Abstracts
The recent technologies that have led to the new field of functional genomics (how the genome of an organism regulates homeostasis and responds to stimuli) are providing a clearer understanding of how organisms interact with their environment and in particular their diet. We are beginning to learn how the diet may have long-term influence on performance and health. A form of epigenetic regulation has been recently described called fetal "programming". Fueled by epidemiological data the "fetal origins" hypothesis suggests that a poor in utero environment resulting from maternal dietary or placental insufficiency may "program" susceptibility in the fetus to cardiovascular or metabolic disorders. We have observed similar apparent programming by dietary manipulation in the chicken. When birds are challenged with a diet low in phosphorus (P) for 90 hours post-hatch they obtain the ability to better utilize P later in life. This increased retention of P from the diet can partially be explained by an enduring increase in the expression of the intestine-specific Na/P cotransporter (NaPcoT) gene during programming as well as later in life when fed P restricted diets. The resulting data provide the first evidence for neonatal programming of gene expression in an oviparous species.
epigenetics; functional genoimcs; nutrition; nutritional conditioning; poultry
As tecnologias recentes que levaram ao novo campo da genômica funcional (como o genoma de um organismo regula a homeostase e responde aos estímulos) estão proporcionando uma compreensão mais clara de como os organismos interagem com seu ambiente e em especial a sua dieta. Nós estamos começando a aprender como a dieta pode ter influência a longo prazo sobre o desempenho e a saúde. Uma forma de regulação epigenética foi descrita recentemente chamada de "programação fetal". Alimentados por dados epidemiológicos da "origem fetal" hipótese sugere que um pobre no útero ambiente decorrentes da insuficiência alimentar ou placentária materna pode programar "suscetibilidade do feto de doenças cardiovasculares ou metabólicas. Tem-se observado programação aparente similar por manipulação dietética da galinha. Quando as aves são desafiadas com uma dieta baixa em fósforo (P) por 90 horas pós-eclosão, elas obtêm a capacidade de utilizar P melhor mais tarde na vida. Esta maior retenção de P da dieta pode ser parcialmente explicada por um aumento permanente na expressão do gene do cotransoprtador Na/P intestinal durante a programação, bem como mais tarde na vida quando alimentadas com dietas P restrito. Os dados obtidos fornecem a primeira evidência para a programação neonatal da expressão gênica em uma espécie ovípara.
aves; condicionamento nutricional; epigenética; genômica funcional; nutrição
NONRUMINANTS
Christopher M. AshwellI; Roselina AngelII
IDepartment of Poultry Science, North Carolina State University Campus Box 7608, Raleigh, NC 27695. Phone: 919-513-7335, Fax: 919-515-2625
IIDepartment of Animal and Avian Sciences, University of Maryland College Park, MD
ABSTRACT
The recent technologies that have led to the new field of functional genomics (how the genome of an organism regulates homeostasis and responds to stimuli) are providing a clearer understanding of how organisms interact with their environment and in particular their diet. We are beginning to learn how the diet may have long-term influence on performance and health. A form of epigenetic regulation has been recently described called fetal "programming". Fueled by epidemiological data the "fetal origins" hypothesis suggests that a poor in utero environment resulting from maternal dietary or placental insufficiency may "program" susceptibility in the fetus to cardiovascular or metabolic disorders. We have observed similar apparent programming by dietary manipulation in the chicken. When birds are challenged with a diet low in phosphorus (P) for 90 hours post-hatch they obtain the ability to better utilize P later in life. This increased retention of P from the diet can partially be explained by an enduring increase in the expression of the intestine-specific Na/P cotransporter (NaPcoT) gene during programming as well as later in life when fed P restricted diets. The resulting data provide the first evidence for neonatal programming of gene expression in an oviparous species.
Key Words: epigenetics, functional genoimcs, nutrition, nutritional conditioning, poultry
RESUMO
As tecnologias recentes que levaram ao novo campo da genômica funcional (como o genoma de um organismo regula a homeostase e responde aos estímulos) estão proporcionando uma compreensão mais clara de como os organismos interagem com seu ambiente e em especial a sua dieta. Nós estamos começando a aprender como a dieta pode ter influência a longo prazo sobre o desempenho e a saúde. Uma forma de regulação epigenética foi descrita recentemente chamada de "programação fetal". Alimentados por dados epidemiológicos da "origem fetal" hipótese sugere que um pobre no útero ambiente decorrentes da insuficiência alimentar ou placentária materna pode programar "suscetibilidade do feto de doenças cardiovasculares ou metabólicas. Tem-se observado programação aparente similar por manipulação dietética da galinha. Quando as aves são desafiadas com uma dieta baixa em fósforo (P) por 90 horas pós-eclosão, elas obtêm a capacidade de utilizar P melhor mais tarde na vida. Esta maior retenção de P da dieta pode ser parcialmente explicada por um aumento permanente na expressão do gene do cotransoprtador Na/P intestinal durante a programação, bem como mais tarde na vida quando alimentadas com dietas P restrito. Os dados obtidos fornecem a primeira evidência para a programação neonatal da expressão gênica em uma espécie ovípara.
Palavras-chave: aves, condicionamento nutricional, epigenética, genômica funcional, nutrição
Introduction
In the current scientific era of the genome the ability to completely decipher the entire genetic code for an organism has provided great insight into the organization of genes and chromosome structure. But simply sequencing a genome does not provide insight into how this information is used in the animal and how it is regulated to perform all functions within an organism. The field of functional genomics has attempted to address the regulation of the genome by characterizing the expression patterns of genes both in homeostasis as well as perturbed situations (ie. stress, disease, drug treatment). It is when the link between genetics and gene expression (or phenotype) does not behave as expected where a more cryptic element comes into play, referred to as epigenetics. This code which overlays the genome sequence can manipulate the expression of genes by both chemical and physical modification of the genome. The role of epigenetics has become a hot bed of activity in the human arena and spawned a post-genome sequencing effort to form the International Human Epigenome Project (IHEP) and characterize how the epigenome differs from tissue to tissue and how it responds to environmental changes (temperature, stress, disease, ect). The impact of epigenetics is most profoundly observed in twin studies where individuals containing exact genome copies have significantly different gene expression patterns often resulting in differing susceptibilities to disease (Fraga et al., 2005).
