Abstracts
In this study, the recolonization speed of the mussels Brachidontes darwinianus and B. solisianus was compared to evaluate their responses to gaps opened by disturbances in the rocky shore. B. darwinianus was much faster than B. solisianus, rapidly closing gaps in the mussel bed. When they co-occurred, distribution patterns of these species were related not only with distinct physiological tolerances but also with responses to disturbance. B. darwinianus was more tolerant to low salinities, dominated estuarine zones generally occuring lower in the midlittoral, and presented a faster response to opened gaps.
Bivalve; Intertidal; Rocky shore; Zonation; Mytilidae
Mexilhões são importantes dominantes do espaço na região entremarés de costões rochosos. Diversos fatores influenciam a distribuição de espécies coexistentes, tais como diferenças de tolerância fisiológica, histórias de vida e uso de recursos distribuídos irregularmente no espaço e tempo. Neste estudo, a velocidade de recolonização dos mexilhões Brachidontes darwinianus e B. solisianus foi comparada para avaliar suas respostas a clareiras abertas por perturbações. B. darwinianus foi muito mais rápido que B. solisianus, fechando rapidamente clareiras abertas no banco de mexilhões. Este resultado sugere que quando estas espécies co-ocorrem, sua distribuição está relacionada não só a tolerâncias fisiológicas distintas, mas também a diferentes respostas a perturbações. B. darwinianus é mais tolerante a baixas salinidades, dominando zonas estuarinas, além de geralmente ocorrer na região mais baixa do mediolitoral. As características morfológicas e ecológicas destas espécies que influenciam este padrão são discutidas.
BIOLOGICAL AND APPLIED SCIENCES
Recolonization of experimental gaps by the mussels Brachidontes darwinianus and B. solisianus in a subtropical rocky shore
Marcel Okamoto Tanaka * * Author for correspondence
Universidade Estadual de Campinas; Instituto de Biologia; Programa de Pós-Graduação em Ecologia; CP 6109; 13.083-970; Brazi.. Present address: Universidade Federal de Mato Grosso do Sul; CCBS; Departamento de Biologia; CP 549; 79.070-900; martnk@yahoo.com; Campo Grande - MS - Brazil
ABSTRACT
In this study, the recolonization speed of the mussels Brachidontes darwinianus and B. solisianus was compared to evaluate their responses to gaps opened by disturbances in the rocky shore. B. darwinianus was much faster than B. solisianus, rapidly closing gaps in the mussel bed. When they co-occurred, distribution patterns of these species were related not only with distinct physiological tolerances but also with responses to disturbance. B. darwinianus was more tolerant to low salinities, dominated estuarine zones generally occuring lower in the midlittoral, and presented a faster response to opened gaps.
Key words: Bivalve, Intertidal, Rocky shore, Zonation, Mytilidae
RESUMO
Mexilhões são importantes dominantes do espaço na região entremarés de costões rochosos. Diversos fatores influenciam a distribuição de espécies coexistentes, tais como diferenças de tolerância fisiológica, histórias de vida e uso de recursos distribuídos irregularmente no espaço e tempo. Neste estudo, a velocidade de recolonização dos mexilhões Brachidontes darwinianus e B. solisianus foi comparada para avaliar suas respostas a clareiras abertas por perturbações. B. darwinianus foi muito mais rápido que B. solisianus, fechando rapidamente clareiras abertas no banco de mexilhões. Este resultado sugere que quando estas espécies co-ocorrem, sua distribuição está relacionada não só a tolerâncias fisiológicas distintas, mas também a diferentes respostas a perturbações. B. darwinianus é mais tolerante a baixas salinidades, dominando zonas estuarinas, além de geralmente ocorrer na região mais baixa do mediolitoral. As características morfológicas e ecológicas destas espécies que influenciam este padrão são discutidas.
INTRODUCTION
Mussels are important components of rocky shore communities throughout the world, commonly dominating the space resource within the intertidal zone (Suchanek, 1985; Little and Kitching, 1996). Coexistence of different mussel species is related to distinct physiological tolerances, life-history attributes, and use of patchy resources in space and time (Harger, 1972; Levinton and Suchanek, 1978; Suchanek, 1981; Petersen, 1984; Nalesso et al., 1990; Iwasaki, 1994). In Brazilian rocky shores, two species of mytilids generally dominate the intertidal zone, Brachidontes darwinianus (Orbigny, 1846) and B. solisianus (Orbigny, 1846). B. darwinianus occurs from SE Brazil to the north of Patagonia, and is generally associated with sources of freshwater on the rocky shore and estuaries, whilst B. solisianus has a wider distribution on the western Atlantic, from Mexico to Uruguai, in a large range of environmental conditions (Klappenbach, 1965; Avelar and Narchi, 1984a; Rios, 1995).
