Resumos
Os fotoassimilados produzidos pelas folhas são translocados por toda a planta, podendo variar conforme a fase de desenvolvimento, inserção da folha, desfolha, entre outros fatores. A distribuição dos fotoassimilados de folhas individuais para os diversos componentes da planta de Panicum maximum cv. Mombaça foi avaliada em função do nível de inserção da folha exposta ao 14CO2 (folha recém-expandida x folha no segundo nível de inserção abaixo), de diferentes perfilhos (perfilho principal, primeiro e terceiro perfilhos primários), de planta intacta ou submetida à remoção das lâminas foliares no nível da lígula, com exceção da folha exposta ao 14CO2. Apesar de os componentes estudados apresentarem quantidades distintas de 14C, significância para o efeito dos tratamentos foi constatada apenas para a porcentagem de 14C presente na parte aérea dos perfilhos, com valores mais altos correspondendo ao perfilho portador da folha exposta, independentemente da hierarquia do perfilho. A parte aérea reteve quantidade de 14C maior que o sistema radicular. A translocação de assimilados da folha da base tendeu a priorizar o conjunto de outros perfilhos, enquanto a folha do topo priorizou a translocação de assimilados para o próprio perfilho. De todos os componentes estudados, a folha do topo teve como principal dreno o meristema terminal, independentemente do tipo de perfilho portador da folha exposta e da condição de desfolhação.
14Carbono; partição de fotoassimilados; perfilhos; relação fonte; translocação
Leaf assimilates are exported to plant organs (sinks) in variable pattern according to several factors such as: plant developmental stage, leaf insertion level, intensity of defoliation. The distribution of single leaf assimilates to different fractions of Mombaçagrass plant was assessed as a function of several factors. The factors considered were: 1 Leaf insertion level (top and bottom leaves); 2 Tiller hierarchy (main and primary tillers), and 3 Plant defoliation (control and defoliated plants). Defoliated plants had their blades removed at the ligule level, with exception of the one exposed to 14 CO2. Significance of treatment effects was only observed for the percentage activity retained in plant aerial part, higher figures being found in the tiller of the exposed leaf, regardless of its hierarchy. The shoot retained more activity than the root system. Bottom leaves tended to favor translocation to new tillers, while the top leaf favored translocation to its own tiller. Considering all plant fractions, the top leaf translocated most to the terminal meristem, regardless of its tiller hierarchy and defoliation condition.
assimilates translocation; 14Carbon; source; tillers
FORRAGICULTURA
Distribuição dos fotoassimilados em plantas de Panicum maximum cv. Mombaça1 1 Parte da tese do primeiro autor; pesquisa financiada pela FAPEMIG.
Assimilates distribution in plants of Panicum maximum "cv" Mombaça
Emerson AlexandrinoI; José Alberto GomideII; Juraci Alves de OliveiraIII; Alice Cristina Bittencourt TeixeiraIV; Daniel Carlos Ferreira LanzaV
IProfessor Adjunto de Nutrição e Pastagem do Departamento de Agronomia, Universidade Federal de Rondônia, Campus de Rolim de Moura, Avenida Norte Sul, 7300, Bairro Nova Morada, Rolim de Moura, Rondônia, CEP: 78987-000
IIBolsista do CNPq
IIIProfessor Departamento de Biologia Geral, UFV
IVMestre em Zootecnia - UFV
VGraduando em Ciências Biológicas, UFV
RESUMO
Os fotoassimilados produzidos pelas folhas são translocados por toda a planta, podendo variar conforme a fase de desenvolvimento, inserção da folha, desfolha, entre outros fatores. A distribuição dos fotoassimilados de folhas individuais para os diversos componentes da planta de Panicum maximum cv. Mombaça foi avaliada em função do nível de inserção da folha exposta ao 14CO2 (folha recém-expandida x folha no segundo nível de inserção abaixo), de diferentes perfilhos (perfilho principal, primeiro e terceiro perfilhos primários), de planta intacta ou submetida à remoção das lâminas foliares no nível da lígula, com exceção da folha exposta ao 14CO2. Apesar de os componentes estudados apresentarem quantidades distintas de 14C, significância para o efeito dos tratamentos foi constatada apenas para a porcentagem de 14C presente na parte aérea dos perfilhos, com valores mais altos correspondendo ao perfilho portador da folha exposta, independentemente da hierarquia do perfilho. A parte aérea reteve quantidade de 14C maior que o sistema radicular. A translocação de assimilados da folha da base tendeu a priorizar o conjunto de outros perfilhos, enquanto a folha do topo priorizou a translocação de assimilados para o próprio perfilho. De todos os componentes estudados, a folha do topo teve como principal dreno o meristema terminal, independentemente do tipo de perfilho portador da folha exposta e da condição de desfolhação.