Imprinting
One of the fundamental assumptions of Mendelian genetics is that a specific allele behaves in the same way no matter from where (which parent) it originates. But as with many assumptions in science this rule does not always hold true. Initial observations in mice looking for the effects of paternal or maternal disomy using genomic translocations indicated that identical chromosomal regions were not equivalent and acted differently in embryo development between maternal and paternal sources (Cattanach & Kirk, 1985). Although the terminology surrounding this phenomenon has been referred to as gametic imprinting the interpretation is described as "an allele-specific reversible epigenetic modification dependent on the parent of origin allele" (Ruvinski, 1999). It was later hypothesized that the reason for gametic imprinting in viviparous species was to ease the conflict during pregnancy between the maternally and paternally inherited genes. Imprinting in mammals is thought to have evolved as a result of the fetus being directly nourished by maternal tissues. The paternal genes stimulating growth of the fetus while the maternal genes restricting growth so that the mother may successfully deliver the offspring. Imprinting can therefore be thought of as a compromise between the maternal and paternal alleles or a compromise between the mother and the fetus. The most well characterized mechanism for the silencing of a single allele of a gene is by methylation of CpG islands in the regulatory regions of imprinted genes such as insulin-like growth factor 2 and mannose 6-phosphate/insulin-like growth factor 2 receptor (Rappolee et al., 1992; Wang et al., 1994; Lau et al., 1994). CpG islands are short, dispersed regions of unmethylated DNA with a high frequency of CpG dinucleotides relative to the bulk genome. DNA with a high frequency of CpG dinucleotides relative to the bulk genome. Other mechanisms including histone modification and chromatin structural modifications can also be responsible for epigenetic regulation of gene expression (Razin, 1998).
DNA methylation
DNA methylation is a naturally occurring event that happens in both prokaryotic and eukaryotic organisms. In prokaryotes DNA methylation provides a way to protect host DNA from digestion by their own restriction enzymes that are designed to eliminate foreign DNA. In higher eukaryotes DNA methylation acts as another level of gene regulation. DNA methylation has also been shown to play a central role in gene imprinting, embryonic development, X-Chromosome gene silencing and cell cycle regulation. Early analysis of 5-methyl-cytosine of several organisms showed a non-random distribution across the genome. This DNA methylation pattern was thought to be correlated with gene expression (Razin & Riggs, 1980). Methylation of DNA primarily occurs within symmetrical dinucleotide CG, or CpG clusters, often referred to as CpG "islands" which are located in the promoter regions of genes (Maeda and Hayashizaki, 2006). The majority of the DNA methylation in mammals is found in 5'-CpG-3' dinucleotides, but other methylation patterns do exist. In fact, about 80 percent of all 5'-CpG-3' dinucleotides in mammalian genomes are methylated, and the majority of the 20 percent that remain unmethylated are found within promoters or in the first exons of genes. Normally a C (cytosine) base followed immediately by a G (guanine) base (a CpG) is rare in vertebrate DNA because the Cs in such an arrangement tend to be methylated. This methylation helps distinguish the newly synthesized DNA strand from the parent strand, which aids in the final stages of DNA proofreading after duplication. However, over evolutionary time cytosine tends to mutate into thiamine (T) because of spontaneous deamination (Razin, 1998). The result is that CpGs are relatively rare unless there is selective pressure to keep them or a region is not methylated for some reason, perhaps having to do with the regulation of gene expression.
The pattern of DNA methylation in these CpG islands varies during mouse cell differentiation and is also dependent on cell lineage (Chapman et al., 1984). Later work provided evidence that the degree of 5-methyl-cytosine present was directly proportional to gene expression where high DNA methylation is equivalent to gene silencing (Li et al., 1993). It has been clearly demonstrated that aberrant methylation is a widespread phenomenon in cancer and may be among the earliest changes during oncogenesis (Jiang et al., 2004).
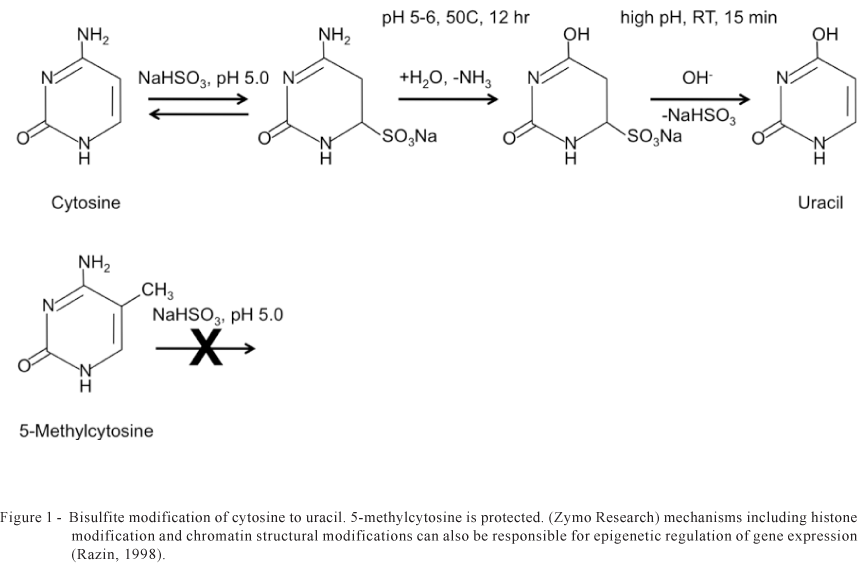
It is obvious that the ability to quantify and detect DNA methylation efficiently and accurately is essential for the study the epigenetic basis of cancer, gene expression, genetic diseases, and many other important aspects of biology. To date, a number of methods have been developed to quantify DNA methylation such as high performance capillary electrophoresis and methylation-sensitive arbitrarily primed PCR, but currently the most commonly used technique is the bisulfite method. This technique consists of treating DNA with bisulfite, which causes unmethylated cytosines to be converted into uracil while methylated cytosines remain unchanged (Figure 1). In this approach the bisulfite modified DNA is amplified by PCR and the resulting PCR products are either analyzed by DNA sequencing or restriction endonuclease digestion. The methylation status of the DNA segment is then determined by comparing the bisulfite treated DNA to the untreated DNA.
