Abstracts
Food allergy is an adverse reaction that occurs in susceptible people when they eat sensitizing foods and is one of the causes of Inflammatory Bowel Disease (IBD). The effort to understand the induction process of these diseases is important as IBD is increasing worldwide, including in Brazil. The aim of this study was to develop an experimental antigen specific inflammatory process of the gut of mice and rats, using peanut seeds. Animals were immunized with peanut protein extract before their exposure to the in natura peanut seeds. Results showed that systemic immunization with peanut protein extracts rendered significantly higher antibody titers than control groups and that immunized animals submitted to a challenge diet containing peanuts presented time dependent alterations of the gut similar to celiac disease. In conclusion, results suggested that this experimental model was a convenient tool to study the evolution of alterations in chronic antigen specific gut inflammatory process.
Food allergy; peanuts; inflammatory bowel disease; rats; mice
A alergia alimentar consiste em uma reação adversa que ocorre em pessoas susceptíveis quando ingerem alimentos sensibilizantes, sendo uma das causas das Doenças Inflamatórias Intestinais (IBD). O objetivo deste estudo foi desenvolver um protocolo experimental de indução de um processo inflamatório intestinal antígeno-específico em camundongos e ratos. Foi escolhida para a indução deste processo a semente de amendoim. Os animais foram imunizados com o extrato protéico previamente à exposição com a semente in natura. Nossos resultados mostram que a imunização sistêmica com extratos protéicos de amendoim ocasiona títulos significativamente maiores de anticorpos quando comparado ao grupo controle e que os animais imunizados submetidos ao desafio com a dieta contendo exclusivamente amendoim apresentam alterações intestinais tempo-dependente similares àquelas observadas na doença celíaca. Os resultados obtidos sugerem que este modelo experimental constitui uma ferramenta conveniente para avaliar as alterações no processo inflamatório intestinal crônico antígeno-específico
HUMAN AND ANIMAL HEALTH
Induction of an Antigen Specific Gut Inflammatory Reaction in Mice and Rats: a Model for human Inflammatory Bowel Disease
Gerlinde Agate Platais Brasil TeixeiraI,II; Danielle Mota Fontes AntunesI,III,* * Author for correspondence: danimfantunes@gmail.com ; Archimedes Barbosa de Castro JúniorI,II; Janilda Pacheco da CostaI,III; Patrícia Olaya PaschoalI,II; Sylvia Maria Nicolau CamposI,II; Monique de Morais Bitetti PedruzziI,II; Vinicius da Silva AraújoI; Luis Antônio Botelho AndradeII; Silvana Ramos Farias MorenoIII; Alberto Felix Antonio da NobregaIV; Gilberto Perez CardosoIII
IDepartamento de Imunobiologia; Laboratório do Grupo de Imunologia Gastrintestinal; Instituto de Biologia; Universidade Federal Fluminense; 24020-150; Niterói -RJ -Brasil
IIPrograma de Pós-Graduação em Patologia; Universidade Federal Fluminense; 24033-900; Niterói -RJ -Brasil
IIIPrograma de Pós-Graduação em Ciências Médicas; Centro de Ciências Médicas; Universidade Federal Fluminense; 24033-900; Niterói -RJ -Brasil
IVDepartamento de Imunologia; Instituto de Microbiologia e Imunologia; Universidade Federal do Rio de Janeiro; 21944-970; Ilha do Fundão -RJ -Brasil
ABSTRACT
Food allergy is an adverse reaction that occurs in susceptible people when they eat sensitizing foods and is one of the causes of Inflammatory Bowel Disease (IBD). The effort to understand the induction process of these diseases is important as IBD is increasing worldwide, including in Brazil. The aim of this study was to develop an experimental antigen specific inflammatory process of the gut of mice and rats, using peanut seeds. Animals were immunized with peanut protein extract before their exposure to the in natura peanut seeds. Results showed that systemic immunization with peanut protein extracts rendered significantly higher antibody titers than control groups and that immunized animals submitted to a challenge diet containing peanuts presented time dependent alterations of the gut similar to celiac disease. In conclusion, results suggested that this experimental model was a convenient tool to study the evolution of alterations in chronic antigen specific gut inflammatory process.
