredes sinuosas. Micropelos (de difícil visualização) entre as células longas, com bases arredondadas e rodeado por uma célula epidérmica. Sobre a superfície adaxial, presença de espículas entre as células longas.
Panicum laxum Sw. (grama-do-carandazal): células de sílica na zona costal na forma intermediária entre cruz e haltere e ocasionalmente na forma de cruz, dispostas em longas fileiras. Presença frequente de espículas sobre as nervuras. Na zona intercostal, poucas células de sílica e quando presentes na forma de cruz. Células subsidiárias dos estômatos são triangulares a ovais. Células longas da zona intercostal são alongadas, estreitas e com ondulações regulares. Micropelos com uma célula basal curta e uma célula distal mais longa com as extremidades afuniladas(tipo panicóide). Presença de alguns macropelos longos e rígidos.
Família CYPERACEAE
Eleocharis minima Kunth (lodo): corpos de sílica de formato oval sobre a superfície da epiderme. Células longas estreitas e alongadas e células subsidiárias dos estômatos alongadas. Geralmente, observa-se uma fileira de estômatos na zona intercostal. Eleocharis acutangula (Roxb.) Steud. (três-quina): corpos de sílica de formato oval sobre a superfície epidérmica. Células subsidiárias dos estômatos alongadas. Células longas e estreitas, com paredes levemente sinuosas. Geralmente, observam-se duas ou mais fileiras de estômatos na zona intercostal. Família PALMAE (ARECACEAE)
Sheelea phalerata (Mart.) Bur. (acuri): diversos corpos de sílica de formato oval dispostos em fileiras sobre a superfície epidérmica. Na zona intercostal, células longas com formatos diversos e irregulares, com paredes finas e lisas. Estômatos tetracíclicos, rodeados por quatro células subsidiárias, duas paralelas às células guarda e duas polares, geralmente menores.
A composição botânica da dieta foi determinada pela técnica microshistológica desenvolvida por Sparks & Malecheck (1968) e modificada por Scott & Dahl (1980). Os animais identificados foram observados mensalmente, durante três dias consecutivos, do nascer ao pôr-do-sol. Assim que os animais defecavam eram anotados o local e o número do animal, e em seguida as amostras eram colhidas diretamente do chão, procurando não influenciar o comportamento animal. Procurou-se colher amostras de todas as categorias, de preferência de diferentes animais e dias, num mínimo de 15 animais (amostras)/mês. As amostras foram acondicionadas em vidros com álcool 70°. No processo de montagem das lâminas foram utilizadas a solução de Hoyer e a de Hertwig, técnica similar à usada na montagem das lâminas de referência. Para cada amostra (indivíduo) coletada foram confeccionadas duas lâminas e em cada lâmina foram selecionados sistematicamente 20 campos de leitura e observados num microscópio de luz com aumento de 100 vezes. Ao todo, foram confeccionadas 726 lâminas, nas quais foram observados um total de 14.520 campos.
A leitura foi efetuada por apenas um observador, após um treinamento intenso através de lâminas confeccionadas manualmente. A identificação dos fragmentos das plantas visualizados em cada campo das lâminas das fezes foi feito com base principalmente, em certas características de diagnóstico da epiderme das folhas, comparadas com materiais de referência e um guia de identificação previamente elaborado. Como em cada campo de leitura, nem todos os fragmentos eram de origem epidérmica, muitos não foram identificados. Para evitar a superestimativa de algumas espécies, tricomas e drusas foram considerados, quando vistos pela primeira vez em cada lâmina, porém, para sua posterior identificação nos outros campos, só foram considerados quando unidos aos tecidos da periderme.
Com base em estudos efetuados por Holecheck & Gross (1982a) e Santos et al. (2001a), optou-se pela estimativa da composição da dieta em termos de peso seco com o uso da freqüência de ocorrência, obtida pela relação entre a freqüência de cada espécie (componente) e a somatória das freqüências. Os valores obtidos não foram corrigidos, pois Santos et al. (2001a) testaram o uso de fatores de correção em dietas preparadas manualmente através da equação de Norbury (1988) e verificaram que, apesar destes melhorarem a estimativa da dieta, o seu uso não foi considerado justificável.