Palavras-chave:14Carbono, partição de fotoassimilados, perfilhos, relação fonte/dreno, translocação
ABSTRACT
Leaf assimilates are exported to plant organs (sinks) in variable pattern according to several factors such as: plant developmental stage, leaf insertion level, intensity of defoliation. The distribution of single leaf assimilates to different fractions of Mombaçagrass plant was assessed as a function of several factors. The factors considered were: 1 Leaf insertion level (top and bottom leaves); 2 Tiller hierarchy (main and primary tillers), and 3 Plant defoliation (control and defoliated plants). Defoliated plants had their blades removed at the ligule level, with exception of the one exposed to 14 CO2. Significance of treatment effects was only observed for the percentage activity retained in plant aerial part, higher figures being found in the tiller of the exposed leaf, regardless of its hierarchy. The shoot retained more activity than the root system. Bottom leaves tended to favor translocation to new tillers, while the top leaf favored translocation to its own tiller. Considering all plant fractions, the top leaf translocated most to the terminal meristem, regardless of its tiller hierarchy and defoliation condition.
Key Words: assimilates translocation, 14Carbon, source/sink relation, tillers
Introdução
Mais de 90% da massa seca vegetal se origina diretamente da assimilação fotossintética do carbono (Robson et al., 1988). Em plantas superiores, o mesófilo é o tecido mais ativo na assimilação de CO2, em razão da riqueza de suas células em cloroplastos (Taiz & Zeiger, 1998).
A translocação dos produtos da fotossíntese da fonte para os diversos drenos se faz pelo floema. Embora o modelo de transporte pelo floema represente simplesmente o movimento da fonte para o dreno, o caminho específico entre eles é altamente complexo, pois nem todos os drenos são igualmente supridos por todas as folhas da planta, uma vez que cada fonte supre drenos específicos. O modelo de distribuição dos assimilados é, em grande parte, função da proximidade entre os órgãos, as fontes e os drenos, do estádio de desenvolvimento dos órgãos drenos, das conexões vasculares e da alteração nos caminhos de translocação em que, na ausência de uma conexão direta entre fonte e dreno (ex. desfolha), as conexões vasculares (anastomoses) constituem um caminho alternativo (Taiz & Zeiger, 1998).
O meristema terminal (meristema apical + folhas em expansão + folhas emergentes), os perfilhos, o colmo, a raiz, a inflorescência e a formação de sementes nas plantas em estádio reprodutivo constituem-se nos diversos drenos das gramíneas. Os diversos drenos podem se tornar ativos e expandir para formar um órgão adulto ou permanecerem reprimidos, dependendo do genótipo, ontogenia e ambiente (Robson et al., 1988). A competição entre esses drenos constitui outro fator que pode determinar o modelo de translocação na planta: a força do dreno, que é dependente do seu tamanho e de sua atividade (Taiz & Zeiger, 1998).
A partição é definida como a distribuição diferencial de fotoassimilados pelos diferentes órgãos da planta e, portanto, determina o modelo de crescimento da planta, que pode ser balanceado entre parte aérea e sistema radicular. A pressão de turgescência no floema e os mensageiros químicos, como hormônios e nutrientes, são fatores que também controlam a relação fonte/dreno de assimilados (Taiz & Zeiger, 1998).
A taxa fotossintética da folha determina a quantidade total de carbono fixada disponível para a folha (Taiz & Zeiger, 1998) e é função de fatores intrínsecos, como o nível de inserção (Ryle & Powell, 1974) e a idade (Woledge, 1971), e de fatores extrínsecos às folhas, como a irradiância, a água no solo, a temperatura e os nutrientes (Larcher, 1995). Como resultado desse conjunto de fatores, as folhas de mais alto nível de inserção no perfilho, mais jovens e melhor iluminadas, fotossintetizam mais e geralmente contribuem mais para o pool de fotoassimilados da planta que as folhas mais velhas, de mais baixos níveis de inserção (Robson et al., 1988).
A partição de assimilados pelas diversas partes da planta pode ser estudada pelo incremento diferencial de biomassa destas partes, assim como pelas técnicas do carbono radioativo (14C), da auto-radiografia vegetal e da contagem da cintilação líquida.
Estudos auto-radiográficos em plantas de capim-timóteo (Phleum pratense L.) (Williams, 1964) evidenciaram que: 1 a folha emergente apenas importa assimilados; 2 as folhas adultas não importam assimilados, mas exportam, inicialmente, para o meristema apical, a folha emergente e o sistema radicular, de modo que, mais tarde, a exportação se faz predominantemente para a região inferior do perfilho; 3 gemas e perfilhos em desenvolvimento recebem assimilados de perfilhos adultos, que não importam de outros perfilhos. Entretanto, Marshal & Sagar (1968a), em estudos sobre a cintilação líquida em plantas de azevém anual (Lolium multiflorum Lam), demonstraram que, embora o assimilado de qualquer folha seja recuperado principalmente no próprio perfilho portador desta folha e no sistema radicular da planta, pequenas quantidades são também exportadas para outros perfilhos, independentemente da hierarquia do perfilho portador da folha em questão. Nos cultivares Mombaça e Tanzânia de Panicum maximum Jacq., a translocação de assimilados do perfilho principal para o perfilho primário mais jovem foi menor (6,5%) que do perfilho primário para o perfilho principal (14%), em razão da maior massa (Carvalho, 2002).