Key words: Food allergy, peanuts, inflammatory bowel disease, rats, mice
RESUMO
A alergia alimentar consiste em uma reação adversa que ocorre em pessoas susceptíveis quando ingerem alimentos sensibilizantes, sendo uma das causas das Doenças Inflamatórias Intestinais (IBD). O objetivo deste estudo foi desenvolver um protocolo experimental de indução de um processo inflamatório intestinal antígeno-específico em camundongos e ratos. Foi escolhida para a indução deste processo a semente de amendoim. Os animais foram imunizados com o extrato protéico previamente à exposição com a semente in natura. Nossos resultados mostram que a imunização sistêmica com extratos protéicos de amendoim ocasiona títulos significativamente maiores de anticorpos quando comparado ao grupo controle e que os animais imunizados submetidos ao desafio com a dieta contendo exclusivamente amendoim apresentam alterações intestinais tempo-dependente similares àquelas observadas na doença celíaca. Os resultados obtidos sugerem que este modelo experimental constitui uma ferramenta conveniente para avaliar as alterações no processo inflamatório intestinal crônico antígeno-específico.
INTRODUCTION
Contrary to what is commonly thought, the largest contact area of the body is not the skin, but the mucosal surface, which is bombarded immediately after birth by a large variety of microorganisms and protein antigens from the environment. The gut itself presents an area approximately 300 to 600 fold that of the skin (Brandtzaeg, 2002; Moog, 1981). In humans, the mucosa of the small intestine is estimated to be 300 m2 and it is constantly exposed to antigenic contacts. Approximately 30 kg of food proteins reaches this organ during a year and 130-190g of these proteins is absorbed daily in the gut (Brandtzaeg, 1998). This means that the principal source of perturbation of immunological activity occurs via the mucosa and events initiated at this site have important systemic reflections due to its large number of immune cells. Despite of this, most of the times that antigens penetrate the system through the gut, the resulting immunological response is not the classical Th1 or Th2 response (Weiner, 2001). This mucosal immune network has a unique aspect -the ability to maintain relative unresponsiveness (tolerance) to an enormous array of antigens (Ags) derived from dietary sources and commensal bacteria (Hyun et al., 2006). Immune responsiveness to these harmless agents must be prevented to maintain the integrity of the gut and allow nutrient absorption (Hyun et al., 2006). Therefore, immunological nonresponsiveness to luminal Ags is more common than mucosal responsiveness.
Another way of viewing this is that oral tolerance allows the mucosal immune system to focus on Ags or pathogens that pose a threat to the host (Faria and Weiner, 2005). The expression oral tolerance is used when a reduction in the systemic immunological response is observed after parentally challenging the animal with the same antigen that it has eaten (Weiner, 2001). The immunological consequence of the oral administration of an antigen normally depends on where and how the antigen is taken up and presented to T cells. The intestinal immune system is characterized by a distinct profile of cytokines (Signore et al., 2002), adhesion molecules, chemokines and cells (Mowat, 2003). The idea that the immunological environment of the gut seems to make great efforts to ensure that tolerance is the default response to antigen is often challenged on the basis that this would be a dangerous strategy for host survival in the face of continuous exposure to pathogens. The inappropriate immune response to foods and commensal bacteria that are responsible for celiac disease and Crohn's disease are due to deregulation of these crucial processes (Mowat, 2003). Thus, an increasing incidence of adverse reactions to ingested foods is being reported, which may reflect a failure of oral tolerance induction or a breakdown of its maintenance. Peanuts appear to consist of particular potent allergenic proteins and the prevalence of peanut allergy is rising in both children and adults, mainly in USA and UK (Strid et al., 2004). It is also known for its persistence through life as just 1020% of peanut allergic patient lose their sensitivity to peanut (Skolnick et al., 2001) compared with 85% for egg and cow's milk allergy (Dannaeus and Inganas, 1981; Host et al., 2002). Peanut (Arachis hypogaea) belongs to the family of Leguminosae and consists of albumins and storage proteins that comprise 87% of the total protein contents (Fries, 1982). Several peanut proteins have been identified as allergens, such as Ara h1, Ara h2, Ara h3 (Burks et al., 1998). However, recent work shows that the food matrix is important in the immune response to peanut and purified peanut allergens may have little intrinsic stimulatory capacity (van Wijk et al., 2005).