Na análise estatística, cada lâmina foi considerada como uma unidade observacional. Através do PROC MEANS (SAS), obteve-se o máximo de ocorrência da composição de cada espécie/lâmina. Posteriormente, aplicou-se o PROC UNIVARIATE (SAS) sobre a distribuição dos valores máximos obtidos. A composição percentual da dieta associada às variáveis do grupo principal foi analisada como uma parcela subdividida não balanceada no tempo, com o uso do PROC MIXED (SAS, 1999). Os efeitos principais das parcelas foram dias, meses, períodos (época), categoria e respectivas interações. Nas subparcelas foram analisados os efeitos do ano hidrológico e da interação de cada um dos fatores com o ano hidrológico. Como o ensaio não é equilibrado, para evitar confundimento de efeitos, as classificações de dia, mês e época (períodos trimestrais) foram analisadas separadamente. Um delineamento de parcelas subdivididas (no tempo) distinto foi ajustado envolvendo cada efeito e suas interações com categoria e ano hidrológico. A melhor análise resultou do modelo com época, seguido do modelo com meses e por último o modelo com dias. Na análise de variância da participação percentual na dieta do grupo principal, a variável resposta foi considerada de três formas: não transformada, com transformação logarítmica e com uso de postos (não paramétrica). A transformação log produziu o maior valor da função log-verossimilhança. Independentemente do modelo utilizado, do ponto de vista de significância dos efeitos, os resultados foram essencialmente os mesmos o que serve ao propósito de validação da ANOVA não transformada, que é de mais fácil interpretação.
A avaliação de efeitos principais e interações foi levada a efeito com o uso do modelo linear:

Em que i representa uma categoria de animal, j um nível do fator de interesse (dia, mês ou época), l um ano, k uma das repetições aninhadas na interação de 2a ordem dos fatores presentes nas comparações de parcelas e v, uma das repetições aninhadas na interação de 3a ordem presente nas comparações envolvendo as subparcelas. Os efeitos dos fatores representados por a, r e d, bem como as respectivas interações (ar), (ad) , (rd), e (ard), são fixos. Os componentes q e e são estocásticos e independentemente distribuídos e correspondem aos erros associados às parcelas e subparcelas, respectivamente. A análise levada a efeito com o PROC MIXED usa o método da verossimilhança restrita no processo de estimação e assume distribuição normal para as duas componentes de erro.
As respostas da composição botânica das espécies do grupo principal para os fatores época e ano foram avaliadas com o teste de Wald (SAS, 1999). A abordagem se justifica face ao elevado número de repetições disponíveis no ensaio.
Resultados e Discussão
Bovinos mantidos em pastagens nativas encontram uma ampla diversidade de plantas de aceitabilidade variável. Na área de estudo foram catalogadas 286 espécies de plantas, pertencentes a 77 famílias. A família com maior número de espécies foi a Gramineae com 47 espécies, seguida de Leguminosae com 45 espécies e Cyperaceae, com 20 espécies. Na leitura das lâminas fecais foram identificadas 83 espécies, compreendendo cerca de 30% do total das espécies da área. Com base nos quantis dos valores máximos obtidos, as espécies foram classificadas em quatro grupos: 1- principal (máx >20%); 2- médio superior (12,3% < máx <= 20,0%); 3- médio (6,7 < máx <= 12,3%); 4- ocasional (máx. <=6,7%). Das 83 espécies identificadas, apenas 11% (9 espécies dentro de 10 componentes) foram incluídas no grupo principal. O grupo médio superior englobou 14%, o grupo médio, 25% e o grupo ocasional, 50% das plantas. Na Tabela 1 são listadas as forrageiras identificadas nas lâminas de microhistologia fecal, com os respectivos valores mínimo e máximo, médias e erros padrão, hábito de crescimento, forma biológica de Raunkiaer, unidade de paisagem nas quais as espécies foram encontradas e o grau de consumo.
