Abstract
Several changes in amino acid levels occur during an infection. As a rule, glycine, taurine and serine levels diminish considerably, while cysteine levels increase. Such changes may be attributed to the intense consumption of sulfur amino acids (SAA- methionine+cysteine) during infectious challenge. Methionine plays an important role in humoral and cellular immune responses. It has been suggested that such effect is exerted by intracellular glutathione and cysteine levels. Four-hundred thirty-two day-old Ross male broiler chickens were fed (from 1 to 42 days of age) three SAA levels in the diet (0.72, 0.82, and 0.92% from 1 to 21 days of age; 0.65, 0.75, and 0.85% from 22 to 42 days of age) and submitted to two immunological stimulus series. Vaccines against Marek's disease, Fowlpox, Infectious Bronchitis and Infectious Bursal disease, Freund's Complete Adjuvant, and avian tuberculin were used as immunological stimuli. The experiment comprised 6 treatments, with 6 replications using 12 birds per replicate. Performance data were collected weekly. Gumboro antibodies were measured by ELISA, and the cellular immune response by the tuberculin test. SAA levels tested did not influence immune response. Nevertheless, the vaccines applied on the 1st day impaired chick performance up to the 21 days of age. The SAA levels generally adopted in poultry husbandry may not be enough to assure weight gain, especially when birds are raised if a low-challenge infectious environment.
immune system; immunological stimulus; methionine; performance; poultry
Influence of sulfur amino acid levels in diets of broiler chickens submitted to immune stress
Rubin LLI; Ribeiro AMLII; Canal CWIII; Silva ICIV; Trevizan LIV; Vogt LKIV; Pereira RAV; Lacerda LV
IDepartamento de Patologia Clínica Veterinária
IIUFRGS Professors. CNPq researchers. Laboratorio de Ensino Zootécnico, Departamento de Zootecnia. Universidade Federal do Rio Grande do Sul UFRGS. Porto Alegre, RS, Brazil
IIIUFRGS Professors. CNPq researchers. Departamento de Patologia Clínica Veterinária
IVGraduated studentes, PPG-Zootecnia, UFRGS. Laboratorio de Ensino Zootécnico, Departamento de Zootecnia. Universidade Federal do Rio Grande do Sul UFRGS. Porto Alegre, RS, Brazil
VGraduated studentes, PPG-Veterinária, UFRGS. Departamento de Patologia Clínica Veterinária
Mail Address Mail Address Laurício L. Rubin Departamento de Zootecnia, UFRGS Av. Bento Gonçalves 7712 Porto Alegre, RS, Brazil E-mail: llrubin@hotmail.com
ABSTRACT
Several changes in amino acid levels occur during an infection. As a rule, glycine, taurine and serine levels diminish considerably, while cysteine levels increase. Such changes may be attributed to the intense consumption of sulfur amino acids (SAA- methionine+cysteine) during infectious challenge. Methionine plays an important role in humoral and cellular immune responses. It has been suggested that such effect is exerted by intracellular glutathione and cysteine levels. Four-hundred thirty-two day-old Ross male broiler chickens were fed (from 1 to 42 days of age) three SAA levels in the diet (0.72, 0.82, and 0.92% from 1 to 21 days of age; 0.65, 0.75, and 0.85% from 22 to 42 days of age) and submitted to two immunological stimulus series. Vaccines against Marek's disease, Fowlpox, Infectious Bronchitis and Infectious Bursal disease, Freund's Complete Adjuvant, and avian tuberculin were used as immunological stimuli. The experiment comprised 6 treatments, with 6 replications using 12 birds per replicate. Performance data were collected weekly. Gumboro antibodies were measured by ELISA, and the cellular immune response by the tuberculin test. SAA levels tested did not influence immune response. Nevertheless, the vaccines applied on the 1st day impaired chick performance up to the 21 days of age. The SAA levels generally adopted in poultry husbandry may not be enough to assure weight gain, especially when birds are raised if a low-challenge infectious environment.
Keywords: immune system; immunological stimulus; methionine; performance; poultry.