The gut-associated lymphoid tissue (GALT) which is responsible for the inductive phase of the immune response can be divided into effecter sites which consist of organized tissues such as the Peyer's patches and mesenteric lymph nodes (MLNs) and of lymphocytes scattered throughout the epithelium and lamina propria of the mucosa (Hamada et al., 2002). Many investigators believe that the lymphoid aggregates of T and B cells of the Peyer patches are the central players in the induction of the immune response after oral contact with macromolecules derived from the diet or flora (Bruce and Ferguson, 1986). In the gut, special sets of T and B cells co-exist, which suggests the existence of a special immunological reactivity, prepared for constant contacts of the immune system and the environment. On the contrary, Inflammatory Bowel Disease (IBD) is associated with classical Th1 and Th2 responses, which are characterized by the breakdown of the immunoregulatory mechanisms that maintain oral tolerance (Andrade et al., 2003). Crohn's disease and Ulcerative colitis both are IBD's with distinct immunoregulatory characteristics and clinical manifestations. They are given a common name because sometimes one resembles the other so closely that even a pathologist cannot distinguish between them (Karlinger et al., 2000). Many potential metabolic problems may complicate IBD, and this usually occurs in patients with severe forms of the disease (Allan, 1998). The study of animal models of mucosal inflammation as a means to probe the pathogenesis of IBD extends for almost a half century (Strober et al., 2002). Several animal models are available for use in IBD research which evaluates pharmacological molecules or agents that could lead to a possible cure for this devastating disease entity. Over the past two decades, a steadily increasing number of more than 20 experimental models with a variable range of clinical manifestations similar to those observed in human IBD have been developed (Jurjus et al., 2004). These models contributed greatly to important advances in our current understanding of the underlying mechanisms of inflammation and disease pathogenesis as well as treatment. In general, an appropriate or an optimal animal model should display certain key characteristics similar or identical to the human IBD course: pathophysiology, symptoms and signs. The gut of this model should also exhibit inflammation and morphological alterations (Jurjus et al., 2004). For this, most of the available models induce gut inflammation through irritation of the mucosal surface by their exposure to chemical reagents and products, such as acetic acid, phorbol ester and various sulfated polysaccharides (carageenan, amylopectin sulfate and dextran sulfate sodium) or use genetically modified animals (Strober et al., 2002).
In this work peanut seeds (Arachis hypogea) were used on the source of antigen as peanuts have been considered to be one of the most potent food allergens causing severe diseases. Although in humans, most of the clinical symptoms are Th2 related and show acute anaphylactic symptoms; here a model with a chronic profile is presented. The aim of this study was to develop an animal model of the antigen specific gut inflammation in rats and mice to analyze the serum, the gut histology and some clinical parameters (as weight measure and presence of diarrhea), comparing control with immunologically manipulated (immune) animals.
MATERIALS AND METHODS
Animals
In this study, 30 male adult Lou-M rats (8-12 weeks) and 30 female (6-8 weeks) BALB/c and C57BL/6J mice were used, bred and maintained at the Animal Facility of the Immunobiology Department of Fluminense Federal University (Rio de Janeiro, Brazil). The animals of each strain were randomly divided into two groups: control group and experimental group. They were individually numbered enabling paired statistical analysis. The Ethics Committee of the Medicine School of Fluminense Federal University approved this study (protocol 167/05).
Peanut protein extracts (PPE)
Peanut seeds were minced in an electrical coffee grinder sieved and defatted with ether (Teixeira et al., 2008). To the defatted seed preparation, extraction buffer (Sodium borate, 0,0125M, SDS 1%, Mercaptoetanol 1% -pH 10) was added, maintaining a 1:10 (w/v) ratio. This mixture was then placed on a rocker for 30 min at room temperature and centrifuged at 600g for 30 minutes. The supernatant was collected and kept frozen at -20ºC until use. The protein concentration was determined using the method of Lowry et al. (1951).
