Resumos
Objetivou-se avaliar a distribuição dos fotoassimilados pelos perfilhos do cultivar Mombaça de Panicum maximum. Os tratamentos experimentais resultaram de um arranjo fatorial 2x2x3 - dois níveis de inserção da folha (topo e base), dois estádios de desenvolvimento da planta e três momentos de colheita (3, 8 e 24 horas após a exposição da folha ao 14CO2). O delineamento utilizado foi inteiramente casualizado, com três repetições. A exposição ao 14CO2 teve duração de 20 minutos. Cada planta foi desmembrada em perfilho principal e perfilhos surgidos no primeiro e segundo estádios de desenvolvimento. O perfilho principal foi dividido em: folha adulta, completamente expandida exposta ao 14CO2 (bainha mais lâmina), demais folhas adultas (bainha mais lâmina), meristema terminal (folha emergente mais folha em expansão mais meristema apical), base e raiz. A atividade das amostras foi determinada em espectrômetro de cintilação líquida. A percentagem de fotoassimilados 14C retida na folha exposta não variou com o tempo de colheita, após exposição. Mais da metade da radioatividade recuperada na planta foi retida no perfilho principal. A folha do topo reteve mais assimilados que a folha da base, que translocou maior percentual de assimilados para os perfilhos que a folha do topo. O percentual de assimilados retidos no perfilho principal foi mais alto em plantas do primeiro estádio de desenvolvimento em relação às do segundo estádio. As folhas adultas e a raiz do perfilho principal se constituíram nos seus principais drenos. Apreciável percentagem de assimilados marcados foi encontrada nas folhas adultas não-expostas do perfilho principal.
dreno; fonte; translocação de assimilados
The objective of this work was to evaluate the distribution of assimilates produced by the top and bottom leaves of the main tiller in plants of Monbaçagrass (Panicum maximum), in two stages of plant development, and three harvest times after plant exposure to 14CO2 (three, eight and twenty four hours). The experimental treatments were arranged in a 2X2X3 factorial combination. The experimental design utilized was completely randomized. The exposure time to 14CO2 was 20 minutes. Each plant was separated into shoot (main tiller) and tillers developed in the first and second stages of plant development. The shoot was dissected into the following fractions: fully expanded leaves (sheath plus blade), fed leaf (sheath plus blade), terminal meristem (expanding leaves plus emerging leaves plus apical meristem), base and root system. The radioactivity of the samples was determined in a liquid scintillation spectrometer. The percentage of 14C-assimilates retained in the fed leaf did not vary with time of harvesting after exposure. Most of the plant 14C assimilates were retained in the main tiller, mainly when the fed leaf was the top one, which retained more assimilates than the bottom leaf. The bottom leaf translocated more assimilates to tillers than the top leaf. Percentage assimilates retained in main tiller was higher in plants of the first developmental stage relative to the second one. Sizeable amount of activity was found in non-fed adult leaves of the main tiller, mainly when the bottom leaf was the fed one.
source; sink and translocation
FORRAGICULTURA
Distribuição de fotoassimilados de folhas do topo e da base do capim-mombaça (Panicum maximum Jacq.) em dois estádios de desenvolvimento1 1 Parte da dissertação do primeiro autor, bolsista do CNPq, parcialmente financiada pela Fapemig.
Distribution of assimilates from top and bottom leaves of mombaçagrass (Panicum maximum Jacq) in two stages of development
Alice Cristina Bittencourt TeixeiraI; José Alberto GomideII; Juraci Alves de OliveiraII; Emerson AlexandrinoIII; Daniel Carlos Ferreira LanzaIV
IEstudante de mestrado do departamento de Zootecnia, UFV-Viçosa-MG; bolsista do CNPq
IIProfessores da UFV (jagomide@ufv.br)
IIIDoutorando bolsista do CNPq
IVEstagiário do departamento de Fisiologia Vegetal-UFV
RESUMO
Objetivou-se avaliar a distribuição dos fotoassimilados pelos perfilhos do cultivar Mombaça de Panicum maximum. Os tratamentos experimentais resultaram de um arranjo fatorial 2x2x3 - dois níveis de inserção da folha (topo e base), dois estádios de desenvolvimento da planta e três momentos de colheita (3, 8 e 24 horas após a exposição da folha ao 14CO2). O delineamento utilizado foi inteiramente casualizado, com três repetições. A exposição ao 14CO2 teve duração de 20 minutos. Cada planta foi desmembrada em perfilho principal e perfilhos surgidos no primeiro e segundo estádios de desenvolvimento. O perfilho principal foi dividido em: folha adulta, completamente expandida exposta ao 14CO2 (bainha mais lâmina), demais folhas adultas (bainha mais lâmina), meristema terminal (folha emergente mais folha em expansão mais meristema apical), base e raiz. A atividade das amostras foi determinada em espectrômetro de cintilação líquida. A percentagem de fotoassimilados 14C retida na folha exposta não variou com o tempo de colheita, após exposição. Mais da metade da radioatividade recuperada na planta foi retida no perfilho principal. A folha do topo reteve mais assimilados que a folha da base, que translocou maior percentual de assimilados para os perfilhos que a folha do topo. O percentual de assimilados retidos no perfilho principal foi mais alto em plantas do primeiro estádio de desenvolvimento em relação às do segundo estádio. As folhas adultas e a raiz do perfilho principal se constituíram nos seus principais drenos. Apreciável percentagem de assimilados marcados foi encontrada nas folhas adultas não-expostas do perfilho principal.
