Abstract
The experiment was carried out on Cynodon spp cv. Tifton 85 pastures grazed by sheep under rotational stocking, with the objective of evaluating the structural characteristics as well as the forage mass of the pastures subjected to three grazing intensities in successive cycles. Treatments were composed of three residual leaf area indices (rLAI; 2.4; 1.6 and 0.8), allocated in completely randomized blocks with seven replications, totaling 21 experimental units. Tiller population density, pasture height, leaf area index, forage morphological composition and pasture forage mass were evaluated. The rLAI modified the tiller population density, which increased linearly with decrease in the rLAI of the pastures. Dry masses of leaf blade, stem and dead material were inferior when the rLAI imposed were lower, which resulted in differentiated forage production among the treatments. Tifton 85 pastures grazed by sheep in rotational stocking under tropical conditions with different rLAI show a modified sward structure over successive grazing cycles, mainly by alteration in the height and LAI of the plants at pre-grazing and by light interception post-grazing, which change the tiller population density. The residual leaf area index of 1.6 is the most suitable for pasture management for being equivalent to the heights of entrance and exit of animals on and from paddocks of 33 and 19 cm, respectively, which avoid great accumulation of dead material and excessive stem elongation, in addition to ensuring tillering in the sward.
leaf area index; light interception; sheep; rotational stocking; tillers
FORAGE CROPS
Structural characteristics and forage mass of Tifton 85 pastures managed under three post-grazing residual leaf areas
Wilton Ladeira da SilvaI; Leandro GalzeranoI; Ricardo Andrade ReisII; Ana Cláudia RuggieriII
IPrograma de Pós-graduação em Zootecnia, Universidade Estadual Paulista "Júlio de Mesquita Filho" (UNESP), 14884-900, Jaboticabal, SP, Brazil. CNPq fellow students
IIDepartamento de Zootecnia, Universidade Estadual Paulista "Júlio de Mesquita Filho" (UNESP), Jaboticabal, SP, Brazil
ABSTRACT
The experiment was carried out on Cynodon spp cv. Tifton 85 pastures grazed by sheep under rotational stocking, with the objective of evaluating the structural characteristics as well as the forage mass of the pastures subjected to three grazing intensities in successive cycles. Treatments were composed of three residual leaf area indices (rLAI; 2.4; 1.6 and 0.8), allocated in completely randomized blocks with seven replications, totaling 21 experimental units. Tiller population density, pasture height, leaf area index, forage morphological composition and pasture forage mass were evaluated. The rLAI modified the tiller population density, which increased linearly with decrease in the rLAI of the pastures. Dry masses of leaf blade, stem and dead material were inferior when the rLAI imposed were lower, which resulted in differentiated forage production among the treatments. Tifton 85 pastures grazed by sheep in rotational stocking under tropical conditions with different rLAI show a modified sward structure over successive grazing cycles, mainly by alteration in the height and LAI of the plants at pre-grazing and by light interception post-grazing, which change the tiller population density. The residual leaf area index of 1.6 is the most suitable for pasture management for being equivalent to the heights of entrance and exit of animals on and from paddocks of 33 and 19 cm, respectively, which avoid great accumulation of dead material and excessive stem elongation, in addition to ensuring tillering in the sward.
Key words: leaf area index, light interception, sheep, rotational stocking, tillers
Introduction
The adoption of rotational stocking can be regarded as an accurate and efficient technique aiming to minimize losses in animal raising systems on pastures, be these losses originated from pastures or grazing animals, in addition to improving productivity in these systems. For this grazing system to succeed, it is necessary that some prerequisites, such as the ideal moment for interruption of pasture regrowth, be well defined (Da Silva & Nascimento Júnior, 2007). Another important aspect is the right moment to stop grazing, so as to ensure a minimum residual leaf area, which, according to Ward & Blaser (1961), is capable of promoting growth of plants until the following defoliation, which should also be stipulated.
Parsons et al. (1988) observed that 95% light interception (LI) by the forage sward can be utilized satisfactorily for the interruption of pasture regrowth. In Brazil, scientific research has demonstrated the utilization of this LI on the pasture management in tropical regions (Da Silva et al., 2009; Cunha et al., 2010; Portela et al., 2011).
