Abstracts
This paper describes a protocol for the efficient vegetative propagation of Cleome rosea by somatic embryogenesis. Leaf and stem explants from nursery-grown seedlings of C. rosea were cultivated on Murashige and Skoog (MS) medium supplemented with indole-3-acetic acid (IAA), a -naphthaleneacetic acid (NAA), 4-amino-3,5,6-trichloropicolinic acid (picloram) or 2,4-dichlorophenoxyacetic acid (2,4-D). Nodular calli were produced from both explant types in the presence of 4.5 and 9.0 µM 2,4-D. Embryo development and maturation were achieved when calli from stem explants were transferred to media containing a ten-fold reduction of 2,4-D concentration initially used (0.45 and 0.90 µM). Leaf-derived calli did not form embryos with the same treatments. The highest frequency of embryogenic callus formation (85%) and number of embryo per callus (13.45 ± 2.8) were achieved during the first subculture on medium supplemented with 0.90 µM 2,4-D. Embryo conversion into plantlets was achieved following transfer to growth regulator-free MS medium solidified with 2 g.L-1 phytagel. An acclimatization rate of 53% was found three months after transfer to ex vitro conditions and the recovered plants presented a normal phenotypic aspect.
auxin; embriogenic callus; histology; phytagel; somatic embryos
O trabalho descreve um protocolo para a propagação in vitro de Cleome rosea por embriogênese somática. Explantes foliares e caulinares, obtidos de plantas germinadas sob condições in vivo, foram cultivados em meio de Murashige and Skoog (MS) suplementado com ácido 3-indolacético (AIA), ácido naftalenoacético (ANA), ácido 4-amino-3,5,6-tricloropicolínico (picloram) ou ácido 2,4-diclorofenoxiacético (2,4-D). Calos de aspecto nodular foram produzidos a partir de ambos os tipos de explante na presença de 4,5 e 9,0 μM de 2,4-D. O desenvolvimento e a maturação de embriões somáticos foram alcançados quando calos obtidos de explantes caulinares foram transferidos para meio de cultura suplementado com uma concentração de 2,4-D dez vezes menor do que aquelas utilizadas na indução do processo de calogênese (0,45 e 0,90 μM). Calos derivados de explantes foliares não produziram embriões ao serem submetidos a estes mesmos tratamentos. Os maiores valores de freqüência de calos embriogênicos (85%) e número médio de embriões por calo (13,45±2,8) foram alcançados durante a primeira subcultura em meio suplementado com 0,90 μM de 2,4-D. O processo de conversão dos embriões somáticos em plantas foi observado após transferência dos embriões para meio MS sem suplementação hormonal solidificado com 2 g.L-1 de fitagel. Três meses após a transferência para condições ex vitro a taxa de aclimatização alcançada foi de 53% e as plantas apresentavam um aspecto fenotípico normal.
BIOLOGICAL AND APPLIED SCIENCES
Somatic embryogenesis and plant regeneration from callus cultures of Cleome rosea Vahl
Claudia SimõesI,* * Author for correspondence: labplan_uerj@yahoo.com.br ; Norma AlbarelloI; Cátia Henriques CalladoII; Tatiana Carvalho de CastroI; Elisabeth MansurI
INúcleo de Biotecnologia Vegetal; Universidade do Estado do Rio de Janeiro; Rio de Janeiro - RJ - Brasil
IILaboratório de Anatomia Vegetal; Universidade do Estado do Rio de Janeiro; Rua São Francisco Xavier, 524; 20550-013; Rio de Janeiro - RJ - Brasil
ABSTRACT
This paper describes a protocol for the efficient vegetative propagation of Cleome rosea by somatic embryogenesis. Leaf and stem explants from nursery-grown seedlings of C. rosea were cultivated on Murashige and Skoog (MS) medium supplemented with indole-3-acetic acid (IAA), a -naphthaleneacetic acid (NAA), 4-amino-3,5,6-trichloropicolinic acid (picloram) or 2,4-dichlorophenoxyacetic acid (2,4-D). Nodular calli were produced from both explant types in the presence of 4.5 and 9.0 µM 2,4-D. Embryo development and maturation were achieved when calli from stem explants were transferred to media containing a ten-fold reduction of 2,4-D concentration initially used (0.45 and 0.90 µM). Leaf-derived calli did not form embryos with the same treatments. The highest frequency of embryogenic callus formation (85%) and number of embryo per callus (13.45 ± 2.8) were achieved during the first subculture on medium supplemented with 0.90 µM 2,4-D. Embryo conversion into plantlets was achieved following transfer to growth regulator-free MS medium solidified with 2 g.L-1 phytagel. An acclimatization rate of 53% was found three months after transfer to ex vitro conditions and the recovered plants presented a normal phenotypic aspect.