Palavras-chave: dreno, fonte, translocação de assimilados
ABSTRACT
The objective of this work was to evaluate the distribution of assimilates produced by the top and bottom leaves of the main tiller in plants of Monbaçagrass (Panicum maximum), in two stages of plant development, and three harvest times after plant exposure to 14CO2 (three, eight and twenty four hours). The experimental treatments were arranged in a 2X2X3 factorial combination. The experimental design utilized was completely randomized. The exposure time to 14CO2 was 20 minutes. Each plant was separated into shoot (main tiller) and tillers developed in the first and second stages of plant development. The shoot was dissected into the following fractions: fully expanded leaves (sheath plus blade), fed leaf (sheath plus blade), terminal meristem (expanding leaves plus emerging leaves plus apical meristem), base and root system. The radioactivity of the samples was determined in a liquid scintillation spectrometer. The percentage of 14C-assimilates retained in the fed leaf did not vary with time of harvesting after exposure. Most of the plant 14C assimilates were retained in the main tiller, mainly when the fed leaf was the top one, which retained more assimilates than the bottom leaf. The bottom leaf translocated more assimilates to tillers than the top leaf. Percentage assimilates retained in main tiller was higher in plants of the first developmental stage relative to the second one. Sizeable amount of activity was found in non-fed adult leaves of the main tiller, mainly when the bottom leaf was the fed one.
Key Words: source, sink and translocation
Introdução
Cerca de 90% da massa seca vegetal são constituídos por compostos oriundos da fotossíntese (Zelitch, 1982). A biomassa vegetal pode ser limitada tanto pela produção como pelo uso dos fotoassimilados por parte dos meristemas foliares (Lemaire & Agnusdei, 1999).
A taxa fotossintética da folha determina a quantidade total de carbono fixada disponível ao metabolismo vegetal (Taiz & Zeiger, 1998) e é função de fatores intrínsecos à planta, como nível de inserção da folha, (Ryle & Powell, 1974), idade da folha (Woledge, 1985), e fatores extrínsecos à planta, como irradiância (Woledge, 1985), água no solo, temperatura e nutrientes (Larcher, 1995). Portanto, as folhas de mais alto nível de inserção no perfilho, mais jovens e mais bem iluminadas, fotosintetizam mais e contribuem mais para o pool de fotoassimilados da planta que as folhas baixeiras, mais velhas (Robson, 1988).
O carbono fixado pelas folhas pode ser alocado para três caminhos metabólicos: (i) para utilização no metabolismo celular, fornecendo energia e esqueletos de carbono para a síntese de outros compostos, (ii) para a síntese de compostos de transporte, exportados para os diversos drenos da planta e (iii) para síntese de compostos armazenados, para utilização durante a noite (Taiz & Zeiger, 1998).
O meristema terminal (meristema apical, folhas em expansão e folhas emergentes), os perfilhos, o colmo, a raiz, a inflorescência e as sementes constituem-se nos diversos drenos da gramínea (Robson, 1988).
A partição de fotoassimilados acompanha a força do dreno, que é produto de seu tamanho (peso total do tecido do dreno) e de sua atividade (taxa de absorção de assimilados por unidade de peso do tecido) (Taiz & Zeiger, 1998). Além disso, a distância do dreno à fonte, a competição entre eles e a arquitetura vascular de folhas e colmos têm grande influência sobre a partição de fotoassimilados em plantas intactas, crescendo sob condições não-limitantes (Chapman et al. 1991a,b; Matthew & Kemball, 1997).
Estudos de partição de fotoassimilados são fundamentais para o entendimento da fisiologia vegetal, pois a dinâmica da relação fonte/dreno varia conforme o meio, o estádio fisiológico da planta e o manejo da cultura.
Após a desfolha, os assimilados recém-sintetizados pela área foliar residual e as reservas orgânicas respondem pela recomposição da área foliar, priorizando os meristemas terminais, além de assistirem nas perdas respiratórias da planta (Schnyder & De Visser, 1999).
O conhecimento da partição e da alocação de assimilados pode subsidiar o manejo do cultivo, objetivando sua produtividade, isto é, seu índice de colheita (Beadle, 1993). O manejo da pastagem deve ser orientado no sentido de favorecer a produção e o acúmulo de folhas, em razão não apenas do papel delas na economia da forrageira como também por constituírem a fração mais nutritiva da dieta do ruminante.
Inúmeros trabalhos têm sido realizados para investigar a partição de fotoassimilados em gramíneas do grupo C3 e leguminosas de clima temperado (Ryle & Powell, 1974; Chapman et al., 1991a,b). Nas regiões de clima temperado, predominam leguminosas, como o trevo branco (Trifolium repens L.), trevo vermelho (Trifolium pratense L.) e gramíneas de metabolismo C3, como o azevém anual (Lolium multiflorum L.) e azevém perene (Lolium perenne L.). As gramíneas de metabolismo C4, como as tropicais estudadas, diferem por serem anuais, como o sorgo (Sorghum bicolor L.) e o milho (Zea mays L.), que não perfilha. Os estudos sobre a relação fonte/dreno de gramíneas forrageiras tropicais são escassos.
