Resumos
Estudaram-se os efeitos da administração de enramicina e monensina sódica sobre a digestão total dos nutrientes da dieta e o consumo de matéria seca digestível em bovinos. Doze fêmeas bovinas não-gestantes e não-lactantes (675 ± 63 kg PV) foram distribuídas inteiramente ao acaso em três tratamentos (controle, enramicina e monensina) e alimentados com dieta contendo 60% de concentrados (milho, farelo de soja e minerais) e 40% de volumoso (cana-de-açúcar). A enramicina foi administrada na dose de 20 mg/animal/dia e a monensina na dose de 300 mg/animal/dia. O experimento teve duração total de 21 dias, de modo que os últimos dez dias foram utilizados para administração do marcador externo (15 g de óxido crômico/animal/dia) e os últimos cinco dias para a coleta de fezes e amostragem dos alimentos. Nenhum dos antibióticos alterou os consumos de matéria seca digestível e NDT e a digestibilidade de matéria seca, proteína bruta, extrato etéreo, fibra em detergente ácido, fibra em detergente neutro, amido, energia bruta e nutrientes digestíveis totais.
consumo; digestibilidade; ionóforos; não-ionóforos; ruminantes
The objective of this trial was to study the effects of enramycin and sodium monensin administration on total digestibility of diet nutrients and digestible dry matter intake in bovine. Twelve non-pregnant and non-lactating cows (675 kg ± 63 BW) were randomly assigned to three treatments: control group, enramycin-treated group or monensin-treated group. Animals received a diet containing 60% of concentrates (corn, soy bean meal and minerals) and 40% of forage (sugarcane). Treatments were 20 mg/animal/day of enramycin or 300 mg/animal/day of monensin. Trial lasted 21 days, the last 10 used for external marker administration (15 g of chromic oxide/animal/day) and the last 5 for feces collection and feed sampling. None of the antibiotics affected digestible dry matter and TDN intake and the digestibilities of dry matter, crude protein, ether extract, acid detergent fiber, neutral detergent fiber, starch, gross energy or total digestible nutrients.
digestibility; intake; ionophores; non-ionophores; ruminants
RUMINANTES
Efeitos da enramicina e monensina sódica sobre a digestão de nutrientes em bovinos alimentados com dietas contendo alto nível de concentrados
Effects of enramycin and sodium monensin on nutrients digestibility in bovine fed with high-concentrate diets
Luiz Felipe de Onofre BorgesI; Roberta PassiniII; Paula Marques MeyerIII; Paulo Henrique Mazza RodriguesIV, V
IMestrando do Departamento de Nutrição e Produção Animal - FMVZ/USP
IIUnidade de Ciências Exatas e Tecnológicas - UnUCET - Universidade Estadual de Goiás
IIIAnalista Agropecuário do Instituto Brasileiro de Geografia e Estatística IBGE
IVDepartamento de Nutrição e Produção Animal - FMVZ/USP, Av. Duque de Caxias Norte, 225 - CEP: 13630-000, Pirassununga - SP
VBolsista Produtividade em Pesquisa do CNPq
RESUMO
Estudaram-se os efeitos da administração de enramicina e monensina sódica sobre a digestão total dos nutrientes da dieta e o consumo de matéria seca digestível em bovinos. Doze fêmeas bovinas não-gestantes e não-lactantes (675 ± 63 kg PV) foram distribuídas inteiramente ao acaso em três tratamentos (controle, enramicina e monensina) e alimentados com dieta contendo 60% de concentrados (milho, farelo de soja e minerais) e 40% de volumoso (cana-de-açúcar). A enramicina foi administrada na dose de 20 mg/animal/dia e a monensina na dose de 300 mg/animal/dia. O experimento teve duração total de 21 dias, de modo que os últimos dez dias foram utilizados para administração do marcador externo (15 g de óxido crômico/animal/dia) e os últimos cinco dias para a coleta de fezes e amostragem dos alimentos. Nenhum dos antibióticos alterou os consumos de matéria seca digestível e NDT e a digestibilidade de matéria seca, proteína bruta, extrato etéreo, fibra em detergente ácido, fibra em detergente neutro, amido, energia bruta e nutrientes digestíveis totais.
Palavras-chave: consumo, digestibilidade, ionóforos, não-ionóforos, ruminantes
ABSTRACT
The objective of this trial was to study the effects of enramycin and sodium monensin administration on total digestibility of diet nutrients and digestible dry matter intake in bovine. Twelve non-pregnant and non-lactating cows (675 kg ± 63 BW) were randomly assigned to three treatments: control group, enramycin-treated group or monensin-treated group. Animals received a diet containing 60% of concentrates (corn, soy bean meal and minerals) and 40% of forage (sugarcane). Treatments were 20 mg/animal/day of enramycin or 300 mg/animal/day of monensin. Trial lasted 21 days, the last 10 used for external marker administration (15 g of chromic oxide/animal/day) and the last 5 for feces collection and feed sampling. None of the antibiotics affected digestible dry matter and TDN intake and the digestibilities of dry matter, crude protein, ether extract, acid detergent fiber, neutral detergent fiber, starch, gross energy or total digestible nutrients.