The use of residual leaf area index (rLAI) has recently been studied for the interruption of grazing under rotational stocking (Cutrim Junior, 2007; Cutrim Junior et al., 2010). This rLAI related to the LI by the forage sward has promoted the understanding of forage production and aided in the management practices adopted. The remaining leaf area is the photosynthetic tissue that remains after grazing and can represent the balance between photosynthesis and plant respiration, allowing the new growth of plants to proceed (Jacques, 1994).
Tiller population density is a structural variable of the pasture that constitutes the LAI and which is essential for the growth of grasses (Lemaire & Chapman, 1996); therefore, any alteration in the tillering demographic patterns, either from cutting or grazing, will promote variations in the pasture leaf area. These alterations will affect the sward capacity of intercepting the incoming light, and might also generate distinct responses in forage accumulation.
The objective of this study was to evaluate the structural characteristics and the forage mass of Tifton 85 pastures subjected the three grazing intensities during successive grazing cycles.
Material and Methods
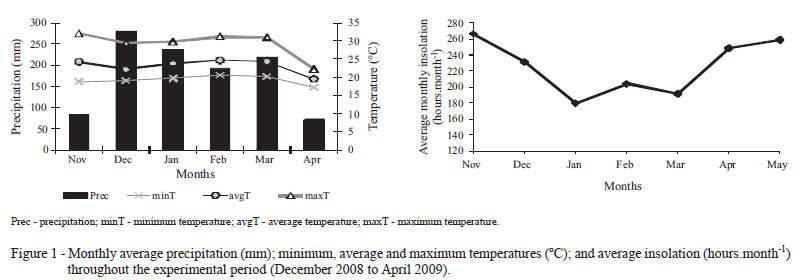
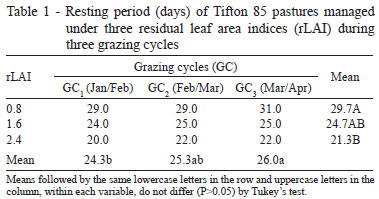
The research was conducted at Universidade Estadual Paulista (UNESP), Campus Jaboticabal, São Paulo, Brazil. The prevailing climate is the Awa type (tropical dry winter), with dry season defined between the months of April and September and concentration of rainfall from October to March, according to the Köppen classification. In the experimental period, comprised between December 2008 and April 2009, the total precipitation was of 1,078.0 mm, with lowest rainfall verified from November 2008 to April 2009 (Figure 1). Minimum, average and maximum temperatures were, on average, 19.28; 29.28 and 23.15 ºC, respectively. The average monthly insolation varied from 180 to 265 hours.month-1 during the experimental period.
The soil of the experimental area was classified as a Dystrophic Red Latosol (Embrapa, 2006), with the following characteristics: pH CaCl2: 4.95; Presin: 14 mg dm-3 ; K+: 3.6 mmolc dm-3; organic matter: 32.5 g dm-3; H+ + Al+3: 40 mmolc dm-3; Ca2+: 31.0 mmolc dm-3; Mg2+: 13.0 mmolc dm-3 and base saturation: 59.0%. Considering the results of soil analysis, there was no need for fertilization for correction of the soil.
The experimental area was an approximately 4,000 m2 Tifton 85 (Cynodon spp.) pasture delimited by a 1.20 m high farm fence mesh. Three grazing intensities defined by the residual leaf area indices (rLAI; 2.4, 1.6 and 0.8) were evaluated in a completely randomized block design with seven replications, totaling 21 paddocks.
The choice for the LAI values and their amplitude as determiners of interruption of animal grazing were based on scientific studies, which reported Tifton 85 LAI varying from 0.59 to 4.48 under different grazing intensities in continuous grazing (Fagundes et al., 1999; Sbrissia et al., 2003). However, no reference of LAI for Tifton 85 pastures managed under rotational grazing was found.
The areas of the paddocks in each treatment were differentiated according to the grazing intensity imposed, as follows: 135 m2 paddocks with pastures maintained under a rLAI of 0.8; 175 m2 paddocks with pastures under a rLAI of 1.6; and 235 m2 paddocks with pastures subjected to a rLAI of 2.4.