Avaliou-se, neste trabalho, o destino dos fotoassimilados sintetizados em folhas de diferentes níveis de inserção no perfilho principal de Panicum maximum cv. Mombaça, em dois estádios de desenvolvimento, em diferentes tempos de colheita, após a exposição da folha ao 14CO2.
Material e Métodos
O capim Panicum maximum cv. Mombaça foi cultivado em casa de vegetação do Departamento de Biologia Vegetal da Universidade Federal de Viçosa (UFV). A semeadura foi realizada em 25/10/2002 em bandeja plástica, perfurada e contendo areia, irrigada três vezes ao dia. Após a germinação, uma semana antes do transplantio, foram realizadas duas aplicações com solução de Hoagland, com um quarto de força iônica e pH 5,5 (Hoagland & Arnon, 1950). O transplantio de três plântulas por vaso foi realizado quando atingiram, além da folha cotiledonar, duas folhas completamente expandidas. Efetuou-se o desbaste, mantendo-se uma planta por vaso, quando as plântulas haviam atingido a terceira folha completamente expandida. Uma mistura de solo e areia na proporção 1:1 foi o substrato utilizado para o enchimento dos vasos. A adubação fosfatada foi feita em dose única de 400 mg.dm-3 de solo, enquanto a potássica e a nitrogenada foram realizadas de forma parcelada, atingindo concentrações de 250 mg.dm-3 e 60 mg.dm-3, respectivamente. A adubação com micronutrientes baseou-se na análise química do solo.
Os tratamentos resultaram de um arranjo fatorial 2x2x3 - dois níveis de inserção de folha no perfilho principal (topo e base), dois estádios de crescimento e três momentos de colheita (três, oito, e 24 h) após a exposição da folha ao 14CO2. Os estádios de crescimento corresponderam: (i) ao momento em que o perfilho principal alcançou valores próximos a nove folhas completamente expandidas e quatro a cinco perfilhos; (ii) dez folhas completamente expandidas e oito a nove perfilhos, respectivamente. O primeiro estádio foi atingido em 09/12/2002, cerca de 25 dias após o transplantio, e o segundo, 11 dias depois (20/12/2002). O delineamento foi inteiramente casualizado, perfazendo 12 tratamentos, com três repetições, totalizando 36 unidades experimentais (vasos com uma planta). Durante o desenvolvimento das plantas, seus perfilhos foram identificados, utilizando-se anéis coloridos, de modo a se reconhecer sua ordem de surgimento.
Ao final de cada estádio de crescimento, metade dos vasos (18 vasos) foi conduzida ao laboratório para exposição da folha, por 20 min, ao 14CO2. A exposição, nos dois estádios de crescimento, foi feita em dois dias consecutivos, sendo expostas nove plantas por dia.
Apenas metade superior da lâmina foliar do topo (folha mais jovem completamente expandida) ou da base (folha mais velha sem sintoma visível de senescência) de cada planta foi exposta ao 14CO2. No momento da exposição da lâmina, o perfilho principal já havia alcançado número constante de três folhas, após atingir mais de seis folhas completamente expandidas. A folha a ser exposta foi encapsulada em um saco plástico, onde 25 mL de NaH14CO3 (5 mCi.10 mL-1) em contato com ácido lático em excesso produziram o 14CO2.
Durante o processo de exposição, as plantas foram submetidas a um regime de luz constante. Foram empregadas como fonte de luz duas lâmpadas de halogênio de 1000 Watts que conferiram, à folhagem dos vasos, radiação fotosinteticamente ativa (RFA) de 600 mmols.m-2.seg-1. Após a exposição, os vasos foram levados à casa de vegetação, onde ficaram sob luz natural até sua colheita.
As plantas, uma por vaso, foram colhidas três, oito ou 24 horas após a exposição ao 14CO2. Depois de serem colhidas, cada planta teve seu perfilho principal separado da raiz e dos demais perfilhos. Posteriormente, o perfilho principal foi dissecação em: folha completamente expandida exposta (bainha mais lâmina), folhas adultas não expostas (bainha mais lâmina), meristema terminal (folha emergente mais folha em expansão mais meristema apical), base e raiz. Os demais perfilhos foram desmembrados em perfilhos primários e secundários surgidos no primeiro e segundo estádios. Em seguida, as partes da planta estudada foram levadas à estufa, a 100ºC por 20 minutos e, posteriormente, foram secas em estufa a 75ºC, durante três dias.
As partes foram pesadas separadamente e posteriormente moídas. As amostras de raiz, perfilhos surgidos na primeira e segunda fases e as folhas completamente expandidas (não expostas) foram trituradas em moinho de faca, em ordem crescente de atividade esperada. Em seguida, uma amostra menor e representativa de cada parte foi retirada e passada em moinho de bola, para se obter um material mais fino. A base, o meristema terminal e a folha exposta foram moídos diretamente em moinho de bola. Após a moagem, foram retiradas amostras de aproximadamente 10 mg para a determinação de 14C.