Key Words: digestibility, intake, ionophores, non-ionophores, ruminants
Introdução
Ionóforos são substâncias produzidas por várias cepas de Streptomyces sp., entre elas, monensina, lasalocida, salinomicina e narasina. Por definição, são moléculas de baixo peso molecular capazes de interagir estequiome-tricamente com íons metálicos servindo como transportadores para esses íons através de uma membrana lipídica biomolecular (Ovchinnikov, 1979).
Algumas dessas substâncias, como a monensina, aumentam a produção de ácido propiônico no rúmen, o que resulta em decréscimo da proporção de ácido acético, mas não altera significativamente a concentração total de ácidos graxos voláteis. Esse efeito é causado pela ação seletiva na população microbiana, que diminui o número de bactérias Gram-positivas (Kone et al., 1989). A resistência das bactérias Gram-negativas aos ionóforos parece ser conferida pela presença de uma camada peptideoglicana em sua membrana, impermeável a grandes moléculas (Russell & Strobel, 1989).
Segundo Schelling (1984), os modos de ação dos ionóforos são dois: um básico, que ocorre na membrana celular dos microrganismos ruminais; e outro sistêmico, composto de sete categorias de ação, resultantes da alteração do metabolismo das bactérias do rúmen e que afeta a resposta animal. Desse modo, os ionóforos podem aumentar a eficiência do metabolismo energético e protéico no rúmen e no animal, além de diminuir a incidência de distúrbios digestivos (Bergen & Bates, 1984); seu efeito sobre a digestibilidade ruminal pode ser influenciado por fatores como adaptação, dieta, estado fisiológico e idade dos animais, tempo de incubação, tipo e dose do produto utilizado, entre outros (Lucci et al., 2001). De acordo com Spears (1990), tanto em rações ricas em concentrados como naquelas ricas em volumoso, a monensina e a lasalocida têm aumentado a digestibilidade da fibra. Este aumento nem sempre é constante, pois autores como Pomar et al. (1989) constataram diminuição na digestibilidade da fibra em dietas predominantemente concentradas e aumento em dietas predominantemente volumosas. Outros pesquisadores, como McCann et al. (1990), observaram capacidade da monensina em aumentar a digestibilidade da fibra e da proteína à medida que a proporção de volumoso foi reduzida.
Van Der Merwe et al. (2001) sugerem também o uso de antibióticos não-ionóforos, como a avoparcina, flavomicina, tilosina, bacitracina e virginamicina, que promovem o crescimento do animal por meio de alterações nas características fermentativas do rúmen.
Novos produtos com potencial para modificar a fermentação ruminal têm sido desenvolvidos e necessitam ser testados. O produto testado neste experimento foi a enramicina, um antibiótico não-ionóforo promotor de crescimento muito usado em aves e suínos, produzido a partir da cepa de Streptomyces fungicidus e que possui estrutura polipeptídica (Kawakami & Nagai, 1971). Neste estudo, estudaram-se os efeitos da administração da enramicina ou da monensina sódica sobre a digestão total dos nutrientes da dieta e os consumos de MS digestível e de NDT em bovinos.
Material e Métodos
O experimento foi realizado nas dependências do Departamento de Nutrição e Produção Animal da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo (Campus de Pirassununga). Utilizaram-se 12 fêmeas bovinas mestiças, não-lactantes e não-gestantes, portadoras de cânulas ruminais e com 675 ± 63 kg de PV ao início do experimento.
O estábulo utilizado possuía baias individuais com cochos de cimento, bebedouros automáticos, piso emborrachado e ventiladores suspensos no teto, que eram ligados nas horas mais quentes do dia. O delineamento experimental foi o inteiramente casualizado com três tratamentos: controle negativo (ausência de antibiótico); enramicina; e monensina sódica. Para o tratamento com enramicina, utilizou-se o produto comercial Enradin F80 (Coopers Brasil Ltda.) na dose de 20 mg de enramicina/animal/dia, ou seja, 250 mg de Enradin F80/animal/dia ou 1,5 ppm de princípio ativo na MS da dieta. Para o tratamento com monensina, utilizou-se o produto comercial Rumensin (Elanco) na dose de 300 mg de monensina sódica/animal/dia, ou seja, 3,0 g de Rumensin/animal/dia ou 21,7 ppm de monensina sódica na MS da dieta, próximo da média de 11,0 a 33,0 ppm recomendada pela empresa que comercializa o produto. Cada produto foi pesado separadamente em balança analítica e acondicionado em envelopes confeccionados em papel absorvente, os quais foram administrados diariamente no interior do rúmen, através da fístula ruminal, e misturados ao conteúdo ruminal por meio de agitação manual. A dose diária foi dividida em duas vezes, correspondendo ao momento das refeições.