As folhas adultas retêm cerca de 10 a 25% de seus assimilados para sua manutenção, sendo o destino dos assimilados exportados determinado pela idade da folha e pelo seu nível de inserção no perfilho (Ryle, 1970a). Em plantas de azevém perene (Lolium perenne L.) no estádio de desenvolvimento vegetativo, o meristema terminal, o sistema radicular e os perfilhos recebem a maior parte dos assimilados da folha recém-expandida; posteriormente, o meristema terminal e as raízes se constituem nos principais drenos. Com a instalação do alongamento do colmo, a quantidade retida de assimilados nos perfilhos e nas raízes é pequena, mas o meristema terminal continua sendo importante dreno; na fase reprodutiva, o colmo se torna forte dreno e a exportação para os perfilhos e raízes é mínima (Ryle, 1970 b).
O carbono requerido para o desenvolvimento de um perfilho é importado principalmente de uma folha do perfilho em estádio hierarquicamente mais avançado. Geralmente, as folhas exportam grande proporção de assimilados para os perfilhos em crescimento, ou para um perfilho pequeno, próximo às suas axilas. O meristema intercalar do perfilho, quando se torna ativo, na maioria das vezes, utiliza os assimilados daquelas folhas que exportavam predominantemente para raízes e perfilhos (Ryle, 1970b; Ryle & Powell, 1972).
A independência do perfilho é estabelecida com o seu desenvolvimento. A habilidade para satisfazer sua necessidade de fotoassimilados ocorre quando o perfilho apresenta uma folha completamente expandida (Clifford et al., 1973). Contudo, a dependência nutricional dos perfilhos tende a se prolongar até a completa expansão da terceira folha, que coincide com a presença da formação do sistema radicular (Klepper et al., 1984).
A desfolha da planta pode alterar seu padrão de partição de assimilados. Segundo Ryle & Powell (1975), a exportação de assimilados de folhas mais baixas de cevada (Hordeum vulgare L), que, em plantas intactas, era predominantemente para o sistema radicular, foi revertida em favor do meristema terminal, imediatamente após desfolha que removeu as folhas mais novas da planta. Também, em plantas intactas de azevém anual, o principal dreno dos assimilados de suas folhas individuais era o sistema radicular, mas perfilhos cortados passaram a receber a maior parte dos assimilados, do perfilho intacto, imediatamente após o corte de todos, exceto de um dos perfilhos da planta (Marshal & Sagar, 1965). Igualmente, a desfolha de azevém anual incrementou a exportação de assimilados do perfilho principal para outros perfilhos (Marshall & Sagar, 1968b).
Qualquer área foliar residual, mesmo aquela que antes da desfolhação supria raízes e perfilhos, tende a exportar para o meristema terminal, por tempo variável, conforme a severidade da desfolhação (Robson et al., 1988). Este mecanismo de inversão da relação fonte/dreno explica o mais intenso perfilhamento e a produção de biomassa em plantas de festuca alta (Festuca arundinacea Schreb.) em rebrotação após desfolha parcial que resultou na permanência de alguns perfilhos intactos e/ou em maior área foliar residual (Matches,1966).
Portanto, a desfolhação é um dos fatores de manejo que interfere na partição de assimilados, cujo efeito no crescimento da planta durante o período de rebrotação é gradativo e dependente da intensidade de remoção da área foliar e da decapitação dos perfilhos. Essa interferência pode ser positiva ou negativa, conforme a habilidade da planta em restabelecer uma nova partição de assimilados das novas fontes para os novos drenos.
Em geral, após a desfolhação, os assimilados recém-sintetizados nas folhas residuais, bem como as reservas orgânicas, contribuem para a reconstituição da área foliar, priorizando os meristemas terminais (Schnyder & De Visser, 1999). Entretanto, a importância dos assimilados recém sintetizados vem ganhando crescente relevância, pois estudos recentes vêm demonstrando que, em perfilhos de azevém perene intensamente desfolhados, já no segundo dia, 63% da quantidade total do carbono presente nos novos tecidos são oriundos da fotossíntese (Schnyder & De Visser, 1999).
O conhecimento da partição e da alocação de assimilados pode subsidiar o manejo da cultura objetivando sua produtividade, ou seja, o aumento do índice de colheita (Beadle, 1993). Assim, o manejo da pastagem deve ser orientado em favor das folhas, em razão de seu papel na economia de carbono da planta e por constituir a fração mais nutritiva da dieta do ruminante.
Objetivou-se, com este experimento, obter informações sobre a partição e a dinâmica da relação fonte/dreno de fotoassimilados recém-sintetizados em plantas de Panicum maximum cv. Mombaça, conforme o nível de inserção da folha completamente expandida, de diferentes tipos de perfilhos e de plantas intactas ou submetidas à desfolhação.
Material e Métodos
O experimento foi conduzido em casa de vegetação da unidade de crescimento de plantas pertencente ao Departamento de Biologia Vegetal da UFV, desde a semeadura (25/10/2002) à colheita das plantas (20/12/2002).