When both Brachidontes species co-occur, they are spatially segregated: B. darwinianus is found in the lower midlittoral, whilst B. solisianus is found in the upper part; also, B. darwinianus replaces B. solisianus at sites subject to low salinities, such as estuaries (Avelar and Narchi, 1983; Nalesso et al., 1990). Earlier studies proposed that these distribution patterns could result from distinct tolerances of the species to desiccation and salinity, and to harsher conditions found in the lower intertidal (Avelar and Narchi, 1983, 1984a,b; Nalesso et al., 1990). Succession patterns in mixed species beds of Brachidontes indicated that B. darwinianus is a faster colonizer of gaps opened on emergent rocky substrate, rapidly depleting the space resource mainly by lateral migration (Tanaka and Magalhães, 2002). However, there is no information to evaluate whether interspecific effects are larger than intraspecific effects, or if both species simply differ on their movement abilities. The objective of this study was to evaluate the responses of both species to disturbance in exclusive beds to test the hypothesis that their movement rates differed; as a result, different recolonization patterns should be found. Also, the characteristics of both species and their relative effects on the structure of Brachidontes mussel beds is discussed.
MATERIALS AND METHODS
This study was carried out at Praia da Lagoinha (23º31'S, 45º11'W) in Ubatuba district, on the northern coast of São Paulo State. It is a semi-sheltered shore, with a small stream on its right side, where boulders of several sizes can be found. Mussels colonize the boulder surface, with dominance of B. darwinianus on boulders within the stream at the beach, and B. solisianus on those farther from the beach. In the intermediate area, both species co-occur forming exclusive zones. Mean air temperatures in the region vary between 19.5 - 25.9°C along the year, while mean monthly rainfall ranges between 82.6 - 247.6mm, with higher values of both variates in the summer, and the opposite in the winter; salinity values vary between 30-32. (Wainer et al., 1996).
A randomised blocks design was used to study the recolonization patterns of both species in exclusive beds. Four boulders (blocks) were randomly selected along a transect of 20m, and only boulders with similar-sized species were used. In each boulder, 25cm² quadrats in the center of each species' dominance zone were scrapped. These gaps were marked and monitored with an acrylic quadrat containing 16 points uniformely marked on it to estimate percent cover. Gaps were created in June 1996 and monitored monthly until November 1996. The size of these gaps and duration of the experiment were small enough to eliminate the importance of larval recruitment on succession patterns (Tanaka and Magalhães, 2002). Further, the rocky substrate was smooth and homogeneous, forming an inadequate substrate for larval attachment. The experiment was analysed with a repeated-measures ANOVA on arcsin-transformed percent cover values (Sokal and Rohlf, 1995). As the condition of sphericity was never observed, F-tests with the Greenhouse-Geiser correction were made (Winer, 1971).
RESULTS
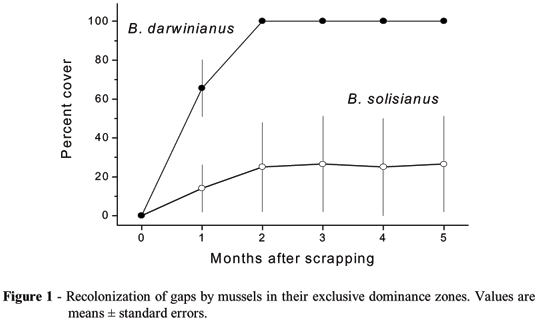
The spatial distribution of both Brachidontes species was stable throughout the experiment, with each species covering 100% of the substrate on their dominance zones. However, both species differed on their rates of lateral migration to occupy the gaps (Fig. 1).
Gaps opened in the B. darwinianus dominance zone closed in just two months, whereas those opened in the B. solisianus zone had mean cover of only 25% at the end of the experiment, as a result of adult movement. There were no juvenile mussels in the experimental gaps. Recolonization speed measured as mean area covered during a period of five months was significantly higher for B. darwinianus (Table 1).