Os animais foram alimentados duas vezes ao dia, às 8 e 16 h, com dieta contendo 60% de concentrados e 40% de volumoso (Tabela 1), fornecida na forma de mistura completa, permitindo-se 10% de sobras.
O período experimental foi de 21 dias, os dez primeiros para adaptação dos animais às dietas e o restante para avaliação da digestibilidade in vivo. A digestibilidade da MS da dieta e suas frações, como proteína bruta (PB), extrato etéreo (EE), extrativos não-nitrogenados (ENN), fibra bruta (FB), energia bruta (EB), fibra em detergente neutro (FDN), fibra em detergente ácido (FDA) e amido, foi avaliada pela estimativa da produção fecal, obtida com uso de óxido crômico (Bateman, 1970). Os animais receberam o óxido crômico via cânula ruminal, na dosagem de 15 g/animal/dia, duas vezes ao dia (7,5 g de marcador/dose), no momento das refeições, por meio de envelopes confeccionados em papel absorvente. O ensaio foi dividido em duas fases, entre os dias 11 e 21, de modo que uma fase (cinco dias) foi destinada à administração do marcador e outra (cinco dias) à administração do marcador e à coleta de fezes. Para compor uma amostra final, procedeu-se à amostragem de alimentos (200 g de alimentos por coleta) e fezes (200 g/animal/coleta) duas vezes ao dia, próximo às refeições (8 e 16 h). As amostras de fezes foram coletadas diretamente do reto.
A concentração do óxido crômico foi determinada por colorimetria considerando sua reação com a s-difenilcarbazida, segundo Graner (1972). As análises bromatológicas de MS, PB, EE, FB, EB e cinzas foram realizadas segundo AOAC (1980) e as de FDN e FDA segundo Van Soest et al. (1991). Para a análise de FDN, foi omitido o sulfito de sódio, mas adicionada a a-amilase. A concentração de amido foi avaliada segundo Pereira & Rossi Jr. (1995) modificando-se esta metodologia com a prévia extração dos carboidratos solúveis, segundo Hendrix (1993).
Os resultados foram analisados por meio do programa computacional Statistical Analysis System (SAS, 1985) e submetidos à análise de variância (PROC GLM do SAS) e ao teste Tukey, a 5% de significância, para separação dos efeitos de tratamento. Adotou-se o nível de significância de 5%, exceto quando especificado.
Resultados e Discussão
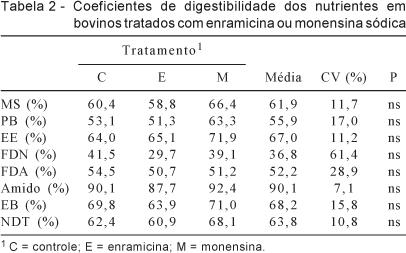
De modo geral, o valor de NDT da dieta (63,8%), independentemente do tratamento, foi um pouco menor que o esperado para esta dieta, que seria de 73,0% (Tabela 2). Essa diferença pode ser explicada pela taxa de recuperação do óxido crômico. Em ensaios anteriores (dados não publicados), observou-se taxa média de 86,0% de recuperação do marcador, portanto, os valores de NDT observados não são compatíveis com os esperados.
Nenhum dos antibióticos testados afetou a digestibili-dade de MS, PB, EE, ENN, FB, FDA, FDN, amido, energia bruta ou NDT. Morris et al. (1990), utilizando monensina, lasalocida ou tilosina, e Ricke et al. (1984), utilizando lasalocida ou monensina, também não observaram efeitos dos aditivos sobre a digestibilidade desses nutrientes, embora as condições experimentais não tenham sido exatamente as mesmas.
Os ionóforos podem causar pequena a moderada melhora na digestibilidade dos alimentos, dependendo das condições experimentais. Estas condições, não estão definidas até o momento, podem sofrer interferência de fatores como consumo de alimentos, enchimento ruminal ou taxa de passagem (Rodrigues et al., 2000).
Segundo McGuefey et al. (2001), a digestão da fibra não é afetada por ionóforos, o que foi claramente confirmado neste ensaio, no qual não foi possível comprovar diferença estatística entre o grupo controle e aqueles tratados com os produtos, o que pode ser conseqüência do aumento do número de bactérias resistentes aos antibióticos e de bactérias fibrolíticas, resultando em redução da quantidade de bactérias sensíveis aos produtos. De acordo com Lemenager et al. (1978), tempo maior de retenção de MS no rúmen, causado pelo ionóforo, pode contribuir para a manutenção da digestão normal da fibra. Entretanto, segundo Pomar et al. (1989), a monensina, quando administrada a dietas basicamente volumosas, causa aumento da digestibilidade da FDA e FDN. Nas dietas predominantemente concentradas, ocasiona diminuição da digestibilidade destas frações fibrosas. Thornton & Owens (1981) e Zinn et al. (1994) não observaram efeitos dos ionóforos sobre a digestibilidade da fibra, independentemente do nível desta fração na dieta. Ao adicionarem monensina ao ambiente ruminal não-adaptado ao produto, Henderson et al. (1981) observaram inibição quase total da digestão celulolítica in vitro ao utilizarem palha de cevada e fibras de algodão como substrato. Entretanto, utilizando a mesma concentração de monensina, a digestibilidade da celulose, também in vitro, foi bem menos inibida quando centeio e papel-filtro pulverizado foram utilizados como substrato. Esses resultados sugerem que propriedades químicas e/ou físicas associadas a diferentes fontes de fibra influenciam a digestibilidade da FDA e FDN em resposta aos ionóforos (Spears, 1990).