The grazing method adopted was rotational stocking with a resting period for pastures that varied in function of the light interception (LI) of 95%. For grazing, Ile de France and Santa Ines sheep of approximately 40 kg were utilized. Animals grazed approximately 11 hours per day. The technique of mob-stocking (Allen et al., 2011) was adopted, inserting or removing the animals on and from paddocks for the occupation days to always be three per paddock. All treatments were evaluated at three grazing cycles (GC) between December 2008 and April 2009, discarding grazing cycle 0 (GC0), when the treatments were implanted.
The monitoring of LI was done at the moment the animals left the paddocks and during pasture regrowth, until the pre-grazing target of 95% LI was reached. Leaf area index (LAI) was monitored at pre-grazing and grazing; in the latter, each paddock with its respective treatment was measured daily until the pre-established rLAI was reached. Once it was true, animals were led to another paddock that already presented 95% LI. For the assessment of LI and LAI, sward analyzer AccuPAR LP-80 (Decagon® ) was used. The device is composed of light sensors that capture the photosynthetically active radiation (PAR) incident on the vegetation. This machine estimates the LAI values in a non-destructive indirect manner, once there is no need for collection of forage samples.
The LAI terminology adopted is a measurement of the device which aims at defining the grazing interruption criterion, in the case of rLAI. The variable LAI provided by the device can include other morphological fractions of the pastures such as stems and dead materials, because Tifton 85 is made up of prostrate plants.
Readings were taken at 20 sampling points per paddock, with measurements always from 11h00 to 13h00. Each point had a reading obtained above and another below the sward.
Sward height measurements were performed with a stick ruler graduated in centimeters. Readings were taken at 50 random points per paddock, both before and after grazing; each point corresponded to the highest average height of the plane of leaves around the ruler. Thus, the average sward heights under each rLAI imposed were obtained. The height of animal entrance on the paddocks was named preHGT, and postHGT was the residual height of the swards after the animals were removed, both in cm.
The estimates of total dry mass (TDM) and dry masses of stem (SDM), dead material (DMDM) and leaf blade (LBDM) in kg DM/ha, both pre- and post-grazing, were obtained by collecting two forage samples per paddock, according to the protocol of mass evaluation, always at points representative of the average sward height. The choice for collecting two representative points per paddock was because of the reduced areas of the paddocks, so that the pasture structure would not be damaged by the successive pre- and post-grazing collections, and also because of the number of repetitions per treatment (seven). Thus, the estimate of forage mass in each treatment was firstly obtained by the average of two collected samples, and the final forage mass value in each treatment was determined by the average of the seven repetitions.
At collections, metal hoops of 0.25 m2 diameter were placed on the soil and all the forage within them was cut at the soil level. Next, samples were taken to the laboratory, where they were weighed. Subsequently, samples were manually separated into fractions leaf blade, stem + sheath and dead material. Each fraction was weighed and dried at 55 ºC for 72 hours in a forced circulation oven.
The evaluations concerning the tiller population densities (TPD) and rates of appearance (TAR) and mortality (TMR) were performed by fixing two PVC tubes of 10 cm in height and 15 cm in diameter on the soil of each paddock at a place corresponding to the average sward height. All tillers existing within the PVC ring were monitored after the end of animal grazing in each paddock, by tagging the new tillers with colored wires, distinguishing the tiller generations with colors. Thus, the total number of tillers in each area delimited by the PVC ring was determined; the average of two PVC rings provided the total number of tillers in each paddock, and the final value in each treatment was obtained by the average of the seven repetitions.
The rates of appearance and mortality (TAR and TMR) in percentage of the total number of tillers were calculated as: TAR = (emerged tillers/total live tillers at the previous tagging) × 100 and TMR = (dead tillers/total live tillers at the previous tagging) × 100, as described by Carvalho et al. (2000).
The statistical analyses were conducted by procedure MIXED of statistical software SAS (Statistical Analysis System, version 9.2). The effects of rLAI, grazing cycle and their information were considered fixed and the block effect was considered random. For a comparison of means, orthogonal contrasts at 5% probability were used. Subsequently, the linear regression equations were fitted for better comprehension of the results.