Foram utilizadas plantas de Panicum maximum cv. Mombaça, obtidas de semeadura em bandejas perfuradas, contendo areia, irrigadas com água três vezes ao dia até a germinação, ocorrida ao quinto dia após semeadura (DAS).
No 20º DAS, as plântulas com uma folha emergente, duas folhas completamente expandidas e a cotiledonar senescente, foram transplantadas para vasos (três plântulas vaso-1) contendo 3,8 dm3 de substrato, constituído por uma mistura de solo, cujo pH foi elevado para 7 com o uso de calcário dolomítico, e areia, na proporção de 1:1. A esta mistura, foi adicionada dose única de superfosfato simples, equivalente a 400 mg.dm-3 de P. Aos 32º DAS, realizou-se o desbaste, deixando apenas uma planta por vaso, que constituiu a unidade experimental.
Após o transplantio, adicionaram-se, semanalmente, N e K, na dosagem de 80 e 70 mg.dm-3, respectivamente, e 200 mL.vaso-1 de solução de Hoagland & Arnon (1938), com força iônica total.
Foram avaliados o perfilho principal e o primeiro e terceiro perfilhos primários. O órgão fonte de fotoassimilados recém-sintetizados de cada um desses perfilhos foi apenas uma porção da lâmina foliar expandida, relacionada ao seu nível de inserção, em que a folha do topo corresponde àquela recém completamente expandida (mais jovem) do perfilho, e a da base, à que estava no segundo nível de inserção abaixo da folha do topo. A desfolhação, quando realizada, foi praticada pela remoção das lâminas foliares da planta no nível da lígula, minutos antes da exposição ao 14CO2, em todas as folhas da planta, inclusive a folha emergente, com exceção da lâmina foliar a ser exposta. A combinação desses três fatores gerou um total de 10 tratamentos (Tabela 1), visto que a folha da base do terceiro perfilho primário não foi exposta, pela dificuldade de seu manuseio para a exposição, decorrente de seu pequeno tamanho.
Os tratamentos foram dispostos em delineamento inteiramente casualizado, com três repetições, totalizando-se 30 unidades experimentais. As plantas encontravam-se em fase de perfilhamento no momento da exposição ao 14CO2.
Ao longo do desenvolvimento da planta, foi realizado o acompanhamento diário do perfilhamento em cada vaso, buscando-se identificar com anel colorido cada novo perfilho visualizado. Assim, foi possível a identificação do perfilho no momento da exposição da lâmina foliar ao 14CO2 e sua separação hierárquica no momento da colheita da planta.
Sacos plásticos transparentes foram utilizados como câmara para isolar a lâmina foliar a ser alimentada com o 14CO2 do restante da planta. Para isso, no momento de exposição, a folha previamente identificada foi introduzida no saco e, posteriormente, vedada com silicone e fita adesiva. Antes do isolamento da lâmina foliar, um pequeno tubo plástico, contendo bicarbonato de cálcio marcado (14C), e uma pipeta de plástico com ácido lático foram colocados no interior do saco para provocar a produção do gás marcado (14CO2) no interior da câmara.
Após o isolamento da folha, o vaso foi colocado sob uma estrutura de iluminação que irradiava sobre a planta um fluxo de fótons fotossintéticos em torno de 600 mmol.m-2.s-1. Uma lâmina d'agua corrente de ± 50 mm foi colocada entre a fonte de luz e a folha, para minimizar o efeito da temperatura das lâmpadas sobre a planta. Após posicionamento da folha sob a fonte luminosa, adicionou-se uma gota de ácido lático a 25 mL de solução do bicarbonato marcado (NaH14CO3), que apresentava atividade específica de 0,5 mCi.mL-1 de solução. Após a produção do 14CO2, o saco plástico foi agitado, a fim de se obter a distribuição do gás marcado por toda a câmara.
O tempo de exposição das lâminas foliares à atmosfera enriquecida com 14CO2 foi de 30 minutos. Decorrido esse período, por medida de segurança, colocou-se o vaso em capela, para a abertura dos sacos plásticos, realizada após o ar do interior do saco plástico ser succionado por alguns segundos e ser passado por uma série de frascos contendo solução de Ba(OH)2. Após retirada dos sacos plásticos, cada vaso foi levado para casa de vegetação, onde ficou sob condições naturais de luz, temperatura e umidade do ar.
Após 24 horas de exposição das lâminas ao 14CO2, as plantas foram individualmente colhidas e fracionadas nos seguintes componentes:
Perfilho principal e sua raiz;
Primeiro perfilho primário e sua raiz;
Segundo perfilho primário e sua raiz;
Terceiro perfilho primário e sua raiz;
Quarto perfilho primário; e
Perfilhos secundários (originados na gema axilar da folha de perfilhos primários).
Além disso, o perfilho cuja lâmina foliar foi exposta foi desmembrado em:
Meristema terminal (meristema apical + folhas em expansão e a folha emergente);
Folha adulta exposta ao 14CO2;
Demais folhas adultas, completamente expandidas verdes (bainha + lâmina ou apenas bainha das plantas desfolhadas); e
Base do perfilho (segmento entre a raiz e o meristema apical).