Os resultados com antibióticos não-ionóforos têm variado bastante. Em estudo realizado por Han et al. (2002), a digestibilidade aparente da matéria orgânica teve aumento apenas numérico com a adição de monensina + tilosina a uma dieta basicamente concentrada. Os efeitos desses produtos foram investigados in vivo por Zinn (1987), que também verificou aumento apenas numérico da digestibilidade da matéria orgânica. Haimoud et al. (1995) concluíram que a monensina + avoparcina diminuíram a degradação ruminal de fibra, amido e nitrogênio e aumentaram o suprimento de aminoácidos e de glicose para o intestino delgado. Alert et al. (1993) afirmaram, ao realizarem experimentos com touros em confinamento, que a suplementação com flavomicina aumentou a digestibilidade da matéria orgânica e do NDT e melhorou os parâmetros de fermentação ruminal.
A digestibilidade do amido também pode ser afetada pelo uso de ionóforos, no entanto, este fenômeno ainda não é bem explicado e não pôde ser confirmado neste estudo. Rogers & Davis (1982) demonstraram que a monensina administrada em novilhos alimentados com dieta mista (50% de volumoso) não alterou a digestibilidade de MS, matéria orgânica ou FDN, mas tendeu a aumentar a digestibilidade do amido em 2,7%, o que foi explicado pela provável diminuição no consumo de alimentos e pelo conseqüente aumento no tempo de retenção da MS no rúmen.
Haimoud et al. (1995) observaram aumento da quantidade de amido digerido no intestino delgado e também aumento da digestibilidade intestinal do amido não-degradável no rúmen causada pela monensina. Essa mudança na digestão do amido provavelmente resultaria em maior quantidade de carbono do amido sendo absorvida como glicose, em vez de AGV, o que tornaria mais eficiente o uso energético pelo animal hospedeiro. Funk et al. (1986) utilizaram lasalocida em ovinos e verificaram resultados semelhantes.
Huntington (1997) concluiu que esta maior quantidade de amido no intestino delgado não traz vantagens energéticas para os ruminantes, primeiramente pela capacidade limitada do intestino delgado em digerir amido e também pela utilização visceral de glicose resultante da digestão amilolítica no intestino delgado. Wedegaertner & Johnson (1983), no entanto, não observaram alterações na digestibilidade do amido em novilhos com dietas predominantemente concentradas, mas obtiveram aumentos de 3,5; 13,9 e 4,2%, respectivamente, na digestibilidade da MS, da FDN e da energia digestível.
Diversos estudos comprovaram que a monensina reduz a digestibilidade da proteína e dos aminoácidos livres no rúmen (Surber & Bowman, 1998). Os resultados obtidos neste estudo confirmam que não houve mudança significativa, o que está de acordo com o observado por Dinus et al. (1976), Johnson Jr. et al. (1988), Pomar et al. (1989), Marounek et al. (1990), Lana et al. (1997) e Rodrigues et al. (2001).
Peixoto Jr. et al. (2001) também não observaram alterações nos coeficientes de digestibilidade de PB, FB, EE e NDT ao utilizarem dieta mista e diferentes níveis de lasalocida. Knowlton et al. (1996), utilizando lasalocida, Haimoud et al. (1995), utilizando monensina, Morris et al. (1990), utilizando monensina, lasalocida ou tilosina, e Darden et al. (1985), utilizando lasalocida, monensina ou avaporcina, também não encontraram efeito dos aditivos sobre a digestibilidade desses nutrientes, embora tenham trabalhado com dietas e condições experimentais diferentes.
Outros autores descreveram aumento na digestibilidade aparente da proteína bruta com o emprego dos ionóforos na dieta (Ricke et al., 1984; Galloway et al., 1993). A teoria mais aceita nos dias de hoje foi citada por Chen & Russell (1991) e Van Kessel & Russell (1992), a qual sugere que os ionóforos podem diminuir a deaminação, fazendo com que os peptídeos e aminoácidos protegidos da deaminação possam ser convertidos em proteína microbiana por cepas resistentes aos ionóforos.
Observando a redução da concentração de nitrogênio amoniacal ruminal e da quebra protéica de origem dietética, Haimoud et al. (1995) concluíram que, com a utilização de monensina, mais proteína passaria intacta ao intestino delgado, sem afetar a síntese de proteína microbiana, aumentando em torno de 8% a digestibilidade total do nitrogênio. Do mesmo modo, Spears (1990) obteve resultados semelhantes, mas este aumento foi em torno de 3,5%.