Immunization protocol for rats
The rats were immunized twice with 200µg PPE subcutaneously (sc). Primary immunization was done with 5mg Al(OH)3 and booster immunization without adjuvant after a 21-28 day interval. Control animals were sham immunized with physiological saline plus 5mg Al(OH)3, and booster immunization without adjuvant.
Immunization protocol for mice
The mice were immunized twice with 100µg PPE sc. Primary immunization was done with 1mg Al(OH)3 and booster immunization without adjuvant after a 21-28 day interval. Control animals were sham immunized with physiological saline plus 1mg Al(OH)3, and booster immunization without adjuvant.
Challenge diet
A challenge diet composed exclusively of peanut seeds in natura offered ad libitum for a 30-day period was introduced one week after secondary immunization.
Bleeding
All the animals were bled from the retrorbital plexus prior to manipulation and 10 days after each immunization withdrawing 1ml and 200µl from rats and mice, respectively. The serum was collected and stored at -20º C until analyses.
Clinical Evaluation
Weekly clinical evaluation included: weight measure (Plenna digital balance 0.1g resolution), presence of diarrhea and fur characteristics.
ELISA
The serum samples were evaluated by ELISA method to quantify specific anti-peanut protein antibody titers. For antigen-specific IgG and IgG1 antibodies, 96-well plates were coated with PPE at 10µg/well in Phosphate Buffered Saline (PBS) at 4ºC overnight. Plates were washed with 0,05% Tween-20 in PBS and then blocked with 100µl of a 1% PBS/gelatin solution for 1h at room temperature. Serum samples were plated in 1:100 dilution (100µl/well) and incubated for 3h at room temperature. Plates were then washed and received peroxidase-labeled rabbit anti-mouse IgG (SIGMA-Aldrich -Germany), goat anti-mouse IgG1 (Southern Biothechnology - USA) or goat anti rat IgG (Southern Biothechnology - USA) and incubated for 3h at room temperature. Reactions were developed with 100µl of solution containing H2O2 and o-phenylene-diamine (OPD) (SIGMA-Aldrich -Germany). Plates were read at 492nm on an automated ELISA reader (Anthos 2010, Germany). The results are reported as arbitrary units of ELISA corresponding to the area under the dilution curve of each serum.
Collection of the Intestine Segments and Histological Parameters
After the diet exposure period, all animals were euthanazed to collect 1-3cm segments of gastroduodenum junction and jejunum. These segments were fixed with 10% buffered formaldehyde and stained with Hematoxilin-Eosin (HE). The histological parameters evaluated from both duodenum and jejunum were the general description of the slide (integrity of the intestinal structure, number of villi per field, edema, congestion and leukocyte infiltrate). The villi height/width ratio; epithelial-cell-height/laminapropriety-width ratio, villi-height/cript-height ratio and Intestinal Epithelial Cells/Intraepithelial Leukocytes (IEC/IEL) ratio were obtained from the previous parameters.
Statistical Analysis
Statistical analysis were performed by using Fisher's test, ANOVA and Tukey´s post test to determine the minimum significance difference (MSD) using GraphPad InStat program by GraphPad Software Inc®.
RESULTS
Weight analysis
The weight of all the rats and mice was analyzed during the peanut feeding period (challenge diet).
In rats, at the beginning of the challenge diet, animals of both groups (control and experimental) presented an average weight of 250.75±12.10g and 258.80±16.21g, respectively. After the 4th week of the challenge diet, it was observed that control animals presented a significant difference of weight compared with experimental rats (p<0.05). Control rats gained 19.55±0.24g while experimental animals lost an average of 5.62±2.82g. Animals of the control group gained weight during the first week of the challenge diet while experimental animals began to lose weight at the second week (Fig. 1A).