DISCUSSION
Intertidal rocky shores in SE Brazil commonly present dominance zones formed by mussels. The composition and width of these zones vary both among shores and years, but patterns of variation and mussel population dynamics are not established. Brachidontes darwinianus and B. solisianus are dominant space occupiers in the midlittoral, although the former species is always associated with sources of freshwater (Nalesso et al., 1990). B. solisianus has a wider distribution and generally occurs in the upper midlittoral, although its dominance is commonly discontinuous, forming mosaics on the rocky substrate. This indicates that recruitment densities in B. solisianus beds are generally low, and there must be a limiting recruitment density, above which this species maintains itself in the community (Petersen et al., 1986).
Tanaka and Magalhães (2002) showed that in mixed Brachidontes beds bare rock represents about 20% of the available substrate, with predominance of gaps smaller than 50cm². In this situation, B. darwinianus rapidly occupied the gaps by lateral migration, dominating the mussel bed. The results of the present experiment indicated that B. darwinianus moved faster than B. solisianus, and the behavior of the latter species was not influenced by the former, at the temporal scale considered. This difference could be due to the distinct intertidal zones occupied by each species. As B. darwinianus occurs in the lower midlittoral, it is subject to greater wave impact and sand scouring, sometimes being covered by sand (Avelar and Narchi, 1983). In these conditions, the ability to move to more favourable positions could be important for species survival, influencing the distribution patterns of B. darwinianus (e.g., Davenport and Wilson, 1995). The colonization dynamics of the two species indicated that B. darwinianus presented a high turnover rate, exploring the space resource that was more frequently available in the lower intertidal, whilst B. solisianus was continuously dislodged from the mussel bed, eventually becoming locally extinct for some time. Dominance of the rocky shore by B. solisianus could only be recovered after a massive recruitment event (Petersen et al., 1986). However, the influence of mussel recruitment depends on the availability of attachment sites, formed mainly by secondary substrata such as algae, barnacles, and mussels, and crevices on the rock surfaces (Petersen, 1984). Thus, recruitment of mussels from the water column have more influence on larger denuded areas than the ones considered here, which provide more physical heterogeneity either through crevices on the rock surfaces or recruitment of species that can function as secondary substrata, as found in other rocky intertidal systems (e.g., Paine and Levin, 1981).
Thus, disturbances could also influence the distribution and zonation patterns of these mussels. B. darwinianus has lower tolerance to desiccation, higher tolerance to low salinities, and more sensitive cilia to cope with the larger amount of sediment found in the lower intertidal, enabling it to occupy both sites subject to lower salinities (e.g., estuaries) and lower positions in the intertidal zone (Table 2). Larger sizes could also help this species to maintain water exchange, even when partially buried by sand. Higher recolonization speed due to higher movement rates could help B. darwinianus to occupy areas more subject to disturbances and gaps opened in mixed beds, thus maintaining itself in the intertidal community. The interaction between B. darwinianus and B. solisianus in field conditions is still poorly understood, and future research is necessary to better understand the competitive abilities of both species, and the role of disturbances on their distribution.
ACKNOWLEDGEMENTS
I thank ALT Souza for discussions and help in the field, and CA Magalhães for discussions. This study received financial support from FAPESP (procs 95/2260-0 and 96/3733-1).
Received: January 22, 2003;
Revised: October 16, 2003;
Accepted: July 20, 2004.
- Avelar, W. E. P. (1980), Ciclo anual da gametogênese de Brachidontes darwinianus darwinianus (Orbigny, 1846) Bivalvia, Mytilidae. Rev. Nordest. Biol., 3, 151-177.
- Avelar, W. E. P. and Boleli, I. C. (1989), Annual cycle of gametogenesis in Brachidontes solisianus (Orbigny, 1846) (Mollusca, Bivalvia, Mytilidae). Iheringia Sér. Zool., 69, 9-18.
- Avelar, W. E. P. and Narchi, W. (1983), Behavioral aspects of Brachidontes darwinianus darwinianus (Orbigny, 1846) and Brachidontes solisianus (Orbigny, 1846) (Bivalvia, Mytilidae) in response to a salinity gradient. Iheringia Sér. Zool., 63, 125-132.