Ives et al. (2002) verificaram que a virginamicina, assim como a monensina, possui efeito "economizador" de proteína, uma vez que ocasiona redução na deaminação de aminoácidos in vivo em animais alimentados com dietas basicamente concentradas, entretanto, estudos in vitro não deram suporte a essa teoria.
Nenhum dos antibióticos testados alterou os consumos de MS digestível e de NDT, expressos em quilos ou em porcentagem do peso vivo dos animais (Tabela 3). Os animais sob suplementação com monensina tenderam a apresentar maior consumo de MS digestível, expresso em porcentagem do peso vivo, em relação ao grupo controle (P<0,10), embora o consumo de MS digestível nos animais tratados com enramicina não tenha diferido dos demais grupos.
Os estudos com ionóforos têm sido realizados, em sua maioria, com gado de corte (Erasmus et al., 2005). Normalmente, estes produtos melhoram a eficiência da utilização de alimentos e, quando adicionados a dietas ricas em concentrados, a ingestão de MS diminui e a média de ganho de peso não é afetada (Nagajara et al., 1997). Resultados de experimentos com vacas leiteiras sugerem efeito nulo sobre a ingestão de MS (Johnson Jr. et al., 1988; Sauer et al., 1989; Phipps et al,. 2000; Plazier et al., 2000), enquanto, em outros estudos, foi observada diminuição do consumo (Sauer et al., 1998). Todavia, qualquer efeito nulo dos ionóforos na ingestão de MS, particularmente em vacas em início de lactação, é crítico para a produtividade e saúde do animal, uma vez que a ingestão de MS deve aumentar rapidamente (Erasmus et al., 2005).
Interação nível de fibra × monensina foi observada por Rodrigues et al. (2004). A quantidade de concentrado causou resposta curvilinear, com maior ingestão de MS quando oferecida a dieta mista. A monensina proporcionou resposta linear, diminuindo a ingestão de MS de acordo com o aumento na dose.
Resultados semelhantes foram obtidos por Salles & Lucci (2000) com diferentes níveis de suplementação de monensina e uma dieta predominantemente concentrada para bezerros. Esses autores constataram comportamento quadrático para o consumo de MS, de modo que a maior ingestão ocorreu quando fornecida monensina (0,8 mg/kg de PV, estimado em 14,3 mg/kg de MS ingerida) e a diminuição da ingestão ocorreu no nível mais alto desse ionóforo (1,2 mg/kg de PV, estimado em 22,7 mg/kg de MS ingerida).
Rodrigues et al. (2001) forneceram monensina a ovinos alimentados com diferentes proporções volumoso:concentrado e não observaram diferenças estatísticas causadas pela monensina sobre o consumo de MS na dieta predominantemente concentrada ou volumosa, no entanto, o fornecimento desse aditivo diminuiu (36,7%) o consumo na dieta mista. Vargas et al. (2001), ao fornecerem monensina a bovinos, observaram redução no consumo de MS proporcional ao aumento do nível de concentrado na dieta.
Os resultados obtidos nesta pesquisa estão de acordo com os de vários outros, nos quais não houve efeitos dos ionóforos sobre o consumo de alimento em diversas condições (Dinus et al., 1976; Ricke et al., 1984; Branine & Galyean, 1990; Galloway et al., 1993; Ramazin et al., 1997; Green et al., 1999; Rodrigues et al., 2000; Plazier et al., 2000; Broderick, 2004; Osborne et al., 2004). Garcia et al. (2000), fornecendo monensina a cabras alimentadas com dieta composta de 50% de concentrado e 50% de volumoso (feno de alfafa), também não constataram mudanças no consumo de MS.
Conclusões
Em bovinos alimentados com dietas predominantemente concentradas, com cana-de-açúcar como único volumoso, não foi possível demonstrar os efeitos benéficos da monensina ou da enramicina, nos níveis utilizados, sobre a digestibilidade dos nutrientes e os consumos de matéria seca digestível e de nutrientes digestíveis totais.
Agradecimento
Aos funcionários Everson Lázaro e Gilmar Botteon, pelo cuidado com os animais. Aos técnicos Ari de Castro, Gilson de Godoy, Simi Aflalo e Isabel Ramos, pela ajuda nas análises laboratoriais.
Literatura Citada
Este artigo foi recebido em 8/5/2006 e aprovado em 17/9/2007.
Correspondências devem ser enviadas para pmazza@usp.br.
- ALERT, H.J.; POPPE, S.; LOHNER, M. The effect of flavomycin on the fattening performance of bulls. Arch Tierernahr, v.43, n.4, p.371-380, 1993.
- ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS - AOAC. Official methods of analysis10.ed. Washington, D.C.: 1980. 1015p.
- BATEMAN, J. Nutricion animal. In: Manual de métodos analíticos México: Herrero Hermanos, 1970. p.405-449.
- BERGEN, W.G.; BATES, D.B. Ionophores: their effect on production efficiency and mode of action. Journal of Animal Science, v.58, n.6, p.1465-1483, 1984.