A colheita e separação dos componentes da planta de cada vaso durou aproximadamente 15 minutos. Nesse procedimento, cada perfilho foi desconectado da planta e o sistema radicular que o acompanhava foi considerado como a raiz desse perfilho.
Após separação e identificação, cada componente da planta foi levado à estufa sob ventilação forçada com temperatura de 100ºC por 20 minutos, para interromper o metabolismo vegetal. Findo este período, todo o material foi transferido para estufa a 70ºC por 72 horas, para posterior determinação da massa seca de cada componente. Cada componente foi, então, moído em almofariz elétrico, até obter-se textura equivalente a 200 mesh.
Alíquota de 10 mg de matéria seca de cada componente da planta foi adicionada em frascos de vidro de 20 mL para a contagem de cintilação líquida, e homogeneizada com 1 mL de Triton-X-100, por 15 segundos em vortex, adicionando-se, em seguida, 10 mL de coquetel de cintilação (30 g de cab-o-sil, 1 L de dioxano, 100 g de naftaleno e 5 g de PPO), procedendo-se a nova homogeneização.
Os frascos assim preparados foram colocados em espectrômetro de cintilação líquida Beckman 6500, o qual corrigia o quenching, automaticamente. Posteriormente, a radioatividade da alíquota foi convertida para a massa total de cada componente. A distribuição percentual da radioatividade de cada componente foi estimada a partir do somatório da atividade de todos os componentes da planta, tomado como 100% do carbono radioativo recuperado.
Os dados experimentais foram submetidos à análise de variância, adotando-se o teste Tukey, a 5,0%, para comparação das médias dos tratamentos.
Resultados e Discussão
Ao 14º dia após o transplantio, surgiu o primeiro perfilho primário na axila da terceira folha do perfilho principal. A partir de então, novos perfilhos surgiram a cada três ou quatro dias, em média. No momento de exposição de suas folhas ao 14CO2, o perfilho principal da gramínea apresentava em torno de 10 folhas completamente expandidas (4 a 6 senescentes) e cada planta possuía ± 6 perfilhos. Segundo Gomide & Gomide (2000), o capim-mombaça apresentou 8-9 perfilhos por planta ao 37º pós-emergência, com número de folhas verdes por perfilho principal estabilizado em 3-4 folhas. Neste estudo, a gramínea encontrava-se em pleno processo de perfilhamento e os perfilhos estudados apresentavam-se em diferentes estádios de desenvolvimento (Tabela 2) no momento de exposição de suas folhas ao 14CO2.
No momento da exposição das lâminas foliares ao 14CO2, os perfilhos estavam com 48, 19, 18, 16 e 12 dias e apresentavam massa seca da parte aérea de 4,1; 2,6, 3,2; 2,8 e 1,7 g, respectivamente, para o perfilho principal, o 1º, o 2º, o 3º e o 4º perfilhos primários. Entretanto, a massa seca do sistema radicular desses perfilhos primários foi inferior à do perfilho principal e ausente no quarto perfilho primário e no conjunto de perfilhos secundários (Figura 1).
A remoção das lâminas foliares antes da exposição da lâmina foliar ao 14CO2 representou grande redução na massa da parte aérea da gramínea (P<0,05), além de perdas menos expressivas do sistema radicular, nas 24 horas subseqüentes à desfolhação (Figura 1). A tendência de redução da massa seca do sistema radicular estaria relacionada à mobilização das reservas orgânicas.
A redução de massa seca dos perfilhos em decorrência da desfolhação é um fator de provável alteração no modelo de distribuição de fotoassimilados da planta, pois a massa do órgão (Taiz & Zeiger, 1998), juntamente com idade da folha (Rawson & Hofstra, 1969), posição e estádio fisiológico da folha fonte (Ryle, 1970a) e do tipo de perfilho (Marshall & Sagar, 1968a), determina o modelo de distribuição de fotoassimilados de determinada folha para os diversos drenos da planta.
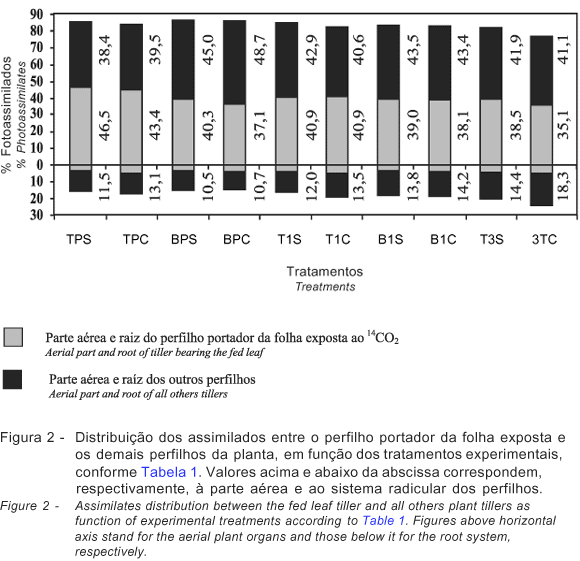
Os tratamentos experimentais, resultantes da combinação dos níveis de inserção da folha, da hierarquia dos perfilhos e da desfolhação, não tiveram efeito (P>0,05) sobre o modelo de distribuição percentual dos assimilados recuperados entre a parte aérea e o sistema radicular, seja do perfilho da folha exposta seja dos outros perfilhos da planta, após 24 horas de exposição (Figura 2).