A amostra foi colocada em frasco de vidro e nele foram adicionados 2 mL de detergente (Triton-X), seguindo-se homogeneização por 15 segundos. Em seguida, foram adicionados 10 mL de coquetel de cintilação [5 g 2,5 difenil oxazol (PPO), 10 g naftaleno e 100 mL de dioxano] mais dispersante (Cab-o-sil), seguindo-se nova homogeneização, por 15 segundos. A radioatividade de cada amostra foi medida em espectrômetro Beckman 6500 de cintilação líquida, por cinco minutos, utilizando-se janela completamente aberta. A partir da atividade da amostra, foram determinadas as atividades totais de cada fração (órgão) da planta.
Os dados experimentais foram submetidos à análise de variância, segundo o modelo:
Yijkl = µ + Ii + Ej + IEij + Tk + ITik + ETjk + e(ijkl)l
em que i = 1,2 níveis de inserção (fixo); j = 1,2 estádios de desenvolvimento (fixo); k = 1,2,3 tempos após a exposição ao 14CO2 (fixo); l = 1,2,3 repetições (aleatórias).
Resultados e Discussão
As maiores biomassas da planta do capim-Mombaça (Tabela 1) corresponderam aos perfilhos primários e secundários, ao sistema radicular e às folhas adultas do perfilho principal. Estas frações constituíram seus principais drenos, onde se acumularam as maiores quantidades exportadas de fotoassimilados-14C. Dessa forma, o tamanho do órgão foi fator determinante da sua força dreno, provavelmente, porque as plantas se apresentavam ainda muito jovens, nos dois estádios estudados, e estavam com seus órgãos em pleno crescimento e grande atividade metabólica.
Carvalho (2002) observou, nos cultivares Mombaça e Tanzânia de Panicum maximum, que, quando todo o perfilho principal foi exposto ao 14CO2, 6,5% dos fotoassimilados-14C estavam presentes no perfilho primário jovem mais próximo. Contudo, quando o perfilho primário mais jovem foi exposto ao 14CO2, 14% dos fotoassimilados-14C se encontravam presentes no perfilho principal. A distância e a maior força dreno do perfilho principal (o maior perfilho) teriam sido os principais fatores a determinar a maior quantidade de fotoassimilados-14C exportada para o mesmo, em relação às demais categorias de perfilhos. Entretanto, Alexandrino (2004) não encontrou a mesma proporcionalidade quando a folha completamente expandida mais jovem do maior perfilho de Panicum maximum, cultivar Mombaça, em um estádio mais avançado de desenvolvimento, foi exposta ao 14CO2. Dos fotoassimilados encontrados na planta, 8,48 e 7,63% foram recuperados no colmo e nos perfilhos maiores, que representavam, respectivamente, 0,84 e 51,4% da biomassa seca total da planta. Carvalho (2002) destaca a importância não só do tamanho do dreno, mas também de sua atividade na determinação de sua força.
Neste trabalho, não houve interação de primeira ordem (P>0.05) dos fatores estudados sobre a percentagem de fotoassimilados-14C presentes em cada órgão da planta. A ausência de interações pode ser decorrente da proximidade entre os dois estádios estudados e da idade das plantas (ainda muito jovens e em plena fase de estabelecimento), além da alta velocidade de translocação dos assimilados em plantas C4.
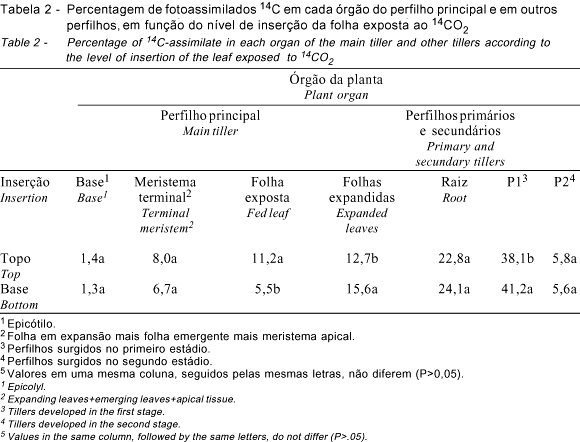
A quantidade de fotoassimilados 14C presente na folha exposta, nas demais folhas adultas do perfilho principal e nos perfilhos surgidos na primeira fase (P1), sofreu influência (P<0,05) do nível de inserção da folha (Tabela 2).
A folha do topo, por apresentar maior massa, reteve mais fotoassimilados que a da base (Tabela 3). No entanto, essas folhas expostas ao 14CO2 diferiam não apenas quanto à massa e ao nível de inserção no perfilho, mas também à idade e ao ambiente em que se encontravam. As folhas da base, mais velhas e desenvolvidas sob condições de baixa luminosidade, têm menor capacidade fotossintética.
Ryle (1970) avaliou em Lolium perenne L., no estádio de desenvolvimento vegetativo, o efeito da idade da folha na percentagem de fotoassimilados-14C retidos na mesma. Maior percentagem de fotoassimilados-14C foi retida pela folha exposta recém-expandida que pela folha exposta mais velha, com duas novas folhas acima dela.
Ryle & Powell (1972) estudaram mais detalhadamente o efeito da inserção e idade da folha na quantidade de fotoassimilados nas folhas de Lolium temulentum L., após 24 horas de exposição, durante os estádios vegetativo e reprodutivo. Maior quantidade de fotoassimilados-14C ficou retida na folha exposta, recém-expandida, havendo queda na quantidade retida uma semana depois, quando uma nova folha se expandiu acima dela. Entretanto, com o decorrer do tempo, houve aumento na quantidade de fotoassimilados retidos na folha, sem todavia, ultrapassar os valores nela presentes, quando se encontrava recentemente expandida.