Early work looking at the methylation status of chicken genes using methyl-sensitive and -insensitive restriction enzymes for tissue-specific differences in methylation were found for the delta-crystallin, beta-tubulin, G3PDH, rDNA, and actin genes but not for the histone genes (Cooper et al, 1983). Genomic imprinting is hypothesized to not occur in oviparous taxa such as birds by a parent of origin mechanism. Examination of the allelic expression of mannose 6-phosphate/insulin-like growth factor 2 receptor (M6P/IGF2R) and insulin-like growth factor 2 (IGF2), imprinted in mammals showed that neither M6P/IGF2R nor IGF2 are imprinted in the chicken (Nolan et al., 2001).
Fetal programming
The growing incidence of metabolic diseases such as obesity, diabetes, and cardio-vascular disease has sparked interest and research into both their genetic and environmental (nutritional) basis. It has long been known that the maternal diet, and therefore the nutrient supply to the developing oocyte, embryo or fetus, is one of the principal environmental factors influencing the development of the offspring. A reliable and balanced supply of amino acids, lipids and carbohydrates is required to support the high rates of cell proliferation and the key developmental processes that take place during the embryonic (pre-implantation) and fetal (post-implantation) stages of life. Eukaryotic cells have evolved a complex series of nutrient sensors that are able to regulate gene expression in response to imbalances in the supply of nutrients. In adults these systems serve two purposes; firstly to protect the cell from damage caused by acute deficiencies and secondly to optimize homeostatic control to deal with a prolonged excess or deficiency of a particular nutrient. This second process may have a critical impact on the long term health of the offspring. It has been proposed that adverse nutritional conditions during fetal development lead to adaptive changes in metabolism that lead to a 'thrifty phenotype' in the offspring (Hales & Barker, 1992). Poor nutrition in early life produces permanent changes in glucose-insulin metabolism, including a reduced capacity for insulin secretion and insulin resistance (Hales & Barker, 2001). However, if this 'programming' of metabolism during embryonic and fetal development is inappropriate for the long-term nutritional environment it may lead to adverse long term consequences for the offspring (Sayer et al., 2004; Yajnik, 2004; Barker, 2004). The initiating factor(s) for fetal programming may be nutrient(s) interacting directly with genes and their regulatory elements at the cellular level, altering patterns of growth and gene expression. In this review we will concentrate on some of the direct interactions between genes and nutrients and their possible influence on fetal development.
A number of disease states in humans are the result of subtle changes in gene expression caused by alterations in chromatin structure (Robertson & Wolffe, 2000). Epigenetic variants of DNA methylation in CpG islands are set in the chromatin during development and determine the accessibility of a particular gene to the transcriptional machinery (Spiegelman & Heinrich, 2004). By regulating gene expression through changes in the promoter region, these epigenetic modifications represent a second level mechanism for the nutritional regulation of gene expression. There is a growing body of evidence showing that transcriptional repression and genome defense is perturbed by changes in metabolism. Embryo culture techniques in farm animals frequently expose the preimplantation embryo to an inappropriate nutritional environment in vitro, leading to defective epigenetic programming and a number of developmental abnormalities collectively known as the large offspring syndrome (Young et al., 2001). Studies have revealed that the expression of a number of key genes is altered in these large offspring (Young & Beaujean, 2004). Defects in epigenetic programming can also affect metabolism, the surviving offspring of mice produced by nuclear transfer (where the transferred nucleus undergoes extensive epigenetic reprogramming) exhibit an obese phenotype (Tamashiro et al., 2002). Many of the cytosine residues modified by methylation reside within parasitic DNA elements or retrotransposons, such as endogenous retroviruses.
It is becoming apparent that embryonic and fetal cells have a complex system to integrate nutritional signals from their environment and adapt their development accordingly to ensure survival. Human diets are comprised of complex mixtures of protein, fats, carbohydrate and vitamins. The full impact of inappropriate programming of metabolic regulation is only just beginning to be appreciated. The available evidence suggests that nutrient sensing regulatory systems are present in many critical tissues during early development. It remains to be seen whether they play an important part in establishing homeostatic control mechanisms early in life.
Thermotolerance in chickens
Similar observations to those found in placental organisms have been made in the chicken where conditioning in early-life imparts long-term effects. The first report of this type of response was to temperature or thermal stress. The basis for these studies was to identify a mechanism to impart tolerance to acute heat stress in chickens produced in sub-tropical climates. It was found that excessive thermal input during the first week of a chicken's life modulated the response to thermal stress later in life (Yahav & McMurtry, 2001). By simply increasing the brooding temperature from 30°C to 37.5°C for 24 hours within the first 5 d post-hatch birds are able to tolerate 6 hr of exposure to 35°C at 42 days of age, while "unconditioned" birds are unable to acclimate. The mechanism for this conditioned response is unknown. Several studies speculate that it is during the period immediately post-hatch (neonatal) that the chick is developing the connections in the brain for the detection and regulation of body temperature (Katz & Meiri, 2006; Labunsky & Meiri, 2006). Brain-derived neurotrophic factor (BDNF) has been implicated in the development of thermal regulation in the hypothalamus by thermal conditioning stimulating its expression, whereas knocking down expression in the same time frame causes aberrant response to thermal stress later in life. The expression of BDNF was not measured during the thermal stress challenge later in life; therefore long-term effects on its expression by thermal conditioning are not known. This innate ability to adapt by early-life conditioning prompted our studies that follow of what we than called early-life dietary adaptation.
Early dietary adaptation
Adaptation to low nutrient diets has been long recognized. Animals respond to nutrient restriction by increasing absorption rates and utilization efficiency, which decreases excretion of the restricted nutrients. The ability of humans to adapt to a diet low in Ca was recognized in the1950s. At that time, the Food and Nutrition Board (1948) recommended an adult daily Ca allowance of 800 mg/d. However, Hegsted et al. (1952) found that adult Peruvians, who had lived on low Ca diets for long periods, only required 100 to 200 mg Ca/d to maintain balance. It is obvious that these Peruvians, who grew up under Ca restriction, were able to better utilize Ca.