- BRANINE, M.E.; GALYEAN, M.L. Influence of grain and monensin suplementation on ruminal fermentation, intake, digesta kinetics and incidence and severity of frothy bloat in steers grazing winter wheat pasture. Journal of Animal Science, v.68, n.3, p.1139-1150, 1990.
- BRODERICK, G.A. Effect of low level monensin supplementation on the production of dairy cows fed alfalfa silage. Journal of Dairy Science, v.87, n.2, p.359-368, 2004.
- CHEN, G.; RUSSELL, J.B. Effect of monensin and photonophore on protein degradation peptide accumulation, and deamination by mixed ruminal microoganisms in vitro Journal of Animal Science, v.69, n.5, p.2196-2203, 1991.
- DARDEN, D.E.; MERCHEN, N.R.; BERGER, L.L. et al. Effects of avoparcin, lasalocid, and monensin on sites of nutrient digestion in beef steers. Nutrition Reports International, v.31, n.4, p.979-989, 1985.
- DINUS, D.A.; SIMPSON, M.S.; MARSH, P.B. Effect of monensin with forage and digestion and the ruminal ecossystem of steers. Journal of Animal Science, v.42, n.1, p.229-234, 1976.
- ERASMUS, L.J.; ROBINSON, P.H.; AHMADI, A. et al. Influence of prepartum and postpartum supplementation of a yeast culture and monensin, or both, on ruminal fermentation and performance of multiparous dairy cows. Animal Feed Science and Technology, v.122, n.3, p.219-239, 2005.
- FUNK, M.A.; GALYEAN, M.L.; ROSS, T.T. Potassium and lasalocid effects on performance and digention in lambs. Journal of Animal Science, v.63, n.3, p.685-691, 1986.
- GALLOWAY, D.L.; GOETSCH, A.L.; PATIL, A. et al. Feed intake and digestion by Holtein steers calves consuming low-quality grass suplemented with lasalocid and monensin. Canadian Journal of Animal Science, v.73, n.4, p.869-879, 1993.
- GARCIA, C.C.G.; MENDOZA, M.G.D.; GONZÁLEZ, M.S. et al. Effect of a yeast culture (Saccharomyces cerevisiae) and monensin on ruminal fermentation and digestion in sheep. Animal Feed Science and Technology, v.83, n.2, p.165-170, 2000.
- GRANER, C.A.F. Determinação do crômio pelo método colorimétrico da s-difenilcarbazida Botucatu: Universidade Estadual Paulista, 1972. 112p. Dissertação (Mestrado em Zootecnia) - Universidade Estadual Paulista, 1972.
- GREEN, B.L.; McBRIDE, B.W.; SANDALS, D. et al. The impact of a monensin controlled-release capsule on subclinical ketosis in the transition dairy cow. Journal of Dairy Science, v.82, n.2, p.333-342, 1999.
- HAIMOUD, D.A.; VERNAY, M.; BAYOURTHE, C. et al. Avoparcin and monensin effects on the digestion of nutrients in dairy cows fed a mixed diet. Canadian Journal of Animal Science, v.75, n.2, p.379-385, 1995.
- HAN, H.; HUSSEIN, H.S.; GLIMP, H.A. et al. Carbohydrate fermentation and nitrogen metabolism of a finishing beef diet by ruminal microbesin continuous cultures as affected by ethoxyquin and (or) supplementation of monensin and tylosin. Journal of Animal Science, v.80, n.4, p.1117-1123, 2002.
- HENDERSON, C.; STEWART, C.S.; NEKPEK, F.V. The effect of monensin on pure and mixed cultures of rumen bacteria. Journal of Applied Bacteriology, v.51, n.1, p.159-169, 1981.
- HENDRIX, D.L. Rapid extraction and analysis of nonstructural carbohydrates in plant tissues. Crop Science, v.33, n.6, p.1306-1311, 1993.
- HUNTINGTON, G.B. Starch utilization by ruminants: from basics to the bank. Journal of Animal Science, v.75, n.3, p.852-867, 1997.
- IVES, S.E.; TITGEMEYER, E.C.; NAGAJARA, T.G. et al. Effects of virginiamycin and monensin plus tylosin on ruminal protein metabolism in steers fed corn-based finishing diets with or without wet corn gluten feed. Journal of Animal Science, v.80, n.11, p.3005-3015, 2002.
- JOHNSON JR., J.C.; UTLEY, P.R.; MULLINIX JR., B.G. et al. Effects of adding fat and lasalocid to diets of dairy cows. Journal of Dairy Science, v.71, n.8, p.2151-2165, 1988.
- KAWAKAMI, M.; NAGAI, Y. Anti-microbial activities of Enduracin (Enramycin) in vitro and in vivo.The Journal of Antibiotcs, v.24, n.9, p.583-586, 1971.
- KNOWLTON, K.F.; ALLEN, M.S.; ERICKSON, P.S. Lasalocid and particle size of corn for dairy cows in early lactation. 2. Effect on ruminal measurements and feeding behavior. Journal of Dairy Science, v.79, n.4, p.565-574, 1996.