INTRODUCTION
Minimum requirements for a given nutrient for maximum production are fully established (NRC, 1994). Yet, whether these requirements that maximize the performance of healthy unchallenged animals are in fact enough to trigger an optimal immune response in naive animals, thus making these individuals more resistant, still needs to be determined. Therefore, knowing the underlying nutritional mechanisms that influence the immune system is essential to understand the complex interactions between diet and disease.
The main role played by the bird immune system is to recognize and to promote the elimination of infectious agents that challenge the system. Alternatively, if this fails, the immune system is required to guarantee the compatible conditions of recovery and adaptation with minimum losses to the production cycle (Montassier, 2000). The resistance against infectious challenges requires an intense response orchestrated by the immune system. From the nutritional standpoint, feed substrates (amino acids, energy, enzymes, etc.) are needed to trigger such a response, which consists in clonal proliferation of lymphocytes, establishment of germinative centers in the bursa of Fabricius to refine immunoglobulin affinity, recruitment of new bone marrow monocytes and heterocytes, and synthesis of effector molecules (immunoglobulins, nitric oxide, lysozyme, complement) and communication molecules (cytokines and eicosanoids, for instance).
Infections lead to several changes in amino acid plasma levels. Such alterations are ascribed to a sharp consumption of sulfur amino acids (SAA) during the infectious challenge, imputable to the metabolic relationships these amino acids maintain with each other (Jeevanandam et al. 1990; Paaw & Davis, 1990).
Methionine is an essential amino acid that plays at least four main roles. First, methionine takes part in protein synthesis. Second, methionine is a precursor for glutathione, a tripeptide that reduces reactive oxygen species (ROS), and thus protects cells from oxidative stress. Third, methionine is needed for the synthesis of polyamines (spermine and spermidine), which take part in nucleus and cell division events. Fourth, methionine is the most important methyl group donor to methylation reaction of DNA and other molecules. Therefore, methionine is the first limiting amino acid in commercial broiler diets, which are prepared essentially with corn- and soy-based nutrients (Kidd, 2004). Research has shown that methionine interferes in the immune system, improving both humoral and cellular response. It has also been observed that methionine requirements are higher when the purpose is to maintain optimal immunity levels, as compared to growth (Swain & Johri, 2000; Shini et al., 2005), and that lower SAA levels result in a severe lymphocyte depletion in the intestine tissues (Peyer's patches) and in the lamina propria (Swain & Johri, 2000).
One of the mechanisms proposed to explain methionine interference in the immune system is based on the proliferation of immune cells, which are sensitive to intracellular variations in glutathione and cysteine levels, compounds which also participate in the metabolism of methionine (Shini et al., 2005). Glutathione is, quantitatively speaking, the most abundant intracellular antioxidant compound, playing a variety of important roles and being vitally important for the protection against the emergence of oxidative stress that follows inflammatory processes (Le Floc'h et al., 2004). Studies carried out with healthy human subjects have hypothesized that oscillations in intracellular glutathione levels are responsible for significant changes in CD4+ T cell counts, a central physiologic parameter that is negatively affected by either the lack or excess of glutathione (Kinscherf et al., 1994).
This study aimed at finding if birds undergoing immunological stress have specific nutritional requirement as compared to non-stressed animals. Different dietary SAA levels were used, by means of methionine hydroxyl-analog supplementation. The pressure the compound exerts on the immune response, as triggered by immunological stimuli in birds, was assessed.
MATERIALS AND METHODS
This study used 432 day-old Ross male broiler chickens. The birds were housed in battery brooders placed in a temperature-controlled room, and were allowed free access to feed and water throughout the growing period (42 days). A basal diet (Table 1) was supplemented with three methionine concentrations: 0.12, 0.22, and 0.32%. Methionine hydroxi-analog (HMTBA liquid-phase methionine), with 88% methionine content, was used as supplement. A 3x2 factorial design was applied: three sulfur amino acid (SAA) levels (0.72, 0.82, and 0.92% for one- to 21-day-old birds, and 0.65, 0.75, and 0.85% for 22- to 42-day-old birds) and two immunological stimuli (IS), affording six treatments with six replicates each, with 12 birds per replicate.