Adaptation to P and Ca restricted diets has also been previously reported in chickens. In an in-vitro trial, using ligated duodenal loops, Morrissey & Wasserman (1971) observed that broiler chicks absorbed a higher percent of a labeled 47Ca (ranging from 70 to 90%) when diets low in Ca (0.08%) were fed for eight d prior to intestinal sampling regardless of dietary P levels, or when low P (0.25%) diets were fed regardless of dietary Ca levels. Chickens receiving a diet with normal P (0.65%) and normal Ca (1.20%) absorbed less than 50% of 47Ca. Duodenal P absorption in 15 to 20 d-old chicks that had been fed a low Ca or a low P diet for eight d, as measured by ligated duodenal loop technique in vivo, increased by 49 and 87%, respectively (Fox et al., 1981). Blahos et al. (1987) reported an increase in duodenal and ileal P absorption in broiler chickens fed a low Ca diet for two wk and a smaller, but still significant increase in duodenal but not ileal P absorption in chicks fed a low P diet. The adaptation to P or Ca restriction was believed to be a result of an increased level of circulating 1,25-(OH)2-D3 (Blahos et al., 1987) and duodenal calbindin content (Morrisey & Wasserman, 1971; Montecuccoli et al., 1977). By comparing the duodenal calbindin concentration and its changing pattern with age for 1991 and 2001 strains of broilers, Bar et al. (2003) concluded that modern broilers exhibit higher capacity of adaptation to P or Ca deficiency and this capacity remains high for the whole growth period. However, no literature could be found on work conducted to evaluate the long-term effects of early P or Ca restriction on growth performance, bone mineralization, and P absorption in poultry. Therefore we proceeded to investigate if birds had the capacity to adapt similarly. The application of the adaptation principle in poultry may allow for decreasing both fed and excreted P and Ca without sacrificing performance and provide an additional low cost tool to decrease P and Ca in poultry litter. Understanding the changes associated with the increase in absorptive capacity in birds that have adapted to low P and Ca diets will be the first step in determining the viability of this method. We evaluated the ability of the chicken to adapt to a moderate early life deficiency in P and Ca and characterized this adaptation changes by examining the impact of the previous P and Ca status (starter phase, hatch to 18 d) on performance, bone characteristics, and nutrients absorption of broilers the grower phase (19 to 32 d) (Yan et al., 2005). In summary, broilers fed a diet moderately deficient in P and Ca from hatch to 18 d demonstrated ability to adapt to the deficiency, which was shown in the increased total P and Ca ileal absorption (Table 1), the increased PP disappearance, compensatory growth, and compensatory improvement in bone parameters including tibia ash, tibia and shank bone mineral density and bone mineral content in a later growth phase (18 to 32 d). However, practical application of the adaptation rule requires more studies to further fine tune the degree, timing, and length of the restriction with the aim of achieving minimal changes in bone, associated with no changes in performance, since bone characteristics are one of the primary determinant of downgrades of poultry products in the processing plants. These published data indicate that in birds during the period immediately post hatch there is a phenomenon occurring that permanently alters the bird's response to its environment. This adaptation or conditioning, whichever term you choose to use, is an observable fact for which no underlying mechanism has been previously proposed.
Focus on phosphorus
Land application of poultry litter, that is comparatively high in P due to the poor utilization of phytin P (PP) (Nelson, 1976) by poultry, is of increasing concern in areas of intensive poultry production in the United States (Sharpley, 1999). The Environmental Protection Agency (EPA) passed federal regulations that limits the amount of poultry litter that can be applied to soils, based mainly on litter P content (EPA, 2003). To address the P issue, poultry nutritionists have developed several nutritional strategies, that include feeding diets with P concentrations closer to requirements (Yan et al., 2001; Dhandu & Angel, 2003), dietary supplementation with feed additives such as microbial phytase (Simons et al., 1990; Denbow et al., 1995; Angel et al., 2005) and vitamin D3 metabolites (Edwards Jr., 1993; Biehl & Baker, 1997; Edwards Jr., 2002; Angel et al., 2005), and use of genetically modified feed ingredients with lower concentrations of PP (Cromwell et al., 1998; Waldroup et al., 2000; Li et al., 2000). Most of these strategies, although affective, can increase the cost of production. Alternative strategies, which can lower costs through epigenetic regulation of genes, will be of significant economic importance to the poultry industry.
Early P restriction - Performance data
To determine the effects of diet P on performance and expression of the chicken intestinal NaPcoT, experimental diets were formulated to be deficient in total P (Angel & Ashwell, 2008). Ross 308 chicks were fed either a control diet (C) consisting of 1.11% Ca and 0.50% available P (NRC levels) or a restricted diet (L) containing 0.59% Ca and 0.25% available P from hatch to 4 d of age (90hr). All birds were then fed a control diet (C) consisting of NRC recommended levels of Ca and P until d 22. From day 22 to d 38 the birds were either maintained on a C diet at NRC levels of Ca and P at 0.7% and 0.3% respectively or a restricted diet (L) consisting of 0.4% Ca and 0.12% P. The three dietary treatments, C-C-C, C-C-L, and L-C-L met all other NRC (1994) nutrient recommendations. Performance data were collected for each dietary phase including weight gain, feed conversion, bone ash, and specific nutrient retention. These data are presented in Table 1. All test diets and dried ileal contents were ground to pass 0.50 mm screen, then analyzed, in duplicate, for dry matter (AOAC), P, Ca, phytate P (PP) (Rounds & Nielsen, 1993), as modified by Newkirk & Classen (1998), and acid insoluble ash (Vogtmann et al., 1975). The non-PP concentrations for the diets were determined by subtracting analyzed PP from analyzed P. Apparent ileal absorption of P and Ca, and disappearance of PP were calculated based on acid insoluble ash analysis where Celite® was used as an indigestible marker.
Broilers fed the moderately deficient diet (L) to 90 hr were better able to handle a deficiency in P in the grower/finisher phase (22 to 38 d of age than those fed a control diet in the first 90 hr). Not only were the broilers fed the L diet early on heavier at 38 days of age, but they were more efficient in converting feed to gain, had high tibia ash and higher P retention than those fed the C diet in the first 90 hr of life. This clearly establishes that "imprinting" or modifications are occurring in the animal that are long term and that allow for improved P utilization when P deficient diets are fed in the grower/finisher phases.