- KONE, P.; MACHADO, P.F.; COOK, R.M. Effect of combination of monensin and isoacids on rumen fermentation in vitro Journal of Dairy Science, v.72, n.10, p.2767-2771, 1989.
- LANA, R.P.; FOX, D.G.; RUSSELL, J.B. et al. Influence of monensin on Holstein steers fed high-concentrate diets containing soybean meal or urea. Journal of Animal Science, v.75, n.6, p.2571-2579, 1997.
- LEMENAGER, R.P.F.N.; OWENS, B.J.; SHOCKEY, B.J. et al. Monensin effects on rumen turnover rate, twenty four hour VFA pattern, nitrogen components and celulose disapperance. Journal of Animal Science, v.47, n.1, p.255-261, 1978.
- LUCCI, C.S.; PEIXOTO, K.C.J.; AMARO, F.R. et al. Efeitos de níveis e de tempos de adaptação a lasalocida sódica sobre a degradabilidade ruminal do feno de coast-cross (Cynodon dactylon) e do farelo de soja em bovinos. Boletim da Indústria Animal, v.58, n.1, p.95-112, 2001.
- MAROUNEK, M.; PETR, O.; MACHAÑOVÁ, L. Effect of monensin on in vitro fermentation of maize starch by hindgut contents of cattle. Journal of Agricultural Science, v.115, n.3, p.389-392, 1990.
- McCANN, M.A.; CRADDOCH, B.F.; PRESTON, R.L. et al. Digestibility of cotton plant by-product diets for sheep at two levels of intake. Journal of Animal Science, v.68, n.2, p.285-295, 1990.
- McGUEFEY, R.K.; RICHADSON, L.F.; WILKINSON, J.I.D. Ionophores for dairy cattle: currents status and future outlook. Journal of Animal Science, v.84, n.1, p.194-203, 2001.
- MORRIS, F.E.; BRANINE, M.E.; GALUEN, M.L. et al. Effects of rotating monensin plus tylosin and lasalocid on performance, ruminal fermentation, and site and extent of digestions in feedlot cattle. Journal of Animal Science, v.68, n.10, p.3069-3078, 1990.
- NATIONAL RESEARCH COUNCIL - NRC. Nutrients requirements for dairy cattle 6.ed. Washington, D.C.: 1989. 157p.
- NAGAJARA, T.G.; NEWBOLD, C.J.; Van NEVEL, C.J. et al. Manipulation of ruminal fermetation. In: HOBSON, P.N.; STEWART, C.S. (Eds.) The rumen microbial ecossystem 2.ed. New York: Blackie, 1997. p.523-562.
- OSBORNE, J.K.; MUTSVANGWA, T.; ALZAHAL, O. et al. Effects of monensin on ruminal forage degradability and total tract diet digestibility in lactating dairy cows during grain-induced subacute ruminal acidosis. Journal of Dairy Science, v.87, n.6, p.1840-1847, 2004.
- OVCHINNIKOV, J.A. Physico chemical basis of ion transport through biological membranes: ionophores and ion channels. European Journal of Biochemistry, v.94, n.2, p.321-336, 1979.
- PEIXOTO JR., K.C.; LUCCI, C.S.; AMARO, F.R. et al. Efeitos de níveis e de tempos de adaptação à lasalocida sódica sobre a digestibilidade aparente da dieta total em bovinos. Boletim da Indústria Animal, v.58, n.1, p.113-123, 2001.
- PEREIRA, J.R.A.; ROSSI JR., P. Manual prático de avaliação nutricional de alimentos 1.ed. Piracicaba: Fundação de Estudos Agrários Luiz de Queiroz, 1995. 25p.
- PHIPPS, R.H.; WILKINSON, J.I.D.; JONKERT, J.I.D. et al. A. Effect of monensin on milk prodution of Holstein-Friesian dairy cows. Journal of Dairy Science, v.83, n.12, p.2789-2794, 2000.
- PLAZIER, J.C.; MARTIN, A.; DUFFIED, T. et al. B.W. Effect of a prepartum administration of monensin in a controlled-release capsule on apparent digestibilities and nitrogen utilization in transition dairy cows. Journal of Dairy Science, v.83, n.12, p.2918-2925, 2000.
- POMAR, C.; BERNIER, J.F.; SEOANE, F.R. et al. High-roughage rations with or without monensin for veal production. 2. Ration digestibility. Canadian Journal of Animal Science, v.69, n.2, p.403-410, 1989.
- RAMAZIN, M.; BAILONI, L.; SCHIAVON, S. et al. Effects of monensin on milk production and efficiency of dairy cows fed two diets differing in forage to concentrate ratios. Journal of Dairy Science, v.80, n.8, p.1136-1142, 1997.
- RICKE, S.C.; BERGER, L.L.; Van der HAAR, P.J. et al. Effects of lasalocid and monensin on nutrient digestion, metabolism and rumen characteristics of sheep. Journal of Animal Science, v.58, n.1, p.194-202, 1984.