Ryle (1970), estudando o efeito da inserção da folha no perfilho, observou maior retenção de fotoassimilados-14C na folha do topo em relação à da base.
O autor argumentou que somente 15 cm da folha foram expostos ao 14C, embora o tamanho dessas folhas tivesse aumentado, à medida que elas ocupavam um nível de inserção mais elevado no perfilho. Conseqüentemente, a área exposta não variou e a quantidade de fotoassimilados-14C retidos foi maior quanto maior o tamanho da folha.
Segundo Gomide & Gomide (2000), as lâminas foliares do cultivar Mombaça mostraram comprimento crescente à medida que se sucediam no perfilho, atingindo valor máximo por volta da oitava ou décima folha no perfilho principal. Desta maneira, no presente trabalho, em que a planta se encontrava nos estádios de nove e dez folhas completamente expandidas no perfilho principal, a folha do topo apresentava-se maior que a da base. No entanto, como não houve acompanhamento da área da folha exposta ao 14CO2, não é aconselhável conjeturar sobre o efeito do tamanho da folha na percentagem de radioatividade retida.
As folhas do topo e da base diferem em idade e proximidade dos drenos da planta. A folha do topo, mais jovem, encontra-se mais próxima do meristema apical, ao passo que a da base, mais velha, localiza-se mais próxima da raiz e dos perfilhos, o que lhes confere diferente padrão de distribuição de assimilados, que este é resultado da força e da distância dos drenos à fonte (Ryle, 1970). Entretanto, em decorrência dos estádios de desenvolvimento das plantas deste estudo, plantas jovens com colmo ainda não alongado e em estádio de estabelecimento e pleno perfilhamento, observou-se diferença na exportação de assimilados somente para as folhas adultas e os perfilhos surgidos na primeira fase, dois dos três grandes drenos da planta (Tabela 2) (P<0,05). Menor percentagem de radioatividade foi encontrada nas folhas adultas não expostas e nos perfilhos primários e secundários do primeiro estádio, quando a folha do topo foi exposta. A quantidade de fotoassimilados exportada para a raiz, segundo maior dreno da planta, tendeu a ser maior quando da exposição da folha da base, porém, sem significância (P>0,05) para a diferença observada (Tabela 2).
A quantidade de fotoassimilados presente no perfilho principal alcançou percentual mais alto (56,1%), quando da exposição da folha do topo, que a observada quando da exposição da folha da base, que foi de 53,2% (P<0,05%) (Tabela 2). Esse fato reflete a menor retenção de fotoassimilados na folha exposta da base e sua maior exportação de fotoassimilados para os perfilhos surgidos na primeira fase, maiores drenos da planta, ficando menos fotoassimilados no perfilho principal.
Ryle (1970) encontrou diferença na distribuição de assimilados de acordo com a inserção da folha, em Lolium perene L. e Lolium temulentum L. durante os estádios de desenvolvimento vegetativo mais avançado e no estádio reprodutivo. As folhas de nível de inserção mais baixo exportaram assimilados predominantemente para as raízes e perfilhos enquanto aquelas de nível de inserção mais alto exportaram para o meristema terminal. Essa diferença na distribuição foi mais acentuada em perfilhos mais velhos, que apresentavam colmo alongado. Diferença na distribuição de assimilados, em função do nível de inserção da folha em milho, sorgo, trigo e azevém perene, durante o estádio vegetativo, também foi encontrada por Ryle & Powell (1974), em que a folha do topo exportou mais assimilados para o meristema terminal, enquanto a folha da base exportou para a raiz e perfilhos.
O estádio de desenvolvimento estudado influenciou a quantidade de fotoassimilados presente nos órgãos (P<0,05), exceto a base (P>0,05) (Tabela 4). Observa-se na Tabela 4 que, enquanto a percentagem de fotoassimilados presente no perfilho principal diminuiu do primeiro para o segundo estádio, aquela exportada para os perfilhos primários e secundários aumentou (P<0,05). A percentagem exportada para os perfilhos formados na primeira fase (P1) foi maior no segundo estádio que no primeiro. No entanto, apesar de esta diferença ter alcançado nível de significância, foi pequena, atribuindo-se, ao aparecimento de novos perfilhos na segunda fase (P2) a diminuição de fotoassimilados retidos no perfilho principal e o aumento nos fotoassimilados exportados para os perfilhos.
Os perfilhos formados no primeiro estádio (P1), a grande maioria primários, não apresentaram alteração substancial na participação da biomassa seca total, apesar de estarem maiores no segundo estádio. Além disso, esses perfilhos já possuíam, no segundo estádio, suficiente área foliar para fixar e exportar assimilados, estando menos dependentes do perfilho principal. Como conseqüência, a percentagem de fotoassimilados recebida pelos mesmos não teve grande aumento do primeiro para o segundo estádio, mas continuou elevada e maior que a percentagem recebida pelos perfilhos formados no segundo estádio. Segundo Ryle & Powell (1972), maior porcentagem de fotoassimilados é exportada para os perfilhos primários de maior porte.