Results and Discussion
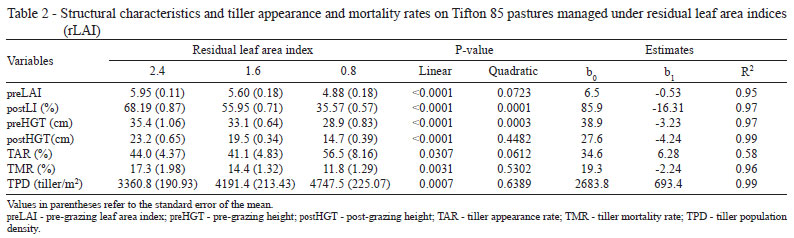
Residual leaf area index affected (P<0.05) the resting period of pastures; it was longer (29.7 days) in the swards managed under a rLAI of 0.8 (Table 1). Pastures managed under rLAI of 2.4 took on average 21.3 days to reach 95% LI and those managed under rLAI of 1.6 took 24.7 days. The resting periods for pastures varied between 24.3 and 26.0 days according to the grazing cycles.
For the pasture to reach 95% LI it must produce a number of leaves capable of intercepting more light, which requires a longer resting period (Cutrim Junior, 2007). Because they have more remaining leaves after grazing, pastures managed under 2.4 rLAI probably utilized the energy reserves at a smaller proportion, which might have contributed to a faster regrowth, unlike the pastures managed under 0.8 rLAI.
A little increase in the resting period was observed as the grazing cycles advanced, with the lowest value (24.3 days) in GC1. This lowest value is probably associated with the fact that pastures still possess enough reserves from the previous grazing cycle or from the grazing at implementation of treatments for faster restructuring of the new forage sward at GC1. In addition to this, the greater monthly average insolation observed in November and December (Figure 1) might have contributed to the faster regrowth of pastures during GC1.
With the reductions in the rLAI of pastures, linear decrease (P<0.05) was observed in all structural characteristics under assessment (Table 2).
The pre-grazing LAI (preLAI) decreased on average 0.53 with the decrease in rLAI from 2.4 to 0.8. Pastures maintained higher (2.4 and 1.6 rLAI) showed higher preLAI: 5.95 and 5.61, respectively. These differences observed in the preLAI values, according to Cooper (1983), can be explained by sward characteristics such as size of the leaf, angle of intersection between the leaf and the stem, leaf rigidity and tiller population density, which can affect the sward structure and consequently the LAI.
The post-grazing LI varied from 68.19 to 35.57% in function of the rLAI imposed, with average reduction of 16.31% among the rLAI. Such variation was somewhat expected, since decrease in LAI promotes drop in LI, because these two variables are greatly correlated (Cutrim Junior, 2007). The variation in post-grazing LAI can also be attributed to the defoliation intensity applied to each pasture. Pastures with lower defoliation intensity (2.4 rLAI) presented lower loss of leaves and stem due to animal consumption, once these components were observed at greater post-grazing levels (Figure 2), so more light was intercepted post-grazing. Grazing sessions with lower defoliation intensity (0.8 rLAI) probably generated more consumption of leaves (Figure 2), which are the component responsible for almost all the light intercepted by the forage sward.
The sward heights at the moment animals entered the paddocks (preHGT) reduced on average 3.23 cm with decrease in the rLAI, with average values of 35.4, 33.1 and 28.9 cm on the pastures under rLAI of 2.4, 1.6 and 0.8, respectively (Table 2).
Residual leaf area index and light interception are the main factors responsible for the maintenance of photosynthesis after grazing, giving the plants conditions of balance between photosynthesis and respiration, allowing for the new growth to be kept with the current product of photosynthesis (Jacques, 1973). Under low rLAI values, as in the case of 0.8 rLAI, in which the average height when animals entered the paddocks was 28.9 cm, there might have been reduction in plant growth due to the lower elongation of stems. Stem elongation is what most influences the height of a sward, as also observed by other authors (Cândido et al., 2005; Barbosa et al., 2007). The greater preHGT in the pastures under 2.4 rLAI can be attributed to the fact that in this situation, plants prioritize the growth of the existing tillers, once the amount of solar radiation on the base of the plants is smaller (Table 2), which significantly reduces the appearance of new tillers (Matthew et al., 1999).