- Avelar, W. E. P. and Narchi, W. (1984a), Anatomia funcional de Brachidontes darwinianus darwinianus (Orbigny, 1846) (Mollusca: Bivalvia). Papéis Avulsos Zool., 35, 331-359.
- Avelar, W. E. P. and Narchi, W. (1984b), Functional anatomy of Brachidontes solisianus (Orbigny, 1846) (Bivalvia: Mytilidae). Bolm. Zool., 8, 215-237.
- Davenport, J. and Wilson, P. C. (1995), Mobility, gregariousness and attachment in four small bivalve mollusc species at Husvik, South Georgia. J. Moll. Stud., 61, 491-498.
- Harger, J. R. (1972), Competitive co-existence: maintenance of interacting associations of the sea mussels Mytilus edulis and Mytilus californianus. Veliger, 14, 387-410.
- Iwasaki, K. (1994), Distribution and bed structure of the two intertidal mussels, Septifer virgatus (Wiegmann) and Hormomya mutabilis (Gould). Publ. Seto Mar. Biol. Lab., 36, 223-247.
- Klappenbach, M. A. (1965), Lista preliminar de los Mytilidae brasileños com claves para su determinación y notas sobre su distribucion. An. Acad. Bras. Ciênc., 37 : (suppl.), 327-352.
- Levinton, J. S. and Suchanek, T. H. (1978), Geographic variation, niche breadth and genetic differentiation at different geographic scales in the mussels Mytilus californianus and M. edulis. Mar. Biol., 49, 363-375.
- Little, C. and Kitching, J. A. (1996), The biology of rocky shores Oxford University Press, New York.
- Nalesso, R. C., Duarte, L. F. L., and Mendes, E. G. (1990), Influência da salinidade e exposição ao ar na distribuição dos mexilhões Brachidontes darwinianus e B. solisianus em estuários do litoral do Estado de São Paulo. Anais do II Simpósio de Ecossistemas da Costa Sul e Sudeste Brasileiro, 3, 111-122.
- Paine, R. T. and Levin, S. A. (1981), Intertidal landscapes: disturbance and the dynamics of pattern. Ecol. Monogr., 51, 145-178.
- Petersen, J. H. (1984), Larval settlement behavior in competing species: Mytilus californianus Conrad and M. edulis L. J. Exp. Mar. Biol. Ecol., 82, 147-159.
- Petersen, J. A., Sutherland, J. P. and Ortega, S. (1986), Patch dynamics of mussel beds near São Sebastião (São Paulo), Brazil. Mar. Biol., 92, 389-393.
- Rios, E. C. (1995), Seashells of Brazil 2nd ed, Museu Oceanográfico da FURG, Rio Grande do Sul.
- Sokal, R. R. and Rohlf, F. J. (1995), Biometry 3rd ed. Freeman, New York.
- Suchanek, T. H. (1981), The role of disturbance in the evolution of life history strategies in the intertidal mussels Mytilus edulis and Mytilus californianus. Oecologia, 50, 143-152.
- Suchanek, T. H. (1985), Mussels and their rôle in structuring rocky shore communities. In The ecology of rocky coasts, eds. P. G. Moore and R. Seed. Hodder and Stoughton, London, pp. 70-96.
- Tanaka, M. O. and Magalhães, C. A. (1999), Morphometric species recognition in Brachidontes darwinianus and Brachidontes solisianus (Bivalvia: Mytilidae). Veliger, 42, 267-274.
- Tanaka, M. O. and Magalhães, C. A. (2002), Edge effects and succession dynamics in Brachidontes mussel beds. Mar. Ecol. Prog. Ser., 237, 151-158.
- Wainer, I. E. K. C., Colombo, P. M. and Miguel, A. J. (1996). Boletim de monitoramento climatológico para as bases Norte e "Dr João de Paiva Carvalho" do Instituto Oceanográfico da Universidade de São Paulo. Relat. Téc. Inst. Oceanogr., 38, 1-13.
- Winer, B. J. (1971), Statistical principles and experimental design 2nd ed. New York : McGraw-Hill.
Publication Dates
-
Publication in this collection
15 Apr 2005 -
Date of issue
Jan 2005
History
-
Received
22 Jan 2003 -
Reviewed
16 Oct 2003 -
Accepted
20 July 2004