At the beginning of the challenge diet, mice, of both groups (control and experimental) presented an average weight of 23.81g ± 1.47g. Mice of both strains presented a similar behavior. Experimental group began to lose weight at the first week of the challenge diet with an average loss of 1.42±0.45g, while the control group began to gain weight at the second week and presented an average of 1.48±0.37g. The weight difference between the groups was significant (p<0.05) (Fig. 1B).
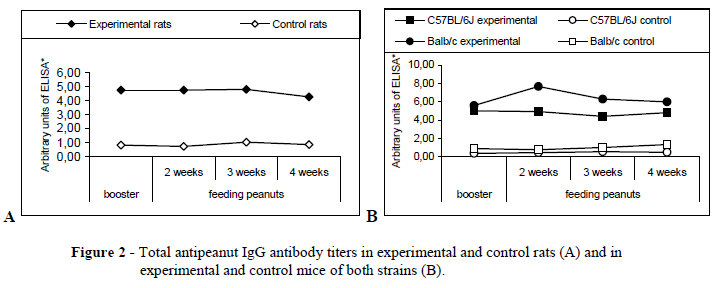
The systemic immunization with crude peanut extract renders significantly higher antibody titers
Experimental rats (submitted to systemic immunization with crude peanut extract) rendered significantly higher total IgG antibody titers (4.651±0.180 units) than control rats (sham immunized) (0.879±0.087 units) (p<0.05); eating peanuts over a 30-day period, after immunization, maintained the high antibody levels of experimental animals (Fig. 2A).
Similar to rats, mice of both strains, submitted to the systemic immunization with crude peanut extract, presented higher titers of IgG and IgG1 when compared to control counterparts. There were no differences between the experimental BALB/c and C57BL/6J IgG1 antibodies titers, although total specific IgG titers of experimental BALB/c mice were significantly higher than of C57BL/6J mice (C57BL/6J: 5.084±0.376 units; BALB/c: 7.673±3.360 units; p<0.05) (Fig. 2B). As seen with the rats, eating peanuts over a 30-day period, after immunization, maintained the high antibody levels of experimental mice (in contrast to control animals that did not present the observed increase in antibody titers). The difference of both strains IgG and IgG1antibody titers between control and experimental mice was significant (p<0.05).
Morphological Analysis of the Intestine
The macroscopic analysis of the rats and mice revealed a frail consistency of the intestinal tissue pertaining to animals of the experimental group in contrast to the intestinal tissue of the control group.
The microscopic analysis agreed with the macroscopy, in which a preserved intestinal structure in control animals was seen while in the experimental animals their villi presented edema, congestion and a high leukocyte infiltration in both duodenum and jejunum (Fig. 3 e 4).
In rats (Fig. 3), the villi from duodenum but not jejunum of experimental animals presented significant alterations (p<0.05) in the height/width ratio (3.54±1.10) when compared to control animals (4.55±0.74). A significant difference (p<0.05) in jejunum but not duodenum was observed in experimental animals when evaluated the epithelial-cell-height/lamina-propria-width ratio (0.62±0.08), in which the control animals presented a higher ratio (0.83±0.38). The third criteria analyzed, villi-height/crypt-height ratio, also showed significant difference (p<0.05) in only one of the intestinal segments, duodenum but not jejunum, in which this ratio was higher in control animals (1.97±0.11) when compared to the experimental animals (1.80±0.07). The fourth criteria used to determine the inflammatory process was to compare the number of Intestinal Epithelial Cells (IEC) and Intraepithelial Leukocytes (IEL) and establish the IEC/IEL ratio. For this criterion, both the segments analyzed presented significant differences when compared to control animals (experimental duodenum: 9.09±1.43 vs. control duodenum: 22.95±2.93 p<0.05 and experimental jejunum: 8.52±2.43 vs. control jejunum: 13.22±0.80; p<0.05), in which control animals presented a higher ratio than the experimental animals. The sera Ab titers and the IEC/IEL ratio of control and experimental animals were correlated and according to Fisher's test, the row/column association was statistically significant and the two-sided p value was 0.0236.