According to Cutrim Junior (2007), controlling the pasture by the height can be used as a practical form of management, for it is an easy method to be adopted. Sward height may not be the most reliable management characteristic, especially in tropical grasses, because the elongation of stems can modify the quantity and quality of the forage available to the animal. However, despite these disadvantages, it is still a practical strategy for the farmer.
The use of LAI would be another viable alternative, provided that their values were transformed into more practical management measures such as height, fixed resting days for the pastures, etc., so there would be more information on the pasture structure, because the LAI measured by AccuPAR, in addition to considering other morphological components of the plants, also takes into account the green leaf blade area, thus being intrinsically related to the photosynthetic process and consequently to forage production (Cândido et al., 2005; Cutrim Junior, 2007).
Post-grazing height (postHGT) in function of the rLAI decreased linearly (4.24 cm) with reduction in the rLAI of the pastures. Under lower grazing intensity, the height when animals left the pastures was 23.2 cm, and under greater intensity (rLAI of 0.8), it was 14.7 cm. This decrease in postHGT in function of rLAI is mainly due to the grazing intensities, once at greater intensities animals tend to consume more forage.
The tiller appearance rates (TAR) on pastures under 2.4 and 1.6 rLAI were similar (44.0 and 41.1%, respectively), with the highest percentage (56.5%) observed on the pastures under 0.8 rLAI. The pastures under rLAI of 0.8 intercepted less light post-grazing (35.57%; Table 2) than the others, which brought about greater incoming light at the base of the sward during the resting period, which might have contributed to the activation of axillary buds and potential appearance of new tillers, once the competition among tillers is mostly for light (Sackville-Hamilton et al., 1995).
Tiller mortality rate (TMR) decreased linearly (P<0.05) as the rLAI reduced. This rate was 31.5% lower on pastures under 0.8 rLAI in comparison with those under 2.4 rLAI. The greater TMR on the pastures kept higher probably occurred because of the greater preLAI observed in these pastures, bringing reduction to a good portion of the incoming light that reached the base of the sward, thus limiting the photosynthetic potential of the leaves located at the base. This fact possibly resulted in the negative carbon balance, because respiration is usually greater than photosynthesis in this situation (Parsons et al., 1983), possibly accelerating the processes senescence and death of leaves, and also causing the death of the tillers that supported them (Sackville-Hamilton et al., 1995).
On average, TPD increased linearly (P<0.05; 693.4 tillers/m2 ) with reduction in pasture rLAI. This trend of pastures kept under lower residual leaf area indices to show lower values for TPD was also verified by other authors studying cultivars of Cynodon and Panicum (Carvalho et al., 2000; Sbrissia et al., 2003; Cutrim Junior, 2007). Such fact can be explained as being a response to the size/population density compensation mechanism of tillers existing in communities of superior plants (Matthew et al., 1995; Sbrissia et al., 2003). When utilizing this mechanism, plants regulate the leaf area of the sward and consequently the capacity to intercept the incoming light. This can promote more shading at the base of the swards managed under lower rLAI, which can decrease the stimulation of basal and axillary buds for production of new tillers.
The GC affected (P<0.05) variables preLAI, postHGT, TAR, TMR and TPD linearly (Table 3).
The pre-grazing leaf area index increased linearly (0.41) along with the GC, possibly because of the increase in the resting periods of pastures (Table 1), which allowed for the swards to have a longer period for reestablishment of the LAI.
Although the postHGT of the pastures decreased with the GC, postLI did not undergo any alteration (P>0.05) with the GC, which can be explained by morphological alterations of the grass and by structural changes of the pastures such as the drop in TAR and TMR and the increase of TPD as the GC advanced.
Both TAR and TMR were greater in GC1, with mean values of 68.05 and 18.68%, respectively, which demonstrates high renovation of the original tiller population, as also observed by Carvalho et al. (2000). Another explanation for these greater values at GC1 may be associated with the adaptation or adjustment of the plant to the new management condition imposed. As the GC developed, the mean values of these two variables decreased linearly. The average precipitation during GC1 was higher in comparison with the subsequent GC (Figure 1), which might have favored plant growth and development, stimulating tillering.