Birds were submitted to two IS series: (i) treatment series (consisting of vaccines commonly used in commercial broiler production, Freund's Complete Adjuvant and bird tuberculin); and (ii) positive-control series (without any stimulus). Birds were vaccinated with one day of age against Marek' disease, Fowlpox, and Infectious Bronchitis. Infectious Bursal Disease vaccine was administered on the 14th day of age, using the live virus vaccine strain V877, with low virus attenuation (minimum doses were 103.0 LD50) mixed in drinking water. This vaccine aimed at triggering a severe inflammatory reaction in the bird's bursa of Fabricius; firstly causing edema and lymphocyte depletion, and then atrophy. On the 28th day of age, twelve birds in each treatment series were challenged with 0.5 mL Freund's Complete Adjuvant (4 mg/mL of Mycobacterium avium) via intramuscular injection in the breast muscle. On the 40th day of age, that is, 12 days after the administration of the adjuvant, tuberculin was inoculated using a 0.1-mL dose via intradermal injection in one of the bird wattles. As birds had previously been sensitized by M. avium, a cell-mediated immune response was triggered, at the administration site (tuberculin reaction).
At 14th, 17th, 35th, and 42nd days of age, six birds of non-stimulated group (one or two birds in each date) were weighed, sacrificed, and submitted to necropsy. Blood, bursas of Fabricius and spleen were collected. Performance data, including weight gain (WG), feed intake (FI), and feed conversion ratio (FCR) were weekly collected.
The immunological parameters measured included humoral immunity (using ELISA for IBD antibodies), and cellular immunity (tuberculin reaction). ELISA for IBD antibodies was conducted in 36 sera per treatment series, classified for bird age (1, 14, 17, 35, and 42 days of age). Optical densities were transformed into antibody titers, using the formula recommended by the kit manufacturer (Symbiotics Corporation). cellular immune response was carried out by measuring the difference in wattle thickness 24 h before and after the administration of avian tuberculin.
Blood smears (eight per treatment series) were performed with blood samples collected on day 42 to quantitatively and qualitatively evaluate blood cells. The bursas of Fabricius collected as of necropsy were weighed, measured using a bursometer (scored 1 to 8), and stored in formaldehyde 10%. The histological sections prepared from the bursas were analyzed and classified according to the lesion caused by the vaccine (lymphocyte depletion) and scored from 0 to 5 (Muskett et al., 1979). Spleens were also weighed.
HMTBA was introduced in the baseline diet only after being premixed with inert vehicle. The mixture was analyzed prior to the elaboration of diets.
The experimental data were analyzed as a completely randomized design using ANOVA of SAS software package (2001), at a 0.05 significance level. When the F test yielded significant results, LS means test was performed to compare treatment means. Optical density data (sera) were antibody-titrated, and these results tested for normality (Shapiro & Wilk, 1965).
RESULTS AND DISCUSSION
During the starter phase (1 to 21 days), no significant interaction between immunological stimulus and SAA levels was observed. As to the main factors, it is possible to observe that the non-vaccinated bird group had better WG and FCR results. Also, the 0.92% SAA concentration yielded the best WG and FCR values for the period, irrespective of stimulus (Table 2). No SAA interference was observed in FI.
Two factors affect the growth rate of broiler chickens during immunological stress: decrease in feed intake, and inefficient metabolism and nutrient absorption. These events are triggered by the bird's own immune response (Klasing et al., 1987). As no reduction in feed intake was observed during the starter phase (1 to 21 days) in the present study, the worse growth rate observed in vaccinated birds is attributed to the inefficient metabolism and nutrition absorption. The underlying cause of this event is the immune response triggered by the vaccines used. Therefore, overall bird requirements increased.
A series of metabolic events may be linked to immunological stimulus, such as bivalent cation exchange, increased amino acid oxidation, increased gluconeogenesis caused by amino acids, and fat build-up (Klasing & Barnes, 1988). Some studies have reported these events as an indirect cause of altered growth rates, which in turn is caused by direct changes in WG, FI, and FCR (Klasing et al., 1987; Powanda, 1977).
SAA levels still did not affect FI during the grower phase (22 to 42 days). As for WG, a positive effect was observed, caused by higher SAA levels (P<0.07). Yet, FCR and the lower SAA levels were positively correlated during the period. In addition, no negative influence caused by immunological stimulus was observed during the grower phase (Table 3).