A partição de 14C pela planta inteira alcançou valores médios de 82,5% para a parte aérea e 17,5% para o sistema radicular. Enquanto a porção referente à parte aérea se distribuiu praticamente em partes iguais para o perfilho cuja folha foi exposta (40%) e para os demais perfilhos (42,5%), maior fração foi exportada para o sistema radicular dos demais perfilhos (13,2%) que para o sistema radicular do perfilho portador da folha exposta (4,3%) (Figura 2). Todavia, considerando-se a incerteza quanto à correta identificação do sistema radicular dos diferentes perfilhos, é difícil atribuir significado maior à diferença entre raízes dos diferentes perfilhos. Nesta condição, vale considerar apenas a grande diferença na partição da parte aérea (82,5%) e o sistema radicular (17,5%).
Os valores de partição de assimilados para a raiz podem ser bem superiores aos encontrados nesta pesquisa. Valores em torno de 45,4 a 57,2% foram observados em Lolium multiflorum em fase de perfilhamento por Marshall & Sagar (1968a). Entretanto, geralmente o nível de inserção da folha e o estádio ontogênico da planta analisada interferem na partição de fotoassimilados da planta (Ryle, 1970b). Ryle (1970b) verificou que o percentual de translocação de assimilados para o sistema radicular variou de 50 a 9,5% em azevém perene (Lolim perenne), conforme seu estádio de desenvolvimento de apenas quatro e treze folhas, respectivamente, o que indica que os valores registrados neste estudo são condizentes com a fase de pleno perfilhamento do capim-mombaça, quando os novos perfilhos constituem-se no principal dreno.
Embora a massa do órgão seja um dos fatores determinantes de sua capacidade para importar assimilados (Taiz & Zeiger, 1998), ao se considerar os quatro componentes estudados (Figura 2), nota-se que a quantidade importada de assimilados não foi proporcional à sua massa, pois com massa percentual de 18; 52; 10 e 20%, perfilho da folha exposta, outros perfilhos, raiz do perfilho da folha exposta e raiz dos outros perfilhos, respectivamente, importaram 40, 42,5, 4,3 e 13,2% (Figura 2).
A distribuição de assimilados em função do nível de inserção da folha apresentou algumas peculiaridades, conforme o tipo de perfilho e a condição de desfolha da planta (Figura 2). Enquanto a folha do topo tendeu a translocar principalmente para o perfilho de origem conforme sua hierarquia, isto é, maior percentual correspondendo ao perfilho principal (45%) e menos para o primeiro (40,9%) e terceiro perfilhos primários (36,8%), a folha da base translocou preferencialmente para os demais perfilhos (46,9% na média do perfilho principal e 43,5% na média do 1º perfilho primário). Valores semelhantes foram obtidos em trigo (47%) e azevém perene (43%) por Ryle & Powell (1974).
Ressalta-se a tendência de mais alto percentual de exportação de assimilados para o sistema radicular pelas folhas dos perfilhos primários, sobretudo de suas folhas da base. Estas observações são consistentes com relatos de que folhas ontogenicamente mais velhas exportam preferencialmente para o sistema radicular. Maior quantidade 14C foi recuperada no sistema radicular de perfilhos mais jovens: perfilho principal 15,3%; 1º perfilho primário 17,7%; e 3º perfilho primário 21,6%, o que provavelmente esteja relacionado à prioridade para o desenvolvimento do sistema radicular daqueles perfilhos.
Tendência de substancial translocação de assimilados da folha do topo do 3º perfilho primário para o sistema radicular foi observada sob condições de desfolha da planta, demonstrando que o sistema radicular é um forte dreno em perfilhos novos (Figura 2). Essa constatação é, em princípio, contraditória aos relatos da literatura, que demonstram a prioridade da recuperação da parte aérea após a desfolhação, em que translocação de assimilados para o meristema terminal é aumentada em detrimento do sistema radicular (Ryle & Powell, 1975). Entretanto, esse comportamento aparentemente contraditório quanto à reconstituição da área foliar da planta após a desfolhação pode ser elucidado pelos resultados obtidos por Gifford & Marshall (1973), que observaram que a redução na translocação de assimilados para a raiz é decorrente da intensidade de desfolhação e que, em plantas de Lolium multiflorum, ela foi reduzida com a remoção tanto das lâminas como das bainhas foliares, de modo que a reconstituição da área foliar ocorreu via incremento na translocação de assimilados para os demais perfilhos. Neste experimento, foi observada apenas tendência de incremento na translocação de assimilados para os outros perfilhos, sob condição de desfolha do perfilho principal (Figura 2).