Tiller population density increased linearly (P<0.05; 507.4 tillers/m2 , on average) with the GC (Table 1). Such variation might have been brought about by the increase in the period for resting of pastures over the GC (Table 1), along with the environmental effects that were not constant during the experimental period; especially rainfall and the average monthly insolation, which significantly increased during GC3 (Figure 1). According to Langer (1963), these are direct factors that affect tillering in a plant community.
The total dry matter (TDM) in the pre-grazing conditions was 27.1% higher in the pastures under rLAI of 2.4 in relation to those under 0.8 rLAI; however, 72.1% of this TDM was composed of dry matter of stem (SDM) and dead material (DMDM) (Figure 2). This decrease in TDM at pre-grazing with the reduction in post-grazing rLAI has also been observed in studies with other grasses (Carnevalli et al., 2006; Da Silva et al., 2009).
The management at 2.4 rLAI fits what is called "management for quantity" (Paulino et al., 2004), which is based on the maintenance of the apical meristem and LAI, allowing for a vigorous and fast regrowth which in the present study was approximately 21 days (Table 1), but with accumulation of residual material characterized by the presence of more lignified stems and dead material in excess, capable of undermining the use of forage by sheep.
The highest TDM post-grazing was verified under 2.4 rLAI (3817.2 kg DM/ha). Overall, the component with greatest participation in post-grazing forage mass was the stem, followed by the dead material. In average percentage values, the leaf blade dead material (LBDM) was reduced by 71.82, 76.59 and 79.58% from pre- to post-grazing and SDM by 30.0, 27.8 and 26.9% under 2.4, 1.6 and 0.8 rLAI, respectively. The dead material dry matter, however, was almost unchanged from pre- to post-grazing, given that it is the least useful fraction for the animals during the rainy period. These percentages indicate the high selectivity by sheep.
The dry matter of stem and dead material at pre-grazing were at some point 29.8 an 47.6% respectively superior in the pastures managed under 2.4 rLAI, compared with 0.8 rLAI; in the pastures managed under 1.6 rLAI, these components had intermediate values of 2453 and 1274 kg DM/ha. These differences are mainly due to the grazing intensity imposed, making the grasses under 0.8 rLAI lower more in comparison with those under 1.6 and 2.4 rLAI. In the pastures kept under 2.4 rLAI, greater values of preLAI and preHGT were observed (Table 2), which might have caused the self-shading at the base of the sward, resulting in greater DMDM. Carnevalli et al. (2001) verified that the Tifton 85 pastures subjected to the highest grazing intensities showed the lowest percentage of dead material. In treatments under lower grazing intensities in turn pastures presented greater self-shading and percentage of dead material. Thus, one can affirm that if the objective is to identify the best moment to remove the animals from pastures so that they have access to better quality forage in the next grazing cycle, adopting 2.4 rLAI rather than only having forage in great amounts would not be indicated, since the fractions stem and dead material, when in excess, are the least desirable fractions in pastures of higher quality. On the other hand, utilizing a residual leaf area index of 0.8 promotes lower percentages of the fractions stem and dead material, in addition to the fraction leaf, and also greater resting period for the pastures.
Pastures kept under 2.4 rLAI showed greater LBDM (2004 kg DM/ha) at pre-grazing, once the prerLAI was higher in those pastures (Table 2); the same pattern was observed post-grazing.
After the first grazing cycle (GC1), LBDM at pregrazing increased by 27.6% between CP1 and CP3.
This increase was caused by the higher LAI values (Table 3) found in the last two cycles at pre-grazing. The grazing for implantation of treatments (grazing cycle zero) might have contributed to a better distribution of light within the sward, resulting in increase in LBDM at GC2 and GC3 (Figure 3).
At post-grazing, SDM and LBDM remained constant as the grazing cycles advanced. Only DMDM decreased with the GC, reducing from 1277 to 936 kg DM/ha. A possible explanation for this decrease in DMDM is the reduction in TMR with the grazing cycles (Table 3), because when tiller mortality reduced, there was decrease in the deposition of dead material on the pastures.