Gene expression of NaPcoT
Birds were euthanized by cervical dislocation at 90 hr, d 22, and 38 to collect tissues for gene expression analysis (Ashwell & Angel, 2008). The duodenum, jejunum, and ileum were collected and total RNA extracted using TRIzol® Reagent (Invitrogen) according to the manufacturer's instructions, and resuspended in diethyl-pyrocarbonate treated water. Concentration and quality of the RNAs were determined by measuring the absorbance at 260nm and agarose gel electrophoresis, respectively. For consistency of comparison across individuals the anatomical location of samples collected were maintained across birds by taking tissue for analysis from a specific site in the intestine which was divided into the 3 regions using the following landmarks: duodenum (from distal of the gizzard to 1 cm distal of the bile duct); jejunum (1 cm distal from the bile duct to the Meckel's diverticulum); ileum (Meckel's diverticulum to 5 cm proximal to the ileocecal junction). The duodenum section was collected as a 1 cm segment in the duodenal loop. The jejunum sample was collected as a 1 cm section at the center of the jejunal segment. The ileum sample was collected as a 1 cm section at 5 cm distal to the Meckel's diverticulum.
Gene expression measurements for the chicken NaPcoT were determined using real-time quantitative PCR. Primers for the NaPcoT mRNA were designed using Beacon DesignerTM software (Premiere Biosoft) for SYBR green detection. Real-time PCR was carried out using the BioRad iQ instrument and with the iScript and iQ-SYBR green Supermix kits using the manufacturers protocols (BioRad). Individual cDNAs were diluted 1:20 prior to amplification. Thermalcycling parameters were as follows: 94°C for 5 min, 50 cycles of (94°C for 30 sec, 58°C for 30sec, 72°C for 30 sec), 72°C for 8 minutes, 4°C forever. Fluorescence measurements were collected at every cycle during the extension step (72°C). The primers used included NaPcoT-FOR 5'-CTGGATGCACTCCCTAGAGC-3', NaPcoT-REV 5'-TTATCTTTGGCACCCTCCTG-3', ch18s-FOR 5'-CCGAGAGGGAGCCTGAGAA-3', and ch18s-REV 5'-CGCCAGCTCGATCCCAAGA-3'. Each gene was amplified independently in triplicate within a single instrument run. Standard curves were also run to determine the efficiency of amplification by pooling undiluted cDNA from the duodenum samples across both treatments and diluting the pooled cDNA to dilutions of 1:5, 1:20, 1:100, and 1:500. Gene expression was normalized for RNA loading using 18s rRNA as an internal control and differences in gene expression were determined using the ΔΔCt method of Pfaffl (2001) including a correction for amplification efficiencies.
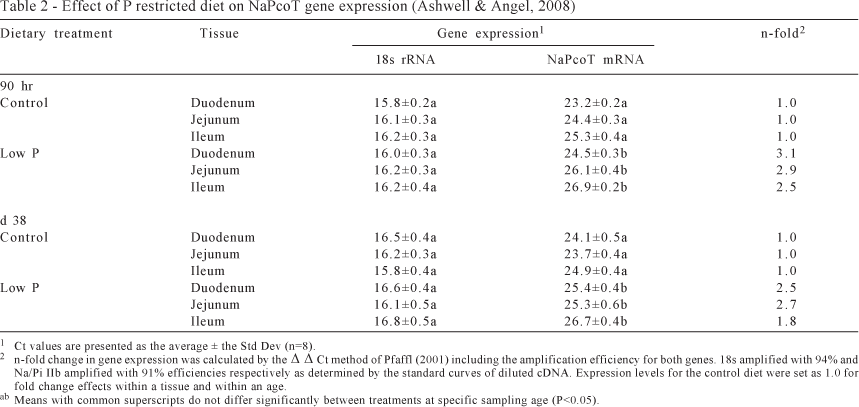
The response of the chicken intestinal NaPcoT to variation in substrate levels (dietary P) was determined by feeding newly hatched chicks a diet deficient in available P followed by the measurement of the gene expression level of the cotransporter mRNA using real-time PCR. RNA extracted from each of the regions of the small intestine were reverse transcribed to produce cDNA and analyzed by real-time PCR for the levels of both the NaPcoT mRNA and 18s rRNA. The effect of dietary treatment was determined by analyzing the resulting Ct values for each amplification and determining the ΔΔCt using the level of 18s rRNA as an internal standard for normalizing the amount of RNA in each reaction. Detection of the amplification by real-time PCR was by SYBR green I fluorescence. Each cDNA was analyzed for the levels of each of the transcripts in triplicate. The average Ct values for each of the tissues and treatments are shown in Table 1. The differences in Ct values for gene expression were determined to be significantly different using a student's t-test and the relative difference in expression caused by the dietary treatment was determined by correcting for the efficiency of the PCR as calculated by the standard curves of dilutions of pooled cDNA across the experiment. N-fold expression changes are also reported in Table 2.
The reduction of P in the post hatch diet had a significant effect on the expression of the NaPcoT by stimulating an average 2.8-fold increase in the mRNA levels in the small intestine. Nearly identical stimulatory effects were seen across all segments of the intestine with relative expression decreasing from duodenum to ileum. This pattern is similar to that in trout where a 2 fold induction of the NaPcoT was observed as a result of a 40% reduction of diet P (Sugiura et al., 2003). This is also in the range of the induction of expression of the NaPcoT as a result of feeding a low P diet in mice (Segawa et al., 2004). The effect of reducing P from a control diet level of 0.5% P to a low P diet level of 0.25% P induced NaPcoT expression an average of 2.3 fold. This change in expression was shown not to be involved in the vitamin D signaling pathway since knockout mice showed similar response to that of wildtype mice. The influence of vitamin D on the absorption of Pi (inorganic P) has long been known but further evidence has shown that 25-hydroxyvitamin-D3-1α-hydroxylase which leads to an increase in the level of 1,25-dihydroxy-vitamin D3 does not influence the expression of the NaPcoT in the intestine. This was demonstrated also through the use of knockout mice (Capuano et al., 2005). Therefore the regulation of expression of the NaPcoT by dietary P must function through a novel pathway that is not influenced by vitamin D or its metabolites.