Em plantas que não perfilham (milho) ou perfilham pouco (sorgo), a translocação de assimilados da folha da base se faz preferencialmente para as raízes, enquanto, em plantas com intenso perfilhamento (trigo e azevém perene), a prioridade cabe aos perfilhos (Ryle & Powell, 1974). Neste estudo (Figura 2), a prioridade de translocação das folhas da base para os perfilhos foi condizente com o relato de Ryle & Powell (1974) quanto ao padrão observado em trigo e azevém. Isso decorre possivelmente da fase de desenvolvimento vegetativo do capim-mombaça, que, apresentando curto pseudo-colmo, propiciou igual proximidade dos diferentes drenos em relação à fonte de assimilados.
Os tratamentos experimentais não tiveram efeito (P>0,05) sobre o percentual de retenção de assimilados pelo sistema radicular dos diferentes tipos de perfilhos da planta. Entretanto, maior percentual de radioatividade na parte aérea correspondeu sempre ao perfilho portador da folha exposta, relativamente àquela encontrada na parte aérea de qualquer um dos demais perfilhos, independentemente de sua hierarquia (Figura 3). Porém, a somatória da radioatividade presente nos demais perfilhos correspondeu, aproximadamente, a 50% da radioatividade encontrada na parte aérea da planta.
O balanço entre retenção e exportação de assimilados foi praticamente nulo, independentemente do tratamento (Figura 3), o que está de acordo com Clifford et al. (1973), que verificaram independência por assimilados, entre perfilhos logo após a expansão de suas primeiras folhas.
Nota-se na Figura 3, tendência de variação do percentual de assimilados retidos na parte aérea do perfilho principal, conforme o nível de inserção da folha exposta e a condição de desfolha da planta. Observou-se tendência de valores mais altos no perfilho principal portador da folha exposta do topo de planta não desfolhada e valores mais baixos no perfilho principal portador da folha exposta da base da planta desfolhada.
Entretanto, houve tendência de perfilhos mais jovens exportarem maior quantidade de 14C para o sistema radicular.
O total de assimilados retidos na parte aérea do perfilho da folha exposta (Figura 3) equivale ao somatório do percentual de translocação de assimilados para cada um de seus quatro componentes: a base do perfilho (base + pseudo-colmo), as folhas expandidas (lâminas + bainhas ou apenas bainhas foliares quando a planta foi desfolhada), a folha exposta ao 14CO2 e o meristema terminal (meristema apical + folhas em expansão + folhas emergentes). A translocação de assimilados para cada um desses componentes é apresentada na Figura 4.
A exportação de assimilados para os diferentes órgãos da parte aérea do perfilho portador da folha exposta não variou (P>0,05) com os tratamentos experimentais (Figura 4), mas algumas tendências foram observadas. Assim, as folhas do topo tenderam a translocar mais assimilados para o meristema terminal que as folhas da base, principalmente para condição do perfilho principal, com valores de 15 e 10,8%, respectivamente (Figura 4).
Por outro lado, a quantidade de radioatividade retida na folha exposta ao 14CO2, que não variou (P>0,05) em razão dos tratamentos experimentais, demonstra tendência de maior retenção de 14C na folha do topo (Figura 4).
Observou-se percentual surpreendentemente alto de atividade nas demais folhas adultas ou em suas bainhas (respectivamente, 12 e 7,3%), sugerindo sua atuação como dreno (Figura 4). Essa constatação contradiz os relatos de Ryle (1970b) e Rawson & Hofstra (1969), que encontraram, respectivamente, valores de 2,0 e 1,0% em folhas adultas de azevém perene e trigo.
Independentemente do tratamento, a translocação de assimilados para a base (base da planta + pseudo-colmo) atingiu valores de 4,27 a 7,80% (Figura 4), sempre inferior àquela translocada para outros componentes do perfilho da folha exposta.
Avaliando-se individualmente todos os componentes da planta, conclui-se que o meristema terminal comportou-se como o dreno de maior força da planta de capim-mombaça quando a folha exposta foi a do topo, independentemente do tipo de perfilho ou da presença de desfolhação (Figuras 3 e 4).
Conclusões
A translocação de assimilados em plantas de capim-mombaça em fase de intenso perfilhamento foi mais acentuada para a parte aérea que para o sistema radicular.
Independentemente da hierarquia do perfilho, do nível de inserção da folha e da desfolhação, o perfilho portador da folha exposta ao 14CO2 reteve aproximadamente 50% dos assimilados encontrados na parte aérea da planta.
A exportação de assimilados para outros perfilhos tendeu ser mais acentuada quando da exposição da folha da base.
Alto percentual de assimilados foi transferido para o meristema terminal do perfilho portador da folha exposta, sobretudo quando esta era a folha recém-expandida.
Expressiva quantidade de assimilados foi recuperada em folhas adultas não expostas. Dos componentes do perfilho portador da folha exposta, a base apresentou o menor percentual de assimilados.