When the complete rearing period (1 to 42 d) was analyzed, a significant interaction was observed for WG (significance level 8%), indicating that the 0.92/0.85% SAA treatments as resulted in higher WG, and this response was observed only in the non-stimulated group. This demonstrates that the immune response had a negative influence on growth, as previously found in other studies (Klasing et al., 1987; Dinarello, 1984; Klasing, 1984).
Broiler chickens fed the lower SAA levels exhibited poorer FCR. No differences were observed in FI. When the immunological stimulus was considered, no statistically significant difference was observed between the group of vaccinated and the group of non-vaccinated broilers on day 42nd (p>0.05) (Table 4).
In the present study, SAA requirement was 0.92% (0.60% methionine) for the optimal WG in the starter phase, and 0.85% (0.57% methionine) in the grower phase, while Rama Rao et al. (2003) concluded that the SAA requirement for optimal WG did not exceed 0.72% in females reared until the 42nd day of age. Tsiagbe et al. (1987b) showed that SAA requirement for the optimal growth of female broilers was 0.78%, varying between 0.72% and 2.17%. Shini et al. (2005) observed that methionine levels (0.45, 0.60, and 0.75%) did not significantly affect WG in the bird population studied. Other authors concluded that there was no improvement inWG of broilers fed methionine levels above the NRC recommendations (1994) (Pesti et al., 1979; Maatman et al., 1993).
The present study found out that the SAA requirement in the third week of life was 0.92% for optimal WG (Table 5).
As the birds were vaccinated on the 14th day of age against infectious bursal disease, the observed immune response is noteworthy, indicating that higher SAA levels may be beneficial to resilience. A study, using rats as animal model, demonstrated that dietary cystine supplementation decreases weight loss and muscle catabolism during the acute phase immune response, and also improves the recovery of rats after Escherichia coli challenge (Breuillé et al., 2006).
The vaccines injected impaired broiler growth until the 21st day of age; however, weights were not different after the 28th day of life (Figure 1).
Chamblee et al. (1992) also verified lower weight, worse FCR, as well as higher mortality in chickens vaccinated on the 1st day of age against IBD, Newcastle disease, bronchitis, and Marek. Therefore, the immunological stress caused by vaccination in broilers impairs performance, even when no disease is present. In the light of the comparative losses as caused by disease and by vaccination, it is suggested that vaccines be cautiously applied.
The compensatory growth observed in vaccinated birds in the present study wasalso reported in other studies that used feed restriction (Plavnik & Hurvitz, 1985), water restriction (Viola et al., 2005), and heat stress (Ribeiro et al., 2001). Compensation is supposed to occur due to a decrease in energy requirements for bird maintenance during the stress period, and provides the advantage of changing bird growth pattern. In most studies , feed efficiency increased during the period after the stress (Robinson et al., 1992; Palo et al., 1995), but this was not observed in the present study.
As to the analysis of cellular immune response, as evaluated by differential wattle thickness 24 h before and after the injection of bird tuberculin, no statistically significant differences were observed between SAA levels, despite the considerable variation observed for this parameter (Table 6).
Tsiagbe et al. (1987a) observed an increase in mitogenic stimulation by Phytohemaggluthinin-P (PHA-P) in broilers supplemented with methionine, in which the optimal level to induce the best possible cellular response, was 0.60% methionine. Rama Rao et al. (2003) showed that SAA requirement for optimal cellular response was 0.83%. Swain & Johri (2000) demonstrated that cellular immune response by inhibition of leukocyte migration index (LMI) significantly increases when methionine level is 0.81%, varying between 0.36 and 0.81%. The authors used a low weight gain animal model (Synthetic male line 1,300 g in 42 days), different from the animal model used in the present study (Ross 2,900 g in 42 days). The present study tested a maximum dietary methionine content of 0.60%. Shini et al. (2005) observed an improvement in the cellular immune response in1 to 21-day-old broilers with 0.75, 0.90 and 1.05%, SAA levels as compared to the basal level of 0.30%. The authors also concluded that methionine (as offered in the diet) influenced the cellular aspects of immune response, suggesting that the proliferation of immune cells might be influenced by an intracellular variation in glutathione and cysteine levels.