Histological evaluation of mice (Fig. 4) showed that experimental BALB/c mice developed signs of gut inflammation in the second week of peanut challenge, whereas in the experimental C57BL/6J mice only presented significant inflammatory signs on the third week of challenge. The presence of edema in lamina propria and loss of vilous integrity in duodenum was observed.
Control animals submitted to peanut diet presented IEC/IEL ratio (9.80±0.97) similar to that observed in chow fed animals either (11.00±1.55).
BALB/c experimental mice presented a significant increase (p<0.05) in intraepithelial leukocytes number in duodenal villus, represented by the decrease of IEC/IEL ratio (5.60±1.14), and lamina propria edema in the second week after receiving the challenge diet (peanut). The IEC/IEL ratio, in these animals, persisted until the fourth week of peanut challenge (5.72±1.11). Experimental C57BL/6J animals only presented a similar decrease in IEC/IEL ration in the the third week of peanut challenge (4.46±1.07). As observed with BALB/c mice, the IEC/IEL ratio in duodenum of C57BL/6J animals did not change after its initial reduction (4.73±0.64). The peanut diet caused gut morphological alterations (p<0.05) in the experimental mice of both strains tested and was characterized by vilous atrophy and lamina propria edema, especially after the fourth week of challenge. Finally, control animals, which underwent peanut consumption, presented normal histological appearance of gut as seen in chow fed animals.
DISCUSSION
The present work demonstrated the reproducibility of a peanut allergy model, originally developed in C57BL/6J mice by Teireixa (2003), in BALB/c mice and Lou-M rats. The features of this model of gut inflammation were vilous atrophy, crypt hiperplasia, prominent mononuclear leukocyte infiltrate and lamina propria edema. These signs were typical of human celiac disease (gluten enteropathy), which has been also observed in other food induced enteropathies (Marsh, 1995). Teixeira (2003) demonstrated that experimental the C57BL/6J mice presented a mild weight loss but no visible diarrhea. A weight loss was observed in rats and BALB/c mice in the experimental group with a gain in weight in all the control animals suggesting that eating a foodstuff rich in calories in normal situations induced fattening while in the presence of an enteropathy, this same food induced weight loss.
In other works, authors have been using intragastric challenge to achieve an allergic reaction (Morafo et al., 2003; Adel-Patient et al., 2005). Nevertheless, the subcutaneous mode of immunization may not be so far from real conditions of primary sensitization. Recently, Strid et al.(2005) have demonstrated that epicutaneous exposure to peanut proteins led to allergic sensitization and prevented oral tolerance, suggesting that epidermal sensitization could be realistic to provide a food allergy condition.
The presence of high and similar titers of IgG1 a Th2 derived IgG isotype, in both C57BL/6J and BALB/c mice in the present model of peanut allergy deserved more attention as well as the fact that no major difference between the gut inflammatory patterns were found into these two mouse strains. Although IgG subtypes in rats were not measured, the results obtained for total specific IgG was similar to that of the mice.
In contrast to the current thought that peanut allergy is primarily a Th2-driven process, there are accumulating data from the literature indicating that peanut allergy is a result of a mixed Th1-Th2 immune response. Van Wijk and colleagues (2004) demonstrated a mixed citokine response and polyisotypic humoral response in murine model of peanut allergy, specifically to major peanut allergens Ara h1, Ara h2, Ara h3 and Ara h6. Both IL-4 and IFN-γ production as well as IgE, IgG1 and IgG2a were found at the very early phase of the response to peanuts. The same group also examined the role of cytotoxic T-lymphocyte associated antigen 4 (CTLA-4), a negative regulator of T cell activation, on peanut sensitization (Van Wijk et al., 2005b). The use of an antibody against CTLA-4 in a murine model of peanut allergy failed to elicit an allergen-specific response. Despite the induction of Th2 cytokines, activation of antigen presenting cell (APC) and an elevation in total serum IgE levels, anti-CTLA-4 treatment did not induce peanut-specific IgE antibodies or mast cell degranulation. These data challenge the pivotal role of a Th2 cytokine environment in the induction phase of peanut allergy.