Levando em consideração a imensa diversidade florística existente no Pantanal da Nhecolândia, poucas espécies tiveram participação expressiva na dieta dos bovinos. Como nas demais áreas marginais, a produtividade animal no Pantanal é influenciada pela estacionalidade de produção das pastagens, variável em nível espacial e temporal. Na região, a composição botânica da pastagem é extremamente dependente do regime hídrico e da água freática, pois os solos são de composição granulométrica grosseira com capacidade baixa de retenção de água (Cunha, 1980).
A composição botânica da dieta de bovinos englobou principalmente as espécies herbáceas pertencentes às famílias Gramineae (45,0%) e Cyperaceae (12,0%), que corresponderam aproximadamente a 76,5 e 13% do peso seco da dieta, respectivamente. Estudos realizados em outros locais têm comprovado que as gramíneas constituem o principal componente da dieta de bovinos (Bóo et al., 1993; Guevara et al., 1996; Lima et al., 1998). Dentre as leguminosas, foram identificadas somente três espécies, pertencentes às famílias Fabaceae e Mimosaceae. Arbustos e arbóreas foram incluídos principalmente no grupo ocasional e compreenderam cerca de 15% do número total de espécies identificadas, que corresponderam apenas a cerca de 6% do peso seco da dieta. Axonopus purpussi foi a única espécie encontrada em todas as unidades amostrais, com valor mínimo de 3,4% e máximo de 50,0%. As principais espécies consumidas por bovinos ocorreram nas áreas sujeitas à inundação, que foram campo limpo, borda de baía permanente, baía temporária, vazantes e baixadas, que coincidiram com as unidades de paisagem mais usadas por bovinos para pastejo (Santos et al., 2001b). Portanto, a composição botânica da pastagem é refletida na composição da dieta animal, conseqüentemente na produção animal. Considerando que as áreas que sofrem inundação apresentam maior variação na composição botânica e como estas são as unidades de vegetação mais utilizadas por bovinos, a produção animal de uma invernada será variável de acordo com as condições climáticas.
Para todos os grupos estudados não foram detectados efeitos significativos (P>0,05) de estado fisiológico das vacas e da interação estado fisiológico com época e ano sobre a composição média da dieta. Portanto, os dados foram generalizados para o grupo de vacas como um todo. Houve efeitos significativos (P<0,05) de época e ano para todos os grupos, porém, a interação época x ano foi significativa somente para o grupo ocasional. Na Tabela 2 constam as médias dos quadrados mínimos da composição percentual dos grupos de forrageiras da dieta, de acordo com as diferentes épocas durante os dois anos hidrológicos. O grupo principal teve participação superior no ano de 1997/98 ('normal') enquanto que os demais grupos tiveram maior participação no ano 1998/99 ('atípico'), indicando influência das condições climáticas na composição botânica da dieta. Diferenças entre as médias de composição da dieta entre os dois anos foram encontradas nas diferentes épocas para todos os grupos. De maneira geral, a participação do grupo principal na dieta foi mais baixa no ano 'atípico', ao contrário dos demais grupos. A participação média do grupo principal foi menor no período de julho a setembro em ambos os anos estudados e maior no período de outubro a junho no ano 'normal' e de outubro a março no ano 'atípico'. Os grupos médio e ocasional tiveram maior participação da dieta no período de abril a setembro (época seca), concordando com diversos estudos que mostraram maior participação de ervas e arbustivas na dieta de bovinos durante a época seca (Guevara et al., 1996; Lima et al., 1998).
Na Tabela 3 constam as médias ajustadas (quadrados mínimos) dos componentes botânicos do grupo principal da dieta. A maior parte dos componentes do grupo principal foi influenciada significativamente (P<0,05) por época e ano hidrológico. As médias estimadas de A. purpusii não diferiram significativamente entre épocas no ano 1997/98, mas diferiram no ano 1998/99, cuja maior participação na dieta ocorreu de janeiro a junho. Já as médias de M. chaseae diferiram significativamente entre os períodos de janeiro a março e abril a junho. Ambas as espécies tiveram participação expressiva nos dois anos estudados. Interações época x ano só não foram significativas para S. phalerata (folhas e frutos). As espécies hidrófilas E. minima, H. amplexicaulis, L. hexandra e P. laxum tiveram participação mais expressiva na dieta no ano 1997/98, enquanto E. acutangula, no ano 1998/99.