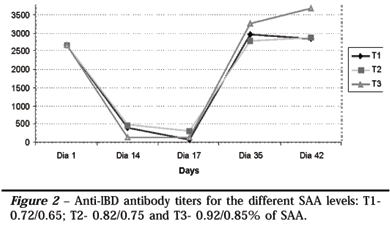
It was observed also that the IBD antibody levels, as analyzed by ELISA, were not significantly different among the different SAA levels (Figure 2).
Similarly, Takahashi et al. (1993) and Takahashi et al. (1994) reported that neither the excess nor the deficiency of methionine in diets influenced primary antibody generation against SRBC in chickens. Rama Rao et al. (2003) showed that methionine supplements increased the anti-SRBC antibody levels, and that the best SAA level for optimal antibody production was 0.88% in an interval 0.72-0.88% tested methionine levels. Tsiagbe et al. (1987a) showed an increase in antibody response against SRBC in chickens fed 0.72 to 0.97% SAA levels
The histological analyses of the bursa of Fabricius for lymphocyte depletion caused by the IBD vaccine did not show any significant difference between SAA levels. Nishizawa (2001) observed lesions in the bursa of Fabricius three days after IBD vaccine was applied by ocular administration as compared to broilers of the same age. The difference in the administration route (drinking water) probably attenuated its effect.
The SAA levels tested in the present study did not influence heterophil:lymphocyte relationship, bursa of Fabricius and spleen weights, orhematological data. The same results were observed by Dunnington et al. (1994) and Rama Rao et al. (2003).
CONCLUSIONS
SAA levels currently used in broiler production may be insufficient to ensure optimal weight gain, especially if the bird is reared in an environment with low infectious challenge. It was also observed that the vaccines usually administered impaired broiler performance until 21 days of age, and that the tested SAA levels failed to improve immune response in broilers.
Arrived: November / 2006
Approved: April / 2007
- Breuillé, D. et al Beneficial effect of amino acid supplementation, especially cysteine, on body nitrogen economy in septic rats. Clinical Nutrition 2006; 25:634642.
- Chamblee TN, Brake JD, Schultz CD, Thaxton JP. Yolk sac absorption and initiation of growth in broilers. Poultry Science 1992; 71:1811-1816.
- Dinarello CA. Interleukin-1 and the pathogenesis of the acute-phase response. New England Journal of Medicine 1984; 311: 1413-1418.
- Dunnington EA, Zulkifli I, Siadak D, Larsen AS, Siengel PB. Dietary methionine and antibody response in broiler cockerels. Archive für Geflúgelksunde 1994; 58:125-129.
- Jeevanandam M, Young DH, Ramais L, Schiller WR. Effect of major trauma on plasma free amino acid concentrations in geriatric patients. American Journal of Clinical Nutrition 1990; 51:1040-1050.
- Klasing KC. Effect of inflammatory agents and interleukin 1 on iron and zinc metabolism. American Journal of Physiology 1984; 247: R901-R904.
- Klasing KC, Barnes DM. Decreased amino acid requirements of grower chicks due to immunological stress. Journal of Nutrition1988; 118:1158-1164.
- Klasing KC, Laurin DE, Peng RK, Fry DM. Immunologically mediated growth depression in chicks: influence of feed intake, corticosterone, and interleukin-1. Journal of Nutrition 1987; 117:1629-1639.
- Kidd MT. Nutritional Modulation of Immune Function in Broilers. Poultry Science 2004; 83:650-657.
- Kinscherf R, Fischbach T, Mihm S, Roth S, Hohenhaus-Sievert E, Weiss C, Edler L, Bärtsch P, Dröge W. Effect of glutathione depletion and oral N-acetyl-cysteine treatment on CD4+ and CD8+ cells. The FASEB Journal 1994; 8:448-451.
- Le Floc'h N, Melchior D, Obled C. Modification of protein and amino acid metabolism during inflammation and immune system activation. Livestock Production Science 2004; 87:37-45.
- Maatman R, Gross WB, Dunnington EA, Larsen AS, Siegel PB. Growth, immune response and behavior of broiler and Leghorn cockerels fed different methionine levels. Archiv für Geflügelkunde 1993; 57:249-256.