DNA methylation - NaPcoT promoter
Upon investigation of the genome sequence upstream of the NaPcoT several CpG islands (5) are predicted. Identification of CpG islands is based on relative concentration of CpGs present in a specific sequence. The location of the islands is predicted by computer algorithm (Gardiner-Garden & Frommer,1987). A sliding-window search is performed looking for GC content of 50% 0r more over a length of 200 bp or more where the ratio of observed to expected CpGs is 0.6 or greater. Using the UC Santa Cruz genome browser (genome.ucsc.edu) one can query a specific gene (genome region) and view the CpG annotations in the display window. Bisulfite sequencing was employed to determine if there was evidence of differential DNA methylation in the CpG islands flanking the NaPcoT as a result of nutritional neonatal programming. Genomic DNA was extracted from the duodenum of both control and programmed birds (90h P restriction) (n=8 per treatment). A portion of this DNA was subjected to bisulfite modification using the DNA Methylation Gold kit from Zymo Research.
Primers targeting the CpG islands located in region 3 were designed the MethPrimer search tool (http://www.urogene.org/methprimer/) (Li & Dahiya, 2002). This region was initially selected due to its larger size and number of CpGs (74) that could be screened in a single assay. Design of the primers must take into account the bisulfite treatment, thus limiting the primer design parameters. PCR was conducted on both bisulfite treated and untreated genomics DNA. The resulting PCR products were sequenced in both the forward and reverse direction using BigDye Terminators (version 1.1) and an ABI3100 genetic analyzer (Applied Biosystems). Protection of the cytosine residues by methylation was apparent by their retention in the bisulfite treated samples. Unmethylated cytosine is converted to uracil and post-PCR sequences as a T. Apparent methylation was observed at 29 positions within the 52 predicted CpGs in the PCR products produced in the control samples. As a comparison only 19 CpGs were methylated in the DNAs extracted from the neonatally programmed duodenums. This 43% reduction in methylation of cytosines in this region may be involved in the increased gene expression observed in the programmed birds relative to controls. Further characterization of the differential methylation patterns is needed both post programming as well as later in life to verify these observations.
This data along with the other preliminary data demonstrating long term effects on performance and gene expression as a result of nutritional neonatal programming in the chick are strong evidence for the role of epigenetics in the regulation of these phenomena. Further work must be conducted to elucidate the specifics of neonatal programming in the chicken, the extent of its effects on DNA methylation of the NaPcoT as well as other genes. Demonstrating these epigenetic effects in the chicken will provide both a significant contribution the understanding of the regulation of genomes, particularly in oviparous species but also provide a potential mechanism for improving performance and the economics of poultry production.
This adaptation or conditioning, which-ever term you choose to use, is a real observable fact for which no underlying mechanism has been previously proposed. Is this new form of genetic effect epigenetic in nature? Studies are ongoing to determine if the mechanism of persistent responses of gene expression to stimuli are epigenetic in nature. If epigenetic regulation is involved in dietary conditioning the opportunities for nutrition to impact both the animal and its offspring are almost limitless.
By helping to understand the interaction between nutrients and molecules in an organism, the implementation of molecular biology and biochemistry in 'classical nutrition' research, followed by the technological revolution of the -omics technologies, will greatly affect nutritional sciences. Unlike biomedical interventions (drug therapy), nutrition is chronic, constantly varying, and composed of a very large amount of known and unknown bioactive compounds. Furthermore, nutrition touches the core of metabolism by supplying the vast majority of ingredients (both macro- and micronutrients) for maintaining metabolic homeostasis. This homeostasis stretches from gene expression to lipid metabolism and from signaling molecules to enzyme cofactors therefore it is essential that nutrition by its nature be studied in an integrated way. Nutrition research in the future will increasingly focus on the ways in which our genes are affected by what we eat or how our animals' genes are affected by what we feed them. Foods and feeds that are safer and more nutritious, new biomedical treatments, and novel ways to help sustain the environment are potential benefits expected to result from these research efforts.
Conclusions
The data included in this summary along with other preliminary data demonstrating long term effects on performance and gene expression as a result of nutritional neonatal programming in the chick are strong evidence for the role of epigenetics in the regulation of these phenomena. Further work must be conducted to elucidate the specifics of neonatal programming in the chicken, the extent of its effects on DNA methylation of the NaPcoT as well as other genes. Demonstrating these epigenetic effects in the chicken will provide both a significant contribution the understanding of the regulation of genomes, particularly in oviparous species but also provide a potential mechanism for improving performance and the economics of poultry production.
This adaptation or conditioning, which-ever term you choose to use, is a real observable fact for which no underlying mechanism has been previously proposed. Is this new form of genetic effect epigenetic in nature? Studies are ongoing to determine if the mechanism of persistent responses of gene expression to stimuli are epigenetic in nature. If epigenetic regulation is involved in dietary conditioning the opportunities for nutrition to impact both the animal and its offspring are almost limitless.
By helping to understand the interaction between nutrients and molecules in an organism, the implementation of molecular biology and biochemistry in 'classical nutrition' research, followed by the technological revolution of the -omics technologies, will greatly affect nutritional sciences. Unlike biomedical interventions (drug therapy), nutrition is chronic, constantly varying, and composed of a very large amount of known and unknown bioactive compounds. Furthermore, nutrition touches the core of metabolism by supplying the vast majority of ingredients (both macro- and micronutrients) for maintaining metabolic homeostasis. This homeostasis stretches from gene expression to lipid metabolism and from signaling molecules to enzyme cofactors therefore it is essential that nutrition by its nature be studied in an integrated way. Nutrition research in the future will increasingly focus on the ways in which our genes are affected by what we eat or how our animals' genes are affected by what we feed them. Foods and feeds that are safer and more nutritious, new biomedical treatments, and novel ways to help sustain the environment are potential benefits expected to result from these research efforts.
Corresponding author: cmashwel@ncsu.edu
- AIHIE SAYER, A.; SYDDALL, H.E.; DENNISON, E.M. et al. Birth weight, weight at 1 y of age, and body composition in older men: findings from the Hertfordshire Cohort Study. American Journal of Clinical Nutrition, v.80, p199-203, 2004.