Entre as nove espécies que compuseram o grupo principal da dieta de bovinos, cinco são hidrófilas: H. amplexicaulis, Leersia hexandra, P. laxum, E. minima e E. acutangula e dentre estas, as três primeiras apresentam via fotossintética denominada C3. Em geral, estas espécies são preferidas, provavelmente por apresentarem menos estruturas fibro-vasculares, sendo mais tenras e de maior digestibilidade. De acordo com Valls (1994), apesar de estar situado em área tropical, o Pantanal tem um número significativo de gramíneas caracterizadas pela via C3.
A Figura 2 ilustra a variação mensal das médias observadas dos componentes da dieta do grupo principal, durante os dois anos hidrológicos. P. laxum e E. minima tiveram maior participação na dieta nos meses de outubro a dezembro (início das chuvas), enquanto S. phalerata nos meses de abril a junho (pico da cheia). O consumo dos frutos de S. phalerata acompanhou o consumo das folhas. E. acutangula teve participação expressiva na dieta somente no ano 'atípico' e na época correspondente ao período de abril a junho (época da vazante), enquanto H. amplexicaulis teve participação maior no ano 'normal', na época seca, de julho a setembro. Leersia hexandra, por sua vez, teve participação constante durante o ano 1997/98, mas no ano de 1998/99 teve uma maior participação no período de julho a agosto (seco).
No ano considerado 'atípico', a participação das gramíneas hidrófilas na dieta de bovinos sofreu um decréscimo no período de janeiro a maio, provavelmente devido à inundação parcial da área neste período, porém, nesse mesmo ano, a participação teve um acréscimo durante o período seco (julho a agosto), com o abaixamento das águas. Este fato indica que estas espécies provavelmente não suportam inundação, diminuindo a sua disponibilidade. No ano 'atípico', as duas ciperáceas do grupo principal tiveram participação diferenciada na dieta, onde E. minima diminuiu e E. acutangula aumentou. Vale salientar que a inundação parcial da área ocorreu fora do período normal, que é de abril a maio, devido à intensa precipitação que ocorreu nos meses de dezembro a janeiro.
As espécies do grupo principal localizadas nas áreas mais elevadas como A. purpusii e M. chaseae tiveram uma participação menos variável na dieta. Destas espécies, destaca-se o capim mimoso (A. purpusii), pois este teve grande participação na dieta durante todo o período de estudo. Resultados semelhantes foram encontrados por Santos et al. (1999) estudando a dieta de bovinos e eqüinos numa área de máxima inundação, localizada no Pantanal da sub-região do Abobral. Esta espécie é perene, encontrada principalmente nas áreas de campo limpo, é resistente à submersão temporária, mas morre quando sujeita à inundação prolongada (Allem & Valls, 1987). Outra espécie importante foi a grama-do-cerrado (M. chaseae), muito comum nas áreas de campo limpo e de caronal. Áreas de caronal são de topografia mais elevada, porém, foram pouco utilizadas para pastejo, exceto quando ocorreu alagamento da área. Durante este estudo, parte da área ficou inundada no período de janeiro a março, no qual observou-se maior participação de Mesosetum chaseae (Figura 1) na dieta, provavelmente consumido nas áreas de caronal. O consumo mais baixo de A. purpusii nos meses de setembro a dezembro coincidiu com a proporção maior de plantas secas na pastagem. A. hypogynus teve uma participação média de 6,33% e 5,51% para o ano 'normal' e 'atípico', respectivamente. O pico de consumo desta espécie ocorreu principalmente no período pós-queima. É uma espécie encontrada principalmente nas áreas de campo limpo e de forma esparsa nas áreas de campo-cerrado e caronal.