- Montassier HJ. Enfermidades do sistema imune. In: Berchieri A, Macari M, editores. Doenças das aves. Jaboticabal: Funep-Unesp; 2000. p. 141-150.
- Muskett JC, Hopkins IG, Edwards KR, Thornton DH. Comparison of two infectious bursal disease vaccine strains and potential hazards in susceptible and maternally immune birds. Veterinary Record 1979; 104:332-334.
- Nishizawa M. Análise morfométrica e histopatológica da bursa de Fabrícius de frangos de corte vacinados com amostras do vírus da doença de Gumboro. Revista Brasileira de Ciência Avícola 2001; Supl. 6:172.
- National Research Council -NRC. Nutrient requirements of poultry. 9th ed. Washington: National Academy Press; 1994.
- Paaw JD, Davis AT. Taurine concentrations in serum of critically injured patients and age- and sex-matched healthy control subjects. American Journal of Clinical Nutrition 1990; 49:814-822.
- Palo PE, Sell JL, Piquer FJ, Sotosalanova MF, Vilaseca L. Effect of early nutrient restriction on broiler chickens. 1. Performance and development of the gastrointestinal tract. Poultry Science 1995; 74:88-101.
- Pesti GM, Harper AE, Sunde ML. Sulfur amino acid and methyl donor status of corn-soy diets fed to starting broiler chicks and turkey poults. Poultry Science 1979; 58:1541-47.
- Plavnik I, Hurwitz S. The performance of broiler chicks during and following a severe feed restriction at an early age. Poultry Science 1985; 64:348-355.
- Powanda MC. Changes in body balances of nitrogen and other key nutrients: description and underlying mechanisms. American Journal of Clinical Nutrition 1977; 30:1254-68.
- Rama Rao SV, Praharaj NK, Reddy MR, Panda AK. Interaction between genotype and dietary concentrations of methionine for immune function in commercial broilers. British Poultry Science 2003; 44: 104-112.
- Ribeiro AML, Penz Jr AM, Teeter RG. Effects of 2-hydroxy-4-(methylthio)butanoic acid as DL-Methionine on broiler performance and compensatory growth after exposure to two different environmental temperatures. Journal of Applied Poultry Research 2001; 10(4):419-426.
- Robinson MK, Rodrick ML, Jacobs DO, Rounds JD, Collins KH, Saporoschetz IB, Mannick JA, Wilmore DW. Glutathione depletion in rats impairs T-cell and macrophage immune function. Archives of Surgery 1993; 128:29-34.
- SAS Institute. SAS user's guide: statistics. Cary: SAS Institute; 1986.
- Shapiro SS, Wilk MB. An analysis of variance test for normality (complete sample). Biometrika Great Britain 1965; 52(3):591-611.
- Shini S, Li X, Bryden WL. Methionine requirement and cell-mediated immunity in chicks. British Journal of Nutrition 2005; 94:746-52.
- Swain BK, Johri TS. Effect of supplemental methionine, choline and their combinations on the performance and immune response of broilers. British Poultry Science 2000; 41:8388.
- Takahashi K, Konashi S, Akiba Y, Horiguchi M. Effects of dietary threonine level on antibody production in growing broilers. Animal Science and Technology 1994; 65:956-960.
- Takahashi K, Konashi S, Akoba Y, Horiguchi M. Effects of marginal excess or deficiency of dietary methionine on antibody production in growing broilers. Animal Science and Technology 1993; 64:13-19.
- Tsiagbe VK, Cook ME, Harper AE, Sunde M.L. Enhanced immune responses in broiler chick fed methionine supplemented diets. Poultry Science 1987a; 66:1147-1154.
- Tsiagbe VK, Cook ME, Harper AE, Sunde ML. Efficacy of cysteine in replacing methionine in the immune responses of broiler chicks. Poultry Science 1987b; 66:1138-1146.
- Viola T, Ribeiro AML, Penz Jr AM.Compensatory water consumption of broilers submitted to water restriction from 1 to 21 days of age. Brazilian Journal of Poultry Science/Revista Brasileira de Ciência Avícola 2005;7(4):243-246.
Publication Dates
-
Publication in this collection
24 July 2007 -
Date of issue
Mar 2007
History
-
Received
Nov 2006 -
Accepted
Apr 2007