Conclusions
The structural characteristics of pastures are affected by the grazing intensities imposed by residual leaf area indices during successive grazing cycles. Interruptions in grazing with 0.8 and 2.4 residual leaf area indices compromise the structure and forage mass of pastures. However, a residual leaf area index of 1.6 favors this structure in relation to management, with aims at the non-compromising of pastures, for promoting a forage mass consistent with the quantitative and qualitative management by avoiding great accumulation of dead material and excessive elongation of the stems, in addition to guaranteeing tillering of the sward. A residual leaf area index of 0.8 promotes low forage mass production, while 2.4 promotes high production, but with excess in fractions stem and dead material, in addition to a low tiller population density. The interruption of pasture regrowth, i.e., the right moment to start grazing, should take place when pastures have an average height of about 33 cm and grazing interruption at around 19 cm, equivalent to a residual leaf area index of 1.6.
Received December 16, 2011 and accepted November 7, 2012.
Corresponding author: wiltonladeira@yahoo.com.br
- ALLEN, V.G.; BATELLO, C.; BERRETTA, et al. An international terminology for grazing lands and grazing animals. Grass and Forage Science, v.66, p.2-28, 2011.
- BARBOSA, R.A.; NASCIMENTO JÚNIOR, D.; EUCLIDES, V.P.B. et al. Capim-tanzânia submetido a combinações entre intensidade e freqüência de pastejo. Pesquisa Agropecuária Brasileira, v.42, p.329-340, 2007.
- CÂNDIDO, M.J.D.; GOMIDE, C.A.M.; ALEXANDRINO, E. et al. Morfofisiologia do dossel de Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Revista Brasileira de Zootecnia, v.34, p.338-347, 2005.
- CARNEVALLI, R.A.; Da SILVA, S.C.; FAGUNDES, J.L. et al. Desempenho de ovinos e respostas de pastagens de Tifton 85 (Cynodon spp.) submetidas a regimes de desfolha sob lotação contínua. Scientia Agricola, v.58, p.7-15, 2001.
- CARNEVALLI, R.A.; Da SILV, S.C.; Bueno, A.A.O. et al. Herbage production and grazing losses in Panicum maximum cv. Mombaça under four grazing managements. Tropical Grasslands, v.40, p.165-176, 2006.
- CARVALHO, C.A.B.; Da SILVA, S.C.; SBRISSIA, A.F. et al. Demografia do perfilhamento e taxas de acúmulo de matéria seca em capim "Tifton 85" sob pastejo. Scientia Agricola, v.57, p.591-600, 2000.
- COOPER, J.P. Physiological and morphological advances for forage improvement. In: INTERNATIONALGRASSLAND CONGRESS, 14., 1981, Lexington. Proceedings... Lexington: Westview Press, Boulder CO., 1983. p.69-73.
- CUNHA, B.A.L.; NASCIMENTO JÚNIOR, D.; SILVEIRA, M.C.T. et al. Effects of two post-grazing heights on morphogenic and structural characteristics of guinea grass under rotational grazing. Tropical Grasslands, v.44, p.253-259, 2010.
- CUTRIM JUNIOR, J.A.A. Crescimento e morfofisiologia do dossel do capim Tanzânia com três frequências de desfolhação e dois resíduos pós-pastejo 2007. 104f. Dissertação (Mestrado em Zootecnia) - Universidade Federal do Ceará, Ceará
- CUTRIM JUNIOR, J.A.A.; CÂNDIDO, M.J.D. VALENTE, B.S.M. et al. Fluxo de biomassa em capim-tanzânia sob três frequências de desfolhação e dois resíduos pós-pastejo. Revista Brasileira de Saúde e Produção Animal, v.11, p.618-629, 2010.
- Da SILVA, S.C.; BUENO, A.A.O.; CARNEVALLI, R.A. et al. Sward structural characteristics and herbage accumulation of Panicum maximum cv. Mombaça subjected to rotational stocking managements. Scientia Agricola, v.66, p.8-19, 2009.
- Da SILVA, S.C.; NASCIMENTO JÚNIOR, D. Avanços na pesquisa com plantas forrageiras tropicais em pastagens: características morfofisiológicas e manejo do pastejo. Revista Brasileira de Zootecnia, v.36, p.122-138, 2007.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA -EMBRAPA. Serviço Nacional de Levantamento e Conservação de Solos. Sistema brasileiro de classificação de solos 2.ed. Rio de Janeiro: Embrapa Solos, 2006. 306p.