- ANGEL, R.; ASHWELL, C.M. Dietary conditioning results in improved phosphorus utilization. In: WORLD'S POULTRY CONGRESS, 23., 2008, Brisbane. Proceedings... Brisbane, 2008.
- ANGEL, R.; SAYLOR, W.W.; MITCHELL, A.D. et al. Effect of dietary phosphorus, phytase, and 25-hydroxycholecalciferol on broiler chicken bone mineralization, litter phosphorus, and processing yields. Journal of Poultry Science, v.85, p1200-1211, 2006.
- ASHWELL, C.M.; ANGEL, R. Dietary conditioning results in enduring effects on gene expression. In: WORLD'S POULTRY CONGRESS, 23., 2008, Brisbane, Australia. Proceedings... Brisbane, 2008.
- BAR, A.; SHINDER, D.; YOSEFI, S.; VAX, E. et al. Metabolism and requirements for calcium and phosphorus in the fast-growing chicken as affected by age. British Journal of Nutrition, v.89, p.51-60, 2003.
- BARKER, D.J. The developmental origins of chronic adult disease. ACTA Paediatrica Supplement, v.93, p26-33, 2004.
- BIEHL, R.R.; BAKER, D.H. 1alpha -hydroxycholecalciferol does not increase the specific activity of intestinal phytase but does improve phosphorus utilization in both cecectomized and sham-operated chicks fed cholecalciferol-adequate diets. Journal of Nutrition, v.127, p2054-2059, 1997.
- BLAHOS, J.; CARE, A.D.; SOMMERVILLE, B.S. Effect of low calcium and low phosphorus diets on duodenal and ileal absorption of phosphate in chick. Endocrinologia Experimentalis, v.21, p.59-64, 1987.
- CAPUANO, P.; RADANOVIC, T.; WAGNER, C.A. et al. Intestinal and renal adaptation to a low-Pi diet of type II NaPi cotransporters in vitamin D receptor- and 1alphaOHase-deficient mice. American Journal of Physiology Cell Physiology, v.288, pC429-34, 2005.
- CATTANACH, B.M.; KIRK, M. Differential activity of maternally and paternally derived chromosome regions in mice. Nature, v.315, p.496-498, 1985.
- CHAPMAN, V.; FORRESTER, L.; SANFORD, J. et al. Cell lineage-specific undermethylation of mouse repetitive DNA. Nature, v.307, p.284-286, 1984.
- COOPER, D.N.; ERRINGTON, L.H.; CLAYTON, R.M. Variation in the DNA methylation pattern of expressed and nonexpressed genes in chicken. DNA, v.2, p.131-140, 1983.
- CROMWELL, G.L.; PIERCE, J.L.; STILBORN, H.L. et al. Bioavailability of phosphorus in low-phytic acid corn for chicks. Journal of Poultry Science, v.77, p177, 1998 (Suppl.1).
- DENBOW, D.M.; RAVINDRAN, V.; KORNEGAY, E.T. et al. Improving phosphorus availability in soybean meal for broilers by supplemental phytase. Journal of Poultry Science, v.74, p1831-42, 1995.
- DHANDU, A.S.; ANGEL, R. Broiler nonphytin phosphorus requirement in the finisher and withdrawal phases of a commercial four-phase feeding system. Journal of Poultry Science, v.82, p1257-1265, 2003.
- EDWARDS JR., H.M. Dietary 1,25-dihydroxycholecalciferol supplementation increases natural phytate phosphorus utilization in chickens. Journal of Nutrition, v.123, p.567-577, 1993.
- EDWARDS JR., H.M. Studies on the efficacy of cholecalciferol and derivatives for stimulating phytate utilization in broilers. Journal of Poultry Science, v.81, p.1026-1031, 2002.
- FOX, J.; BUNNETT, N.W.; FARRAR, A.R. et al. Stimulation by low phosphorus and low calcium diets of duodenal absorption of phosphate in betamethasone-treated chicks. Journal of Endocrinology, v.88, p.147-153, 1981.
- FRAGA, M.F.; BALLESTAR, E.; PAZ, M.F. et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proceedings of the National Academy of Sciences USA, v.102, p.106-109, 2005.
- GARDINER-GARDEN, M.; FROMMER, M. CpG islands in vertebrate genomes. Journal of Molecular Biology, v.196, p.261-282, 1987.
- HALES, C.N.; BARKER, D.J. The thrifty phenotype hypothesis. British Medical Bulletin, v.60, p.5-20, 2001.
- HALES, C.N.; BARKER, D.J. Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty phenotype hypothesis. Diabetologia, v.35, p.595-601, 1992.
- HEGSTED, D.M.; MOSCOSO, I.; COLLAZOS, C. A study of the minimum calcium requirements of adult men. Journal of Nutrition, v.46, p.181-201, 1952.
- KATZ, A.; MEIRI, N. Brain-derived neurotrophic factor is critically Involved in thermal-experience-dependent developmental plasticity. Journal of Neuroscience, v.26, p.3899-3907, 2006.
- LABUNSKAY, G.; MEIRI, N. R-Ras3/(M-Ras) is involved in thermal adaptation in the critical period of thermal control establishment. Journal of Neurobiology, v.66, p56-70, 2006.
- LAU, M.M.; STEWART, C.E.; LIU, Z. et al. Loss of the imprinted IGF2/cation-independent mannose 6-phosphate receptor results in fetal overgrowth and perinatal lethality. Genes and Development, v.8, p2953-63, 1994.
- LI, E.; BEARD, C.; JAENISCH, R. Role for DNA methylation in genomic imprinting. Nature, v.366, p.362-365, 1993.
- LI, L.-C.; DAHIYA, R. MethPrimer: designing primers for methylation PCRs. Bioinformatics, v.18, p.1427-1431, 2002.
- LI, Y.C.; LEDOUX, D.R.; VEUM, T.L. et al. Effects of low phytic acid corn on phosphorus utilization, performance, and bone mineralization in broiler chicks. Journal of Poultry Science, v.79, p.1444-1450, 2000.
- MAEDA, N.; HAYASHIZAKI, Y. Genome-wide survey of imprinted genes. Cytogenetic Genome Research, v.113, p.144-152, 2006.