- RODRIGUES, P.H.M.; LUCCI, C.S.; CASTRO, A.L. Efeitos da lasalocida sódica e proporção volumoso/concentrado sobre a degradabilidade in situ do farelo de soja e do feno Coast Cross [Cynodon dactylon (L) Pers] em vacas secas. Brazilian Journal of Veterinary Research and Animal Science, v.37, n.3, p.253-258, 2000.
- RODRIGUES, P.H.M.; MATTOS, W.R.S.; MELOTTI, L. et al. Monensina e digestibilidade aparente em ovinos alimentados com proporções de volumoso e concentrado. Scientia Agrícola, v.58, n.3, p.449-455, 2001.
- RODRIGUES, P.H.M.; MATTOS, W.R.S.; MEYER, P.M. et al. Effects of monensin level and roughage/concentrate ratio on ruminal fermentation in bovines. Journal of Animal and Feed Science, v,13, p.195-198, 2004 (suppl. 1).
- ROGERS, J.A.; DAVIS, C.L. Rumen volatile fatty acid production and nutrient utilization in steers fed a diet supplemented with sodium bicarbonate and monensin. Journal of Dairy Science, v.65, n.6, p.944-952, 1982.
- RUSSELL, J.B.; STROBEL, H.J. Minireview: effect of ionophores on ruminal fermentation. Applied and Environemental Microbiology, v.55, n.1, p.1-6, 1989.
- SALLES, M.S.V.; LUCCI, C.S. Monensina para bezerros ruminantes em crescimento acelerado. Revista Brasileira de Zootecnia, v.29, n.2, p.573-581, 2000.
- STATISTICAL ANALYSIS SYSTEM - SAS. SAS user's guide: statistics. 5.ed. Cary: SAS Institute, 1985. 441p.
- SAUER, F.D.; KRAMER, J.K.G.; CANTWELL, W.J. Antiketogenic effect of monensin in the early lactancy. Journal of Dairy Science, v.72, n.2, p.436-442, 1989.
- SAUER, F.D.; FELLNER, V.; KINSMAN, R. et al. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added to the diet. Journal of Animal Science, v.76, n.3, p.906-914, 1998.
- SCHELLING, G.T. Monensin mode of action in the rumen. Journal of Animal Science, v.58, n.6, p.1518-1527, 1984.
- SPEARS, J.W. Ionophores and nutrient digestion and absorption in ruminants. Journal of Nutrition, v.120, n.6, p.632-638, 1990.
- SURBER, L.M.M.; BOWMAN, J.G.P. Monensin effects on digestion of corn or barley high-concentrate diets. Journal of Animal Science, v.76, n.7, p.1945-1954, 1998.
- THORNTON, J.H.; OWENS, F.N. Monensin supplementation and in vivo methane production by steers. Journal of Animal Science, v.52, n.3, p.628-634, 1981.
- Van DER MERWE, B.J.; DUGMORE, T.J.; WALSH, K.P. The effect of flavophospholipol (Flavomycin) on milk prodution and milk urea nitrogen concentration of grazing dairy cows. South Africa Journal of Animal Science, v.31, n.2, p.101-105, 2001.
- Van KESSEL, J.S.; RUSSELL, J.B. Energetics of arginine and lysine transport by whole cells and membrane vesicles of strains SR, a monensin-sensitive ruminal bacterium. Applied and Environmental Microbiology, v.58, n.3, p.969-975, 1992.
- Van SOEST, P.J.; ROBERTSON, J.B.; LEWIS, B.A. Symposium: Carboydrate metodoloy, metabolism, and nutritional implications in dairy cattle. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science, v.74, n.10, p.3583-3597, 1991.
- VARGAS, L.H.; LANA, R.P.; MÂNCIO, A.B. et al. Influência de Rumensin®, óleo de soja e níveis de concentrado sobre o consumo e os parâmetros fermentativos ruminais em bovinos. Revista Brasileira de Zootecnia, v.30, n.5, p.1650-1658, 2001.
- WEDEGAERTNER, T.C.; JOHNSON, D.E. Monensin effects on digestion, methanogenesis and heat increment of a cracked corn-silage diet fed to steers. Journal of Animal Science, v.57, n.1, p.168-177, 1983.
- ZINN, R.A. Influence of lasalocid and monensin plus tylosin on comparative feeding value of steam-flaked versus dry-rolled corn in diets for feedlot cattle. Journal of Animal Science, v.65, n.1, p.256-266, 1987.
- ZINN, R.A.; PLASCENCIA, A.; BARAJAS, R. Interation of forage level and monensin in diets for feedlot cattle on growth performance and digestive function. Journal of Animal Science, v.72, n.9, p.2209-2215, 1994.
Datas de Publicação
-
Publicação nesta coleção
03 Abr 2008 -
Data do Fascículo
Abr 2008
Histórico
-
Recebido
08 Maio 2006 -
Aceito
17 Set 2007