Vale enfatizar que os perfilhos desenvolvidos no segundo estádio (P2) ainda estavam pequenos, representando somente 12,8% da biomassa total da planta. Esses perfilhos eram, em sua maioria, secundários, ou seja, formados nas axilas dos perfilhos primários, de modo que, para que o assimilado produzido pelo perfilho principal pudesse alcançá-los, teria que passar pelos primários. Como conseqüência de sua menor massa e distância do perfilho principal, portador da folha exposta ao 14CO2, houve menor exportação para os mesmos. De acordo com Carvalho (2002), no caso dos cultivares Mombaça e Tanzânia, em que todo o perfilho principal foi exposto, foi observada diminuição gradual da porcentagem de fotoassimilados 14C presente nos perfilhos, proporcionalmente às suas idades e distâncias do perfilho principal. Dessa forma, a porcentagem exportada para os perfilhos formados no segundo estádio foi bem menor que a exportada para aqueles formados no primeiro estádio.
Na Tabela 4, avaliando-se mais detalhadamente a distribuição de fotoassimilados 14C na planta, é possível observar que os perfilhos primários e secundários foram beneficiados no segundo estádio, em detrimento da folha exposta e, principalmente, das folhas adultas do perfilho principal. As percentagens de fotoassimilados 14C presentes na folha exposta, nas folhas adultas e na raiz foram maiores na primeira fase, respectivamente de 10,4; 18,7 e 25,2% que na segunda fase, que foram, respectivamente, de 6,3; 9,6 e 21,7% (P<0,05).
Nos estádios estudados neste trabalho, de nove e dez folhas completamente expandidas no perfilho principal, a folha do topo foi maior que a da base e a da segunda fase maior que a da primeira, o que pode ser inferido de suas respectivas massas. Porém, como as plantas ainda estavam muito jovens em ambos os estádios, a quantidade de fotoassimilados-14C retida na folha exposta correlacionou-se não só com seu tamanho, como também com a participação desta na planta como um todo. Ryle (1972) encontrou diferença na percentagem de fotoassimilados-14C na folha exposta em função dos estádios de desenvolvimento estudados. Com exceção da folha mais jovem completamente expandida, as demais folhas retiveram maior quantidade de fotoassimilados-14C no estádio vegetativo que no reprodutivo.
A base da planta, constituída basicamente pelo epicótilo, não sofreu influência de nenhum dos fatores estudados ou de interação entre os mesmos (P>0,05), o que, provavelmente ocorreu porque, além de esse órgão representar pequeno percentual da matéria seca da planta, corresponde somente a um canal de passagem entre a parte aérea e a raiz.
As folhas adultas do perfilho principal constituíram seu segundo maior dreno, contendo elevada percentagem de fotoassimilados-14C, principalmente no primeiro estádio, que pode ser atribuída tanto aos perfilhos em desenvolvimento nas suas axilas como à grande biomassa dessas folhas.
Em muitos estudos de partição de assimilados, a percentagem de -14C exportada para as folhas adultas é irrisória, fazendo com que não sejam consideradas drenos. De fato, segundo Fetene et al. ( 1997), a folha completamente expandida não importa, somente exporta fotoassimilados. Todavia, Ryle (1970) encontrou pequeno percentual de radioatividade em folhas adultas de Lolium perenne e L. temulentum, tanto no estádio reprodutivo como no vegetativo. Porém, esses estudos são realizados em sua maioria com plantas de clima temperado, que possuem metabolismo diferente das plantas tropicais, do grupo C4.
A distribuição de biomassa total na planta entre folhas, pseudocolmos e sistema radicular, em Panicum maximum, cultivar Moçamba, em dois estádios de desenvolvimento, equivalente à deste trabalho, foi estudada por Gomide et al. (2003), que observaram que, nas duas primeiras semanas após a emergência de quatro perfilhos primários, 14% da biomassa total foram alocados para o sistema radicular, enquanto 29 e 57,5% foram alocados para o pseudocolmo e folhas adultas, respectivamente, evienciando que, nesse período inicial, o meristema apical e as folhas em expansão, seguidos do perfilhamento, constituíram-se no principal dreno. No 37° dia do estabelecimento, após o desenvolvimento de quatro novos perfilhos, as folhas, o pseudocolmo e o sistema radicular representaram, respectivamente, 51, 41 e 8% da biomassa da planta. Houve, portanto, da primeira para a segunda fase, em termos percentuais, intensificação da alocação de assimilados para o perfilhamento, em detrimento da alocação de assimilados principalmente para o sistema radicular. O mesmo comportamento foi observado com o aumento na relação parte aérea/sistema radicular e com a queda na relação folha/colmo no período de 21 dias entre o 16º e o 37º dia de estabelecimento, o que reflete acréscimo no número de perfilhos e folhas em razão da alocação preferencial para os mesmos. Dessa forma, em plantas jovens em pleno desenvolvimento, em que todos os drenos estão em pleno crescimento, a distribuição da biomassa mostra-se bem representativa da exportação de assimilado.