- MONTECUCCOLI, G.; HURWITZ, S.; COHEN, A. et al. The role of 25-hydroxycholecalciferol-1-hydroxylase in the responses of calcium absorption to the reproductive activity in birds. Comparative Biochemistry and Physiology Part A: Physiology, v.57, p.335-339, 1977.
- MORRISSEY, R.L.; WASSERMAN, R.H. Calcium absorption and calcium-binding protein in chicks on differing calcium and phosphorus intakes. American Journal of Physiology, v.220, p.1509-1515, 1971.
- NATIONAL RESEARCH COUNCIL - NRC. Nutrient requirements of poultry 9.rev.ed. Washington, D.C.: National Academy Press, 1994.
- NELSON, T.S. Thy hydrolysis of phytate phosphorus by chicks and laying hens. Journal of Poultry Science, v.55, p2262-4, 1976.
- NEWKIRK, R.W.; CLASSEN, H.L. In vitro hydrolysis of phytate in canola meal with purified and crude sources of phytase. Animal Feed Science and Technology, v.72, p.315-327, 1998.
- NOLAN, C.M.; KILLIAN, J.K.; PETITTE, J.N. et al. Imprint status of M6P/IGF2R and IGF2 in chickens. Developmental Genes and Evolution, v.211, p.179-183, 2001.
- PFAFFL, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research, v.29, p.45, 2001.
- RAPPOLEE, D.A.; STURM, K.S.; BEHRENDTSEN, O. et al. Insulin-like growth factor II acts through an endogenous growth pathway regulated by imprinting in early mouse embryos. Genes and Development, v.6, p.939-952, 1992.
- RAZIN, A. CpG methylation, chromatin structure and gene silencing[mdash]a three-way connection. European Molecular Biology Organization Jounal, v.17, p.4905-4908, 1998.
- RAZIN, A.; RIGGS, A.D. DNA methylation and gene function. Science, v.210, p.604-610, 1980.
- ROBERTSON, K.D.; WOLFFE, A.P. DNA methylation in health and disease. Nature Reviews Genetics, v.1, p.11-19, 2000.
- ROUNDS, M.A.; NIELSEN, S.S. Anion-exchange high-performance liquid chromatography with post-column detection for the analysis of phytic acid and other inositol phosphates. Journal of Chromatography A, v.653, p.148-152, 1993.
- RUVINSKY, A. Basics of gametic imprinting. Journal of Animal Science, v.77, p.228-237, 1999 (suppl. 2).
- STATISTICAL ANALYSES SYSTEM - SAS. JMP 5.1. Cary: SAS Institute, 2004.
- SEGAWA, H.; KANEKO, I.; YAMANAKA, S. et al. Intestinal Na-Pi cotransporter adaptation to dietary Pi content in vitamin D receptor null mice. American Journal of Physiology Renal Physiology, v.287, p.F39-47, 2004.
- SHARPLEY, A. Agricultural phosphorus, water quality, and poultry production: are they compatible? Journal of Poultry Science, v.78, p.660-673, 1999.
- SIMONS, P.C.M.; VERSTEEGH, H.A.J.; JONGBLOED, A.W. et al. Improvement of phosphorus availability by microbial phytase in broilers and pigs. British Journal of Nutrition, v.64, p525-540, 1990.
- SPIEGELMAN, B.M.; HEINRICH, R. Biological control through regulated transcriptional coactivators. Cell, v.119, p157-67, 2004.
- SUGIURA, S.H.; MCDANIEL, N.K.; FERRARIS, R.P. In vivo fractional Pi absorption and NaPi-II mRNA expression in rainbow trout are upregulated by dietary P restriction. American Journal of Physiology Regulative Integrative and Comparative Physiology, v.285, pR770-781, 2003.
- TAMASHIRO, K.L.; WAKAYAMA, T.; AKUTSU, H. et al. Cloned mice have an obese phenotype not transmitted to their offspring. Nature Medicine, v.8, p.262-267, 2002.
- TUKEY, J. The philosophy of multiple comparisons. Statistical Sciences, v.6, p.100-116, 1991.
- VOGTMANN, H.; PFIRTER, H.P.; PRABUCKI, A.L. A new method of determining metabolisability of energy and digestibility of fatty acids in broiler diets. British Journal of Poultry Science, v.16, p.531-534, 1975.
- WALDROUP, P.W.; KERSEY, J.H.; SALEH, E.A. et al. Nonphytate phosphorus requirement and phosphorus excretion of broiler chicks fed diets composed of normal or high available phosphate corn with and without microbial phytase. Journal of Poultry Science, v.79, p.1451-1459, 2000.
- WANG, Z.Q.; FUNG, M.R.; BARLOW, D.P. et al. Regulation of embryonic growth and lysosomal targeting by the imprinted Igf2/Mpr gene. Nature, v.372, p.464-467, 1994.
- YAHAV, S.; MCMURTRY, J.P. Thermotolerance acquisition in broiler chickens by temperature conditioning early in life-the effect of timing and ambient temperature. Journal of Poultry Science, v.80, p.1662-1666, 2001.
- YAJNIK, C.S. Obesity epidemic in India: intrauterine origins? Proceedings of the Nutrition Society, v.63, p.387-396, 2004.
- YAN, F.; ANGEL, R.; ASHWELL, C. et al. Evaluation of the broiler's ability to adapt to an early moderate deficiency of phosphorus and calcium. Journal of Poultry Science, v.84, p.1232-1241, 2005.
- YAN, F.; KERSEY, J.H.; WALDROUP, P.W. Phosphorus requirements of broiler chicks three to six weeks of age as influenced by phytase supplementation. Journal of Poultry Science, v.80, p.455-459, 2001.
- YOUNG, L.E.; BEAUJEAN, N. DNA methylation in the preimplantation embryo: the differing stories of the mouse and sheep. Animal Reproduction Science, v.82-83, p.61-78, 2004.
- YOUNG, L.E.; FERNANDES, K.; MCEVOY, T.G. et al. Epigenetic change in IGF2R is associated with fetal overgrowth after sheep embryo culture. Nature Genetics, v.27, p.153-154, 2001.
Nutritional genomics: a practical approach by early life conditioning with dietary phosphorus
Publication Dates
-
Publication in this collection
09 Aug 2010 -
Date of issue
July 2010