Foi constatado um número reduzido de espécies lenhosas na dieta de bovinos, concordando com as observações efetuadas por Pott (1988). Este fato coincide com as observações diretas no campo realizadas durante o estudo. Santos et al. (2001b) avaliaram o grau de consumo das espécies presentes na área e encontraram que, de um total de 85 espécies de arbustos e arbóreas, 7 (8%) foram classificadas como 'preferida', 38 (45%) como 'desejável' e o restante como 'indesejável' e não consumidas. Observou-se diretamente que o consumo destas plantas ocorreu de forma casual (meados de setembro), e que as partes consumidas foram principalmente as folhas em brotação, ponteiros e rebrota pós-queima. Dos arbustos e arbóreas consumidos destacam-se o 'acuri' (Scheelea phalerata), 'cipó-de-fogo' (Doliocarpus dentatus), 'uva-brava' (Alchornea discolor), 'embaúba' (Cecropia pachystachya), 'olho-de-boi' (Tocoyena formosa) e 'rosquinha' (Helicteres guazumaefolia). Das espécies arbóreas identificadas na dieta destaca-se o 'acuri', presente no grupo principal da dieta de bovinos, consumida especialmente no período de abril a junho em ambos os anos estudados.
Cabe salientar que muitas espécies da área de estudo podem ter sido sub ou superestimadas pela análise microhistológica fecal. As características de diagnóstico (tricomas, estômatos etc) da epiderme são conservadas pela cutina, substância depositada sobre a superfície da célula epidérmica, extremamente resistente aos microorganismos (Cutter, 1986), cuja cutinização aumenta com o estágio de crescimento das plantas. Portanto, a porcentagem de fragmentos identificáveis nos brotos e folhas novas de muitas espécies, especialmente as arbóreas e arbustos pode estar sendo subestimada nos períodos de maior consumo. Gramíneas, por sua vez, foram identificadas com maior facilidade devido ao maior número de caracteres de diagnóstico, concordando com os resultados de Henley et al. (2001). Entretanto, de forma geral os resultados encontrados confirmam as observações diretas, onde arbustos e arbóreas foram consumidos de forma ocasional, principalmente no ano 'atípico', de janeiro a junho, enquanto que as herbáceas, principalmente gramíneas, foram as mais consumidas durante todo o período de estudo. A preferência ou não por árvores e/ou arbustos, em especial as leguminosas, pode estar relacionada com a composição química, especialmente a presença de compostos secundários que inibem o consumo efetuado pelos animais. Outro fator que, provavelmente, influenciou a não utilização destas espécies foi a adoção de uma pressão de pastejo considerada moderada a leve, a qual propiciou a manutenção das unidades de paisagem selecionadas numa condição 'boa' (não degradada) durante o período estudado.
Conclusões
Das 286 espécies presentes na área de estudo, apenas 9 foram identificadas como principais, representando cerca de 70% da composição em peso seco da dieta de bovinos. As espécies A.purpusii, M. chaseae e A. hypogynus, presentes nas cotas mais elevadas do mesorelevo tiveram participação mais expressiva e menos variável no biênio estudado. A participação de arbustos, arbóreas e leguminosas na dieta de bovinos foi verificada principalmente durante o período seco, de agosto a setembro, quando as plantas apresentaram rebrota.
Independente do estado fisiológico das vacas, a composição botânica da dieta foi similar entre os membros do grupo, indicando que a dieta depende de aprendizado.
A técnica da análise fecal pareceu apropriada para identificação da composição botânica da dieta de bovinos criados extensivamente no Pantanal, por possibilitar a confirmação das principais espécies consumidas observadas diretamente no campo.
Agradecimento
A João Batista Garcia, pela dedicação intensiva na execução deste trabalho. Ao técnico Ernande Ravaglia, aos estagiários Anderson e Átila e aos motoristas, pelo auxílio nos trabalhos de campo. Aos colegas Rodiney de Arruda Mauro e Ana Caroline Soares, pelo auxílio na aplicação da técnica microhistológica. À equipe do herbário da Embrapa-Pantanal. À Regina Célia Rachel dos Santos, que colaborou na confecção das figuras. Por fim, ao apoio financeiro da FUNDECT (Fundação de Apoio e de Desenvolvimento do Ensino, Ciência e Tecnologia) de MS e ao CNPq, pela bolsa concedida.