Na Tabela 4, é possível observar maior presença de fotoassimilados-14C no meristema terminal no segundo estádio de desenvolvimento (P<0,05). Da radioatividade recuperada na planta, 6,2 e 8,4% foram encontrados no meristema terminal, respectivamente no primeiro e segundo estádios, provavelmente em decorrência das condições de baixa luminosidade a que as plantas estiveram submetidas da primeira para a segunda fase. Nessas condições, a parte aérea, as folhas e o colmo são beneficiados em detrimento do sistema radicular (Dias Filho, 1999) e do perfilhamento (Davies et al., 1983). Portanto, a formação da área foliar ocorre principalmente a partir da emissão de novas folhas durante o perfilhamento e da priorização na alocação de assimilados para o meristema terminal.
A quantidade de fotoassimilados-14C presente na folha exposta foi pequena (Tabela 4). A força do dreno interfere não apenas na distribuição de assimilados como também na quantidade de assimilados exportados pela fonte. Neste experimento, a radiação global incidente sobre a superfície da folha apresentou-se baixa nos dias 9, 10 e 20, 21 de dezembro, que antecederam à exposição das folhas ao 14CO2. Essas condições teriam comprometido a eficiência fotossintética das folhas e favorecido a exportação de fotoassimilados da folha exposta. Ryle & Powell (1976) observaram queda na quantidade de fotoassimilados -14C retida na folha exposta, quando submetida a um ambiente de menor luminosidade. Alexandrino (2004) obteve retenção equivalente a 56,9% de fotoassimilados14C na folha do cultivar Mombaça, 24 horas após a exposição ao 14CO2. Esse valor, bem superior ao encontrado neste experimento, foi, provavelmente, resultante da menor demanda dos drenos. Essas plantas estavam em avançado estádio de desenvolvimento e sob temperaturas amenas, apresentando, portanto, menor atividade metabólica.
A quantidade de fotoassimilados-14C exportada do perfilho principal para os perfilhos primários e secundários também foi elevada (Tabela 4), provavelmente em razão da condição de baixa luminosidade que antecedeu a exposição ao 14CO2 nos dois estádios de desenvolvimento, o que pode ter elevado ainda mais a dependência de assimilados por parte do maior dreno da planta (os perfilhos) (Ryle & Powell, 1976).
Acerca do efeito do tempo de colheita após a exposição sobre a porcentagem de fotoassimilados-14C presente nos órgãos estudados, apenas houve diferença estatística na percentagem de fotoassimilados-14C exportados para o meristema terminal do perfilho principal (P<0,05), que cresceu até a 8ª hora pós-exposição ao 14CO2, enquanto, no sistema radicular, a quantidade mostrou-se inconsistente, com valor menor (P>0,05) após 8 horas de exposição ao 14CO2 (Tabela 5).
A percentagem de fotoassimilados-14C retida na folha exposta permaneceu constante, não variando(P>0,05) com os tempos de colheita pós-exposição estudados. Segundo Taiz & Zeiger (1998), a velocidade de translocação de assimilados é, em média, de 1 m.h-1, variando em torno de 0,3 a 1,5 m.h-1. De acordo com Ryle & Powell (1974), a translocação de assimilados das duas últimas folhas recém-expandidas para o meristema apical e as raízes, nos primeiros 25 minutos após exposição ao 14CO2, foi mais alta em plantas de milho e sorgo que em plantas de trigo e azevém.
A gramínea, ora, estudada, uma planta de metabolismo C4, possuiria, então, elevada taxa de exportação inicial. Associada à elevada velocidade de translocação de assimilados no cultivar, enfatiza-se que, nos estádios estudados, a planta se encontrava em fase de estabelecimento e de intenso perfilhamento. Além disso, o período de crescimento ocorreu em época favorável do ano (verão), condição que intensificaria ainda mais a sua velocidade de translocação. Portanto, infere-se que a velocidade de translocação de assimilados foi o fator determinante da estabilização da quantidade de 14C retida na folha exposta, após três horas à sua exposição ao 14CO2. Em estudos da translocação de fotoassimilados na cultivar Mombaça, deve-se observar intervalo de tempo menor que três horas após a exposição.
Conclusões
A quantidade de assimilados retidos na folha exposta ao 14CO2 foi influenciada pelo seu nível de inserção e estádio de desenvolvimento. A folha exposta reteve mais assimilados quando localizada em nível de inserção mais alto no perfilho e no primeiro estádio de desenvolvimento.
As folhas do topo e da base mostraram modelo diferente de distribuição de assimilados. A folha da base exportou mais assimilados para os perfilhos surgidos na primeira fase e para as folhas adultas.
Na fase de pleno desenvolvimento vegetativo, os principais drenos da gramínea foram: perfilhos, raiz, folhas adultas e meristema terminal (meristema apical mais folhas folha em expansão mais folha emergente), nesta ordem.
Após o sistema radicular, as folhas adultas constituíram o segundo maior dreno do perfilho principal, fato até então ainda não registrado em literatura pertinente a plantas do grupo C3.
Literatura Citada
Recebido em: 02/03/04
Aceito em: 29/12/04
- ALEXANDRINO, E. Translocação de assimilados em capim Panicum maximum cv Mombaça, crescimento, características estruturais da gramínea e desempenho de novilhos em piquetes sob pastejo de lotação intermitente Viçosa, MG: Universidade Federal de Viçosa. 2004. 123p. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa. 2004.