Key words: auxin; embriogenic callus; histology; phytagel; somatic embryos
RESUMO
O trabalho descreve um protocolo para a propagação in vitro de Cleome rosea por embriogênese somática. Explantes foliares e caulinares, obtidos de plantas germinadas sob condições in vivo, foram cultivados em meio de Murashige and Skoog (MS) suplementado com ácido 3-indolacético (AIA), ácido naftalenoacético (ANA), ácido 4-amino-3,5,6-tricloropicolínico (picloram) ou ácido 2,4-diclorofenoxiacético (2,4-D). Calos de aspecto nodular foram produzidos a partir de ambos os tipos de explante na presença de 4,5 e 9,0 μM de 2,4-D. O desenvolvimento e a maturação de embriões somáticos foram alcançados quando calos obtidos de explantes caulinares foram transferidos para meio de cultura suplementado com uma concentração de 2,4-D dez vezes menor do que aquelas utilizadas na indução do processo de calogênese (0,45 e 0,90 μM). Calos derivados de explantes foliares não produziram embriões ao serem submetidos a estes mesmos tratamentos. Os maiores valores de freqüência de calos embriogênicos (85%) e número médio de embriões por calo (13,45±2,8) foram alcançados durante a primeira subcultura em meio suplementado com 0,90 μM de 2,4-D. O processo de conversão dos embriões somáticos em plantas foi observado após transferência dos embriões para meio MS sem suplementação hormonal solidificado com 2 g.L-1 de fitagel. Três meses após a transferência para condições ex vitro a taxa de aclimatização alcançada foi de 53% e as plantas apresentavam um aspecto fenotípico normal.
INTRODUCTION
The higher plants constitute one of the most important sources of major bioactive compounds and the world market for medicinal plants has been estimated on US $ 60 billion per year (Kartal, 2007). However, the indiscriminate extraction and overexploitation have caused a significant reduction in natural populations. Furthermore, environmental impact of human action in all the ecosystems results in the extinction of a great number of plant species not yet studied with regard to their pharmacological activities.
Cleome rosea is a Brazilian herbaceous annual species whose medicinal potential has been indicated in studies using plants collected from natural populations, as well as in vitro plants (Simões et al., 2006). It is frequently found in sandy coastal plains (restingas), ecosystems intensely affected by human occupation for housing, tourism and land speculation, resulting in serious environmental damages (Zamith and Scarano, 2006). This species is also employed as garden ornamental due to its pink and attractive inflorescences.
Considering the rapid depletion of important natural wealth, the search for effective multiplications methods has assumed added significance in conservation and reintroduction programs. This is especially interesting for medicinal plants, in order to allow the supply of botanical material with uniform quality for biotechnological, pharmacological and phytochemical studies. In this context, plant cell and tissue culture technologies, which include the induction of organogenesis and somatic embryogenesis, present many advantages over the conventional methods, such as independence of geographical and seasonal variations and rapid production (Ramachandra Rao and Ravishankar, 2002).
Somatic embryogenesis is an important system for vegetative propagation, whereby somatic cells can generate embryos due to the totipotency of plant cells, returning to a dedifferentiated state that is redetermined towards the embryogenic pathway. This process provides a useful experimental model to investigate the events of plant embryogenesis and is also one of the most important means for mass propagation of elite or genetically modified plants (Santos et al., 2002).
Although in vitro regeneration has been achieved through organogenesis in some Cleome species (Nassen and Jha, 1994; 1997; Simões et al., 2004; Albarello et al., 2006), somatic embryogenesis has not been reported on the genus.
The present work was undertaken to study the effect of different explant types and plant growth regulators on induction, maturation and conversion of somatic embryos of C. rosea.
MATERIALS AND METHODS
Induction of embryogenic calluses
Two month-old nursery-grown seedlings were washed with triclosan detergent (1% v/v) and rinsed in running tap water, followed by the removal of the roots. The aerial part was immersed for 10 min in NaOCl (0.5% v/v) plus Tween 80 (0.05% v/v) under agitation (100 rpm) and rinsed three times with sterile distilled water (5 min each). Segments from stems (0.5 cm) and leaves (0.5 cm2) were excised and cultured during 30 days on MS medium (Murashige and Skoog, 1962), containing 30 g.L-1 sucrose and supplemented with the auxins indole-3-acetic acid (IAA) at 5.7 and 11.4 µM, a-naphthaleneacetic acid (NAA) at 5.4 and 10.7 µM, 4-amino-3,5,6-trichloropicolinic acid (picloram) at 4.1 and 8.3 µM or 2,4-dichlorophenoxyacetic acid (2,4-D) at 4.5 and 9.0 µM. Media were adjusted to pH 5.8 prior to adding agar (8 g.L-1, Merck), autoclaved (121º C, 104 KPa) for 15 min and dispensed into 8 x 7 cm flasks (30 mL per flask) closed with polypropylene caps. Cultures were incubated in a growth chamber under 16-h photoperiod provided by cool-white fluorescent tubes (45 µmol m-2s-1) at 26 ± 2º C. Four explants were inoculated into each flask with a total of 40 explants per treatment.