Literatura Citada
Recebido em: 28/08/01
Aceito em: 11/01/02
2 Pesquisadora da Embrapa Pantanal, Cx. Postal 109, 79320-900, Corumbá, MS. Bolsista do CNPq. E.mail: sasantos@cpap.embrapa.br
3 Professore da FMVZ-UNESP, Campus de Botucatu, SP. E.mail: ciniro@fca.unesp.br
4 Pesquisador da Embrapa-Secretaria de Adiministração Estratégica (SEA), Brasília, DF. E.mail: geraldo@sede.embrapa.br
5 Pesquisador da Embrapa Gado de Corte, Campo Grande, MS. E.mail: apott@cnpgc.embrapa.br
6 Estudante do CEUC-UFMS, Corumbá, MS. E.mail: alvarez01@bol.com.br
7 Professora do Instituto de Ciências Biológicas, UNESP, Campus de Botucatu, SP. E.mail: rmachado@ibb.unesp.br
- ALLEM, A.C.; VALLS, J.F.M. Recursos forrageiros nativos do Pantanal Mato-Grossense. Brasília: EMBRAPA-CENARGEN, 1987. 339p (EMBRAPA-CENARGEN, Documentos, 8).
- BÓO, R.M.; LINDSTRON, L.I.; ELÍA, O.R. et al. Botanical composition and seasonal trends of cattle diets in central Argentina. Journal of Range Management, v.46, n.6, p.479-482, 1993.
- CARVALHO FILHO, A.; CARDOSO, E.L.; NAIME, U.J. et al. Solos como fator de diferenciação fitofisionômica na sub-região da Nhecolândia ž Pantanal Mato-Grossense. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL, 3., 2000, Corumbá. Os desafios do Novo Milênio. Resumos... Corumbá: Embrapa Pantanal, p.59. 2000.
- COMASTRI-FILHO, J.A. Pastagens nativas e cultivadas no Pantanal Mato-Grossense Corumbá: EMBRAPA-CPAP, 1984. 48p. (Circular Técnica, 13)
- CUNHA, N.G. Considerações sobre os solos da sub-região da Nhecolândia, Pantanal Mato-Grossense. Corumbá: EMBRAPA-UEPAE de Corumbá, 1980. 45p. (Circular Técnica, 1).
- CUTTER, E.G. Anatomia vegetal. Parte I - células e tecidos São Paulo: Roca, 1986. 304p.
- DUARTE, C.M.L.; NASCIMENTO Jr., D.; SILVA, E.A.M. et al. Métodos para estimar a composição botânica da dieta de herbívoros. Revista da Sociedade Brasileira de Zootecnia, v.21, n.2, p.179-180, 1992.
- GUEVARA, J.C.; ESTEVEZ, O.R.; STASI, C.R. et al. Botanical composition of the seasonal diet of cattle in the rangelands of the Monte Desert of Mendoza, Argentina. Journal of Arid Environments, v.32, p.387-394, 1996.
- HENLEY, S.R.; SMITH, D.G.; RAATS, J.G. An evaluation of three techniques applied to the study of diet selection in free-ranging large herbivores, Journal of Range Management (no prelo). 2001.
- HOLECHEK, J.L.; VAVRA, M.; PIEPER, R.D. Botanical composition determination of range herbivore diets: a review. Journal of Range Management, v.3, n.3, p.309-315, 1982.
- HOLECHEK, J.L.; GROSS, B.D. Training needed for quantifying simulated diets from fragmented range plants. Journal of Range Management, v.35, n.5, p.644-647, 1982a.
- HOLECHEK, J.L.; GROSS, B.D. Evaluation of different calculation procedures for microhistological analysis. Journal of Range Management, v.35, n.6, p.721-723, 1982b.
- LIMA, J.A.; NASCIMENTO Jr., D.; PEREIRA, J.C. et al. Seletividade por bovinos em pastagem natural.1.composição botânica. Revista da Sociedade Brasileira de Zootecnia, v.27, n.3, p.434-443, 1998.
- McINNIS, M.L.; VAVRA, M. Dietary relationships among feral horses, cattle, and pronghorn in Southeastern Oregon. Journal of Range Management, v.40, n.1, p.60-66, 1987.
- METCALF, C.R. Anatomy of the monocotyledons. I. Gramineae Oxford: Clarendon Press, 1960. 731p.
- METCALF, C.R.; CHALK, L. Anatomy of the dicotyledons Oxford: Clarendon Press, 1979. v.1, 276p.