- FAGUNDES, J.L.; Da SILVA, S.C.; PEDREIRA, C.G.S. et al. Indice de área foliar, interceptação luminosa e acúmulo de forragem em pastagens de cynodon spp. sob diferentes intensidades de pastejo. Scientia Agricola, v.56, p.1141-1150, 1999.
- JACQUES, A.V.A. Fisiologia do crescimento de plantas forrageiras (área foliar e reservas orgânicas). In: SIMPÓSIO SOBRE MANEJO DA PASTAGEM, 1973, Piracicaba. Anais... Piracicaba: ESALQ, 1973. p.95-101.
- JACQUES, A.V.A. Caracteres morfofisiológicos e suas implicações com o manejo. In: CARVALHO, M.M.; ALVIM, M.J.; XAVIER, D.F. (Eds.) Capim-elefante, produção e utilização Coronel Pacheco: EMBRAPA/CNPGL, 1994. p.31-48.
- LANGER, R.H.M. Tillering in herbage grass. A review. Herbage Abstracts, v.33, p.141-148, 1963.
- LEMAIRE, G.; CHAPMAN, D. Tissue flows in grazed plant communities. In: HODGSON, J.; ILLIUS, A.W. (Eds.) The ecology and management of grazing systems Wallingford: CABI International, 1996. p.3-36.
- MATTHEW, C.; LEMAIRE, G.; SACKVILLE HAMILTON, N.R. et al. A modified self thinning equation do describe size/density relationships for defoliated swards. Annals of Botany, v.76, p.579-587, 1995.
- MATTHEW, C.; ASSUERO, S.G.; BLACK, C.K. et al. Tiller dynamics of grazed swards. In: SIMPÓSIO INTERNACIONAL GRASSLAND ECOPHYSIOLOGYAND GRAZING ECOLOGY, 1., 1999, Curitiba. Anais... Curitiba: UFPR, 1999. p.109-133.
- PARSONS, A.J.; LEAFE, E.L.; COLLETT, B. et al. The phisiology of grass production under grazing. Characteristics of leaf and canopy photosynthesis of continuosly grazed sward. Journal Applied Ecology, v.20, p.117-126, 1983.
- PARSONS, A.J.; JOHNSON, I.R.; HARVEY, A. Use of a model to optimize the interaction between frequency and severity of intermittent defoliation to provide a fundamental comparison of the continuous and intermittent defoliation of grass. Grass and Forage Science, v.43, p.49-59, 1988.
- PAULINO, M.F. FIGUEIREDO, D.M.; MORAES, E.H.B.K. et al. Suplementação de bovinos em pastagens: Uma visão sistêmica. In: SIMPÓSIO DE PRODUÇÃO DE GADO DE CORTE, 4., 2004, Viçosa, MG. Anais... Viçosa, MG: Universidade Federal de Viçosa, 2004. p.93-144.
- PORTELA, J.N.; PEDREIRA, C.G.S.; BRAGA, G.J. Demografia e densidade de perfilhos de capim-braquiária sob pastejo em lotação intermitente. Pesquisa Agropecuária Brasileira, v.46, p.315-322, 2011.
- SACKVILLE-HAMILTON, N.R.; MATTHEW, C.; LEMAIRE, G. In: Defence of the-3/2 boundary rule: a re-evaluation of self thinning concepts and status. Annals of Botany, v.76, p.569-577, 1995.
- SBRISSIA, A.F.; Da SILVA, S.C.; MATTHEW, C. et al. Tiller size/density compensation in grazed Tifton 85 bermudagrass swards. Pesquisa Agropecuária Brasileira, v.38, p.1459-1468, 2003.
- WARD, C.Y.; BLASER, R.E. Carboydrate food reserves and leaf area in regrowht of Orchardgrass. Crop Science, v.1, p.366-370, 1961.
Publication Dates
-
Publication in this collection
05 Apr 2013 -
Date of issue
Apr 2013
History
-
Received
16 Dec 2011 -
Accepted
07 Nov 2012