- BEADLE, C.L. Growth analysis. In: HALL, D.O.; BOLHARNORDENKAMPF, H.F.; LEEGOOD, R.C. (Eds.) Photosynthesis and production in a chaging environment London: Pergamon Prees, 1993. p.36-45.
- CARVALHO, D.D. Leaf morphogenesis and tillering behaviour in single plants and simulated swards of guinea grass (Panicum maximum jacq) cultivars New Zealand: University Palmerston 2002. 186p. Tese (PhD Plant Sience) - University Palmerston, 2002.
- CHAPMAN, D.F.; ROBSON, M.J.; SNAYDON, R.W. The influence of leaf position and defoliation on the assimilation and translocation of carbon in white clover (Trifolium repens L.). 1. Carbon distribution patterns. Annals of Botany, v.67, n.4, p.295-302, 1991a
- CHAPMAN, D.F.; ROBSON, M.J.; SNAYDON, R.W. Quantitative carbon distribution in clonal plants of white clover (Trifolium repens): source sink relationship during undisturbed growth. The Journal of Agricultural Science v.116, n.2, p.229-238, 1991b.
- DAVIES, A.; EVANS, M.E.; EXLEY, J.K. Regrowth of perennial ryegrass (Lolium perenne L.). as affected by simulated leaf sheaths. The Journal of Agricultural Science, v.101, n.1, p.131-137, 1983.
- DIAS-FILHO, M.B. Physiological responses of two tropical weeds to shade 1- growth and biomass allocation. Pesquisa Agropecuária Brasileira, v.34, n.6, p.945-952, 1999.
- FETENE, M.; BENKER, C.; BECK, E. The pathway of assimilate flow from source to sink in Urtica dioica L., studied with 14C under ambient atmospheric condition. Annals of Botany, v.79, n.6, p.585-591, 1997.
- GOMIDE, C.A.M.; GOMIDE, J.A. Morfogênese de cultivares de Panicum maximum Jacq. Revista Brasileira de Zootecnia, v.29, n.2, p.341-348, 2000.
- GOMIDE, C.A.M.; GOMIDE, J.A.; ALEXANDRINO, E. Índices morfogênicos e de crescimento durante o estabelecimento e a rebrotação do capim-mombaça (P. Maximum Jacq.). Revista Brasileira de Zootecnia, n.4, p.795-803.
- HOAGLAND, D.A.; ARNON, D.I. The water culture method for growing plants without soil Berkeley: California Agric. Exp. Sta., 1950.39p. (Circular, 347).
- LARCHER, W. Physiological plant ecology 3.ed. Berlin: Springer-Verlag, 1995. 506p.
- LEMAIRE, G.; AGNUSDEI, M. Leaf tissue turnover and efficiency of herbage utilization. In: GRASSLAND AND ECOPHYSIOLOGY AND ECOLOGY, 1999, Curitiba. AnaisCuritiba: Universidade Federal do Paraná, 1999. p.165-186.
- MATTHEW, C.; KEMALL, W.D. Allocation of carbon14 to roots of different ages in perennial ryegrass ( Lolium perenne L.), In: GRASSLAND CONGRESS, 18., 1997, Manitoba/Saskachewan. Proceedings... Manitoba/Saskachewan: 1997. v.1, p.7.
- ROBSON, M.P.; RYLE, G.J.A.; WOLEDGE, J. The grass plant- its form and function. In: JONES, M.B.; LAZENBY, A. (Eds.) The grass crop London: Champman & Hall Ltda, 1988. p.25-83.
- RYLE, G.J.A. Distribution patterns of assimilated 14C in vegetative and reproductive shoots of Lolium perene and L.temulentum Annals of Applied Biology, v.66, n.97, p.155-167, 1970.
- RYLE, G.J.A.; POWELL, C.E. The export and distribution of 14C-labelled assimilates from each leaf on the shoot of Lolium temulentum during reproductive and vegetative growth. Annals of Botany, v.36, n.145, p.363-375, 1972.
- RYLE, G.J.A.; POWELL, C.E. The utilization of recently assimilated carbon in graminaceous plants. Annals of Applied Biology, v.77, n.2, p.145-158, 1974.
- RYLE, G.J.A.; POWELL, C.E. Effect of rate of photosynthesis on the pattern of assimilate distribution in the graminaceous plant. Journal of Experimental Botany, v.27, n.97, p.189-199, 1976.
- SCHNYDER, H.; DE VISSER, R. Fluxes of reserve-derived and currently assimilated carbon and nitrogen in perennial ryegrass recovering from defoliation. The regrowing tiller and its component functionally distinct zones. Plant Physiology, v.119, n.2, p.1423-1435, 1999.
- TAIZ, L.; ZEIGER, E. Translocation in the phloem. In: TAIZ, L.; ZEIGER, E. (Eds.) Plant physiology 2.ed. Sunderland: Sinauer Associates, 1998. p.251-259.
- WOLEDGE, J. The effect of age and shade on the photosynthesis of white clover leaves. Annals of Botany, v.57, n.2 , p.257-262, 1985.
- ZELITCH, I. The close relationship between net photosynthesis and crop yield. BioScience, v.32, n.10, p.796-802, 1982.
Datas de Publicação
-
Publicação nesta coleção
12 Set 2005 -
Data do Fascículo
Abr 2005
Histórico
-
Aceito
29 Dez 2004 -
Recebido
02 Mar 2004