Literatura Citada
Recebido em: 29/06/04
Aceito em: 18/04/05
- BEADLE, C.L. Growth analysis. In: HALL, D.O.; BOLLARNORDENKAMPF, H.F.; LEEGOOD, R.C. et al. (Eds). Photosynthesis and production in a changing environment London: Pergamon Press, 1993. p.36-45.
- CARVALHO, D.D. Leaf morphogenesis and tillering behaviour in single plants and simulated sward of guineagrass (Panicum maximum Jacq.) cultivars New Zealand: Palmerston University, 2002. 186p. Thesis (PhD Plant Science) - Palmerston University, 2002.
- CLIFFORD, P.E.; MARSHALL, C.; SAGAR, G.R. The reciprocal transfer of radiocarbon between a developing tiller and its parent shoot in vegetative plants of Lolium multiflorum Lam. Annals of Botany, v.37, n.152, p.777-785, 1973.
- GIFFORD, R.M.; MARSHALL, C. Photosynthesis and assimilate distribution in Lolium multiflorum Lam. following differential tiller defoliation. Australian Journal of Biological Science, v.26, n.3, p.517-526, 1973.
- GOMIDE, C.A.M.; GOMIDE, J.A. Morfogênese de cultivares de Panicum maximum Revista Brasileira de Zootecnia, v.29, n.2, p.341-348, 2000.
- HOAGLAND, D.R.; ARNON, D.I. The method for growing plants without soil: water-culture. Berkeley: University of California, 1938. 32p. (Circular of the Agricultural Experimentation Station, 347).
- KLEPPER, B.; BELFORT, R.K.; RICKMAN, R.W. Root and shoot development in winter wheat. Agronomy Journal, v.76, n.1, p.117-122, 1984.
- LARCHER, W. Physiological plant eccology 3.ed. Berlin: Springer-Verlag, 1995. 506p.
- MARSHALL, C.; SAGAR, G.R. The interdependence of tillers in Lolim multiflorum: a quantitative assessment. Journal of Experimental Botanny, v.19, n.61, p.785-794, 1968a.
- MARSHALL, C.; SAGAR, G.R. The distribution of assimilates in Lolium multiflorum Lam. following differential defoliation. Annals of Botany, v.19, n.128, p.715-719, 1968b.
- MARSHALL, C.; SAGAR, G.R. The influence of defoliation on the distribution of assimilates in Lolium multiflorum Lam. Annals of Botany, v.29, n.115, p.365-372, 1965.
- MATCHES, A.G. Influence of intact tillers and height of stubble on growth responses of tall fescue. Crop Science, v.6, n.5, p.484-487, 1966.
- RAWSON, H.M.; HOFSTRA, G. Translocation and remobilization of 14C assimilated at different stages by each leaf of the wheat plant. Australian Journal Biological Science, v.22, n.2, p.321-331. 1969.
- ROBSON, M.J.; RYLE, G.J.A.; WOLEDGE, J. The grass plant its form and function. In: Jones, M.B.; Lazenby, A. (Eds.) The grass crop London: Chapman & Hall Ltda, 1988. p.25-83.
- RYLE, G.J.A.; POWELL, C. E. The export and distribution of 14C-labelled assimilates from each leaf on the shoot of Lolium temulentum during reproductive and vegetative growth. Annals of Botany, v.36, n.145, p.363-375, 1972.
- RYLE, G.J.A.; POWELL, C.E. The utilization of recently assimilated carbon in graminaceous plants. Annals of Applied Biology, v.77, n.2, p.145-158, 1974.
- RYLE, G.J.A.; POWELL, C.E. Defoliation and regrowth in the graminaceous plant: The role of current assimilate. Annals of Botany, v.39, n.160, p.297-310, 1975.
- RYLE, G.J.A. Distribution pattern of assimilated 14C in vegetative and reproductive shoots of Lolium perenne and L. temulentum Annals of Applied Biology, v.66, n.97, p.155-167, 1970b.
- RYLE, G.J.A. Partition of assimilates in an annual and a perennial grass. Journal of Applied Ecology, v.7, n.1, p.217-227, 1970a.
- SCHNYDER, H.; DE VISSER, R. Fluxes of reserve-derived and currently assimilated carbon and nitrogen in perennial ryegrass recovering from defoliation. The regrowing tiller and its component functionally distinct zones. Plant Physiology, v.119, n.2, p.1423-1435, 1999.
- TAIZ, L.; ZEIGER, E. Plant physiology 2.ed. Sunderland: Sinauer Associates, 1998. 792p.
- WILLIAMS, R.D. Assimilation and translocation in perennial grasses. Annals of Botany, v.28, n.111, p.419-427, 1964.
- WOLEDGE, J. The effect of light intensity during growth on the subsequent rate of photosynthesis of leaves of tall fescue (Festuca arundinacea Schreb). Annals of Botany, v.35, n.140, p.311-322, 1971.
Datas de Publicação
-
Publicação nesta coleção
01 Dez 2005 -
Data do Fascículo
Out 2005
Histórico
-
Aceito
18 Abr 2005 -
Recebido
29 Jun 2004