In the present work, although Th2 derived antibody isotype (IgG1) was found in mice, the cellular infiltrate of the gut epithelium was not typical of an anaphylactic profile. In addition, the absence of anaphylactic systemic reactions in the model corroborated the idea of a mixed Th1-Th2 presence of peanut allergy. Finally, the verification of similar inflammatory response in the gut of rats and mice with different genetic backgrounds validated this model of peanut allergy and provided an interesting way for the study of this mixed immune response to peanut proteins. Our group has been interested in the study of natural products derived from local plants as possible alternatives for anti-inflammatory drugs (Moreno et al., 2005). We are now conducting new experiments in which this animal model shall be used.
In conclusion, results confirmed that systemic immunization to food antigens could make animals susceptible to challenge diets, leading to an antigen specific inflammatory bowel disease, what would make this a convenient tool to study the evolution of alterations in chronic antigen specific gut inflammatory process. As these findings were similar for more than one species, it could be argued that this was a robust model for food allergy.
ACKNOWLEDGEMENTS
This study was supported by CAPES and UFF. We would also like to thank Ms. Maira Platais for her significant contribution in the revision of the language.
Received: December 27, 2006; Revised: July 10, 2007; Accepted: July 09, 2008.
- Adel-Patient K., Bernard H., Ah-Leung S., Créminon C. and Wal J.M. (2005), Peanut and cow's milkspecific IgE, Th2 cells and local anaphylactic reaction are induced in Balb/c mice orally sensitized with cholera toxin. Allergy, 60, 658-64.
- Allan, R.N. (1998), Metabolic problems in inflammatory bowel disease. Neth. J. Med., 53, S9-12.
- Andrade, M.C.; Vaz, N.M. and Faria, A.M. (2003), Ethanol-induced colitis prevents oral tolerance induction in mice. Braz. J. Med. Biol. Res., 36, 1227-1232.
- Brandtzaeg, P. (1998), Development and basic mechanisms of human gut immunity. Nutr. Rev., 56, S5-S18.
- Brandtzaeg, P. (2002), Current understanding of gastrointestinal immunoregulation and its relation to food allergy. Ann. N.Y. Acad. Sci., 964, 13-45.
- Bruce M.G. and Ferguson, A. (1986), Oral tolerance to ovalbumin in mice: studies of chemically modified and 'biologically filtered' antigen. Immunol., 57, 627-630.
- Burks, W.; Sampson, H.A. and Bannon, G.A. (1998), Peanut allergens. Allergy, 53, 725-730.
- Dannaeus, A. and Inganas, M. (1981), A follow-up study of children with food allergy. Clinical course in relation to serum IgE-and IgG-antibody levels to milk, egg and fish. Clin. Allergy, 11, 533-539.
- Faria, A.C. and Weiner, H.L. (2005), Oral tolerance. Immunol. Rev., 206, 232-59.
- Fries, J.H. (1982), Peanuts: allergic and other untoward reactions. Ann. Allergy, 48, 220-226.
- Hamada, H. et al.(2002), Identification of multiple isolated lymphoid follicles on the antimesenteric wall of the mouse small intestine. J. Immunol., 168, 57-64.
- Host, A.; Halken, S.; Jacobsen, H.P.; Christensen, A.E.; Herskind, A.M. and Plesner, K. (2002), Clinical course of cow's milk protein allergy/intolerance and atopic diseases in childhood. Pediatr. Allergy Immunol., 13, Suppl 15, 23-28.
- Hyun, J.G. and Barrett, T.A. (2006), Oral tolerance therapy in inflammatory bowel disease. Am. J. Gastroenterol., 101, 569-571.
- Jurjus, A.R.; Khoury, N.N. and Reimund, J.M. (2004), Animal models of inflammatory bowel disease. J. Pharmacol. Toxicol. Meth., 50, 81-92.
- Karlinger, K.; Gyorke, T.; Mako, E.; Mester, A. and Tarjan, Z. (2000), The epidemiology and the pathogenesis of inflammatory bowel disease. Eur. J. Rad., 35, 154-167.