- MIDDELETON, B.A.; ROJAS, E.S. Microhistological analysis of the food habits of herbivores in the tropics. Vida Silvestre Neotropical, v.3, n.1, p.41-47, 1994.
- NORBURY, G.L. Microscopic analysis of herbivore diets - a problem and a solution. Australian Wildlife Research, v.15, p.51-57, 1988.
- NORBURY, G.L.; SANSON, G.D. Problems with measuring diet selection of terrestrial, mammalian herbivores. Australian Journal of Ecology, v.17, p.1-7, 1992.
- POTT, A. Forrageiras não gramíneas dos cerrados e campos do Pantanal Mato-Grossense. In: SIMPÓSIO SOBRE O CERRADO: SAVANAS: ALIMENTO E ENERGIA, 6., 1982, Brasília. Anais... Brasília: EMBRAPA-CPACC, 1988. p.381-388.
- POTT, A. Pastagens nativas. In: Tecnologias e informações para a pecuária de corte no Pantanal. Corumbá, MS: EMBRAPA-CPAP, 1997. 161p.
- POTT, A.; Da SILVA, J.V.; ABDON, M. et al. Vegetação. In: BRASIL. Ministério do Meio Ambiente, dos Recursos Hídricos e da Amazônia Legal. Plano de Conservação da Bacia do Alto Paraguai (pantanal) - PCBAP. Diagnósticos dos meios físicos e bióticos: meio biótico. Brasília, v.2, n.3, p.1-179, 1997.
- POTT, A.; POTT, V.J. Flora do Pantanal - listagem atual de fanerógamas. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL, 2., 1996, Corumbá. Anais... Corumbá: Embrapa Pantanal, 1999. p.297-325.
- PROVENZA, F.D. Viewpoint: range science and range management are complementary but distinct endeavors. Journal of Range Management, v.44, n.2, p.181-183, 1991.
- SANTOS, S.A.; SOARES, A.C.; CRISPIM, S.M.A. et al. Índices de sobreposição e preferência alimentar de bovinos e equinos mantidos em pastagens nativas no Pantanal do Abobral, MS. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL, 2., 1996, Corumbá. Manejo e Conservação. Anais... Corumbá: Embrapa Pantanal, 1999.
- SANTOS, S.A.; COSTA, C.; POTT, A. et al. Test of a conversion equation to increase the accuracy of microhistological analysis of herbivore diets in the Pantanal. In: INTERNATIONAL GRASSLAND CONGRESS, 19., 2001, São Pedro, SP. Proceedings. Piracicaba: FEALQ, 2001a. p.337-338.
- SANTOS, S.A., COSTA, C., CRISPIM, S.M.A. et al. Seleção das fitofisionomias da sub-região da Nhecolândia, Pantanal, por bovinos. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL. 3, 2000, Corumbá, MS. Os desafios do novo milênio. Anais... Corumbá: Embrapa Pantanal, CD ROOM. 2001b.
- SAS INSTITUTE INC. System for Microsoft 0windows, Release 6.12, Cary, NC, USA, CD ROOM. 1999.
- SCOTT, G., DAHL, B.E. Key to selected plant species of Texas using plant fragments. Lubbock, Texas, U.S.A.:Texas Tech. Press. 1980. 9p.
- SPARKS, D.R., MALECHEK, J.C. Estimating percentage dry weight in diets using a microscope technique. Journal of Range Management, v.21, n.4, p.264-265, 1968.
- SORIANO, B.M.A. Caracterização climática da sub-região da Nhecolândia, Pantanal-MS. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL, 2., 1996, Corumbá, MS. Manejo e Conservação. Anais... Corumbá: Embrapa Pantanal, 1999. p.151-158.
- VALLS, J.F.M. O espectro taxonômico das gramíneas do Pantanal. In: PUIGNON, J.P. (Eds.) Utilizacion y manejo de pastizales Montevideo: IICA-PROCISUR, 1994. p.227-237 (IICA - PROCISUR. Dialogo, 40).
Datas de Publicação
-
Publicação nesta coleção
09 Set 2005 -
Data do Fascículo
Jul 2002
Histórico
-
Aceito
11 Jan 2002 -
Recebido
28 Ago 2001