Development, maturation and conversion of somatic embryos
To promote somatic embryo (SE) development and maturation, calli were transferred to MS medium as follows: a) growth regulator-free; b) supplemented with 1.8 µM 6-furfurylaminopurine (kinetin); c) similar auxin composition used to promote callus induction; d) ten-fold reduction in the auxin concentration used to promote callus induction. The cultures were kept under the same physical conditions described above. Subcultures into fresh medium were performed at 20-day intervals. At each subculture, the SE developed on callus surfaces were isolated and transferred to growth regulator-free full or half-strength (MS1/2) MS medium solidified with 8 g.L-1 agar (Merck) or 2 g.L-1 phytagel (Sigma) to promote germination (conversion stage). A total of 46 embryos were used per treatment on conversion assays and the results were expressed as the percentage of whole plants or other morphogenetic responses obtained.
Histological studies
Embryogenic calli and SE were fixed in FAA 50, dehydrated in a graded ethanol series and embedded in paraffin (Johansen, 1940). Serial sections (10 µm) were double-stained in astra blue-basic fuchsin. Photographs were obtained with the image capture system Image-Pro Plus for Windows using a video camera Optronics (Olympus, German) attached to an Olympus BX40 microscope (Olympus, German).
Acclimatization
The plantlets were transferred to plastic pots (7.5 x 7.0 cm) containing a mixture of garden soil and sand (2:1). The pots were placed into glass chambers (80 cm x 40 cm x 40 cm) at 28±2º C under a 12-h photoperiod for 30 days. In order to reduce the relative humidity inside the chambers, the covers were gradually opened after the second week and completely removed 30 days after transplanting. Plant survival was evaluated after three months.
Statistical analysis
Data were analyzed using one-way analysis of variance (ANOVA) and the means were compared by Tukey test. The statistical analysis was performed at the 5% level of significance using the MSTATC statistical software package.
RESULTS AND DISCUSSION
The morphogenetic responses of C. rosea callus cultures varied according to the treatment employed. Embryogenic, rhizogenic and non-embryogenic callus lines were obtained. Media supplemented with IAA did not cause any morphogenetic response in leaf explants, while inducing the development of shoots in stem segments. On the other hand, media containing NAA stimulated rhizogenesis in both explant types. In callus cultures from shoot apices of Crataeva nurvala, another Capparaceae species, the use of NAA promoted a low calogenic growth, whereas the supplementation with IAA induced root differentiation (Inamdar et al., 1990).
The presence of 2,4-D or picloram induced the development of friable light beige calli both from leaf and stem explants. The calogenic process began within the first week of culture and a positive correlation between growth regulator concentration and callus yield was observed (data not shown). When these calli were transferred to MS supplemented with 1.8 µM kinetin or to growth regulator-free MS medium, only small white roots with dense root hairs were formed. On the other hand, KIN associated with 2,4-D was effective in inducing SE in callus cultures of C. nurvala (Inamdar et al., 1990).
Calli maintained on media with the same concentrations of 2,4-D or picloram used to induce calogenesis, showed a continuous callus growth. However, embryo development and maturation were achieved when calli derived from the stem explants in response to 4.5 and 9.0 µM 2,4-D were transferred to media supplemented with 10-fold reduced concentration of this growth regulator. On the other hand, picloram-induced calli did not originate embryos when transferred to the same treatment. The ability of 2,4-D in triggering the embryogenic pathway may be related to its capacity to induce stress genes, which have been shown to contribute to the cellular reprogramming of somatic cells toward embryogenesis (Kitamiya et al., 2000).
The acquisition of embryogenic competence by somatic cells must involve reprogramming of gene expression patterns as well as changes in the morphology, physiology and metabolism. In addition, a high level of endogenous auxin is considered as one of the crucial signals determining embryogenic competence (Fehér et al., 2003; Gaj, 2004) and in most species further development of SE usually requires reduction of auxin level (Jiménez, 2005), as observed in the present work. However, in some species, both stages were achieved in the same medium (Inamdar et al., 1990; Magioli et al., 2001; Park et al., 2005).
In contrast to calli obtained from stem explants, leaf-derived calli did not follow an embryogenic route when transferred to medium with a reduction on 2,4-D concentration. This variation between the responses displayed by different explant types could be attributed to the physiological state of the donor organs, mainly related to the level of endogenous phytohormones and sensitivity to auxins due to variations in the number of receptors or receptor affinity (Jiménez, 2005).
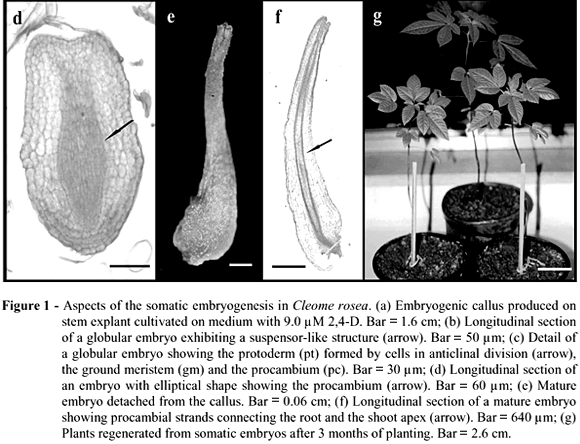
Calli induced by 2,4-D presented a nodular appearance (Fig. 1a), which is considered a typical physical feature of embryogenic calli. However, this correlation between appearance and embryogenic competence must be used carefully, since calli that present the same features observed in C. rosea, like rough surface, friable texture and pale coloration, are considered as non embryogenic (Quiroz-Figueroa et al., 2006).

Calli transferred to media supplemented with 0.45 or 0.90 µM 2,4-D showed an asynchronous and continuous differentiation of SE. Embryos at the globular stage were observed 7-9 days after transfer to media with a reduced 2,4-D concentration and some of them presented a proliferation of cells near the callus tissue, suggesting the presence of a suspensor-like structure (Fig. 1b). Although suspensor-like structures are not frequently reported (Quiroz-Figueroa et al., 2002), their presence could be considered as an indication of the unicellular origin of somatic embryos (Nonohay et al., 1999). The occurrence of both multicellular and unicellular origins of SE, as observed in the present work, is considered a common phenomenon, although the multicellular origin is more likely to occur in indirect somatic embryogenesis (Fernando et al., 2001; Gaj, 2004).
The globular embryos presented a protoderm formed by cells dividing in anticlinal plane (Fig. 1c). The protoderm is considered one of the unique features of SE development and could regulate the embryogenic process by applying physical and cell division limitations (Sharma et al., 2004). The ground meristem under the protoderm was characterized by a pronounced cell vacuolization and narrow cells with dense cytoplasm formed the procambium at the central core. Embryos with an elliptical shape, which could be recognized as an intermediary stage between the heart and torpedo stages were visualized (Fig. 1d). Quiroz-Figueroa et al. (2002) also reported the presence of transition stages between the globular and heart stages, as well as between the heart and torpedo stages in the indirect somatic embryogenesis system of Coffea arabica.
Mature embryos loosely attached to the callus surface were observed 15 days after transfer to medium with reduced 2,4-D concentration (Fig. 1e). These embryos showed a bipolar structure with the presence of procambial strands connecting the root and the shoot apices and with no vascular connection to the callus tissue (Fig. 1f). The frequency of embryogenic callus formation as well as the mean number of SE produced per callus were significantly higher on medium supplemented with 0.90 µM 2,4-D (Table 1). A gradual reduction in the embryogenic competence was observed after the second subculture, probably due to the prolonged culture on medium containing 2,4-D. In order to explain reduced embryogenic competence in sweet potato callus lines during continuous exposure to 2,4-D, Padmanabhan et al. (2001) suggested that this process was related to a decrease in auxin-responsive gene expression correlated with ageing of the culture. Changes in cellular competence to form embryos in the presence of 2,4-D were also observed in sugarcane callus culture (Guiderdoni and Demarly, 1988). In addition, Myers and Simon (1999) reported a decrease in the organogenic potential and an increase in genetic instability in callus cultures of Allium sativum maintained for extended periods of time on medium supplemented with this growth regulator.
The conversion stage, which is characterized by the germination of the SE and development of plantlets, demonstrating their physiological and morphological maturity, was achieved on growth regulator-free MS medium. Embryo conversion efficiency was influenced both by salt concentration and type of gelling agent (Table 2).
Media solidified with agar only induced the development of roots or callus at the basal end of some embryos. The highest rate of whole plantlets formation (41.3%) was achieved on full-strength MS medium solidified with phytagel. Saito and Nishimura (1994) and Magioli et al. (2001) also observed highest conversion rates of SE on media solidified with phytagel, as compared to those solidified with agar. The use of phytagel could reduce water availability in the culture medium and contribute to desiccation (Klimaszewska et al., 2000; Choudhury et al., 2008), similarly to the natural process that occurs in most zygotic embryos (Bradford, 1994). Reduced water availability seems to modulate embryo metabolism inducing the synthesis of proteins and enzymes found in germinating seeds and is considered by some authors as the most important factor to improve germination of SE (Malabadi et al., 2004).
In all treatments applied to induce the conversion step, some SE did not present any morphogenic response and some of them showed high oxidation. The lack of SE conversion