- Lowry, Q.H. et al (1951), Protein measurement with folin phenol reagent. Biol. Chem., 193, 265-275.
- Marsh, M.N. (1995), Intestinal manifestations of food hypersensitivity. In: Intestinal Immunology and Food Allergy New York: Raven Press, 34, 57-71.
- Moog, F. (1981), The lining of the small intestine. Scient Amer., 245, 116-125.
- Morafo, V. et al.(2003), Genetic susceptibility to food allergy is linked to differential TH2-TH1 responses in C3H/HeJ and BALB/c mice. J. Allergy Clin. Immunol., 111, 1122-1128.
- Moreno S. R. F. et al (2005), Bioavailability of the Sodium Pertechnetate and Morphometry of Organs Isolated from Rats: Study of Possible Pharmacokinetic Interactions of a Ginkgo Biloba Extract. Braz. Arch. Biol. Technol., 48-Special, 73-78.
- Mowat, A.McI. (2003), Anatomical basis of tolerance and immunity to intestinal antigens. Nat. Rev. Immunol., 3, 331-341.
- Signore A.; D`Alessandria C.; Annovazzi A. and Scopinaro F. (2002), Radiolabelled Cytokines for Imaging Chronic Inflammation. Braz. Arch. Biol. Technol., 45-Special: 15-23.
- Skolnick, H.S.; Conover-Walker, M.K.; Koerner, C.B.; Sampson, H.A.; Burks, W. and Wood, R.A. (2001), The natural history of peanut allergy. J. Allergy Clin. Immunol., 107, 367-374.
- Strid, J.; Thomson, M.; Hourihane, J.; Kimber, I. and Strobel, S. (2004), A novel model of sensitization and oral tolerance to peanut protein. Immunol., 113, 293-303.
- Strid, J.; Hourihane, J.; Kimber, I.; Callard, R. and Strobel, S. (2005), Epicutaneous exposure to peanut protein prevents oral tolerance and enhances allergic sensitization. Clin. Exp. Allergy, 35, 757-766.
- Strober, W.; Fuss, I.J.; Blumberg, R.S. (2002), The immunology of mucosal models of inflammation. Annu. Rev. Immunol., 20, 495-549.
- Teixeira, G.A.P.B. (2003), Um modelo murino de inflamação intestinal crônica -tolerância e imunização oral: antigenicidade versus temporalidade, PhD Thesis, Fluminense Federal University, Niterói, RJ, Brazil.
- Teixeira, G.A.P.B.; Paschoal, P.O.; Oliveira, V.L.; Pedruzzi, M.M.B.; Campos S.M.N.; Andrade L. and Nóbrega A. (2008), Diet selection in immunologically manipulated mice. Immunobiology, 213(1), 1-12.
- Van Wijk, F.; Hartgring, S.; Koppelman, S.J.; Pieters, R. and Knippels, L.M. (2004), Mixed antibody and T cell responses to peanut and the peanut allergens Ara h1, Ara h2, Ara h3 and Ara h6 in an oral sensitization model. Clin. Exp. Allergy, 34, 1422-1428.
- Van Wijk, F.; Hoeks, S.; Nierkens, S.; Koppelman, S.J.; Van Kooten, P.; Boon, L.; Knippels, L.M. and Pieters, R. (2005), CTLA-4 signaling regulates the intensity of hypersensitivity responses to food antigens, but is not decisive in the induction of sensitization. J. Immunol., 174, 174-179.
- Van Wijk, F.; Nierkens, S.; Hassing, I.; Feijen, M.; Koppelman, S.J.; de Jong, G.A.; Pieters, R. and Knippels, L.M. (2005b), The effect of the food matrix on in vivo immune responses to purified peanut allergens. Toxicol. Sci., 86, 333-341.
- Weiner, H.L. (2001), Induction and mechanism of action of transforming growth factor-beta-secreting th3 regulatory cells. Immunol. Rev., 182, 207-214.
Publication Dates
-
Publication in this collection
22 July 2009 -
Date of issue
June 2009
History
-
Accepted
09 July 2008 -
Reviewed
10 July 2007 -
Received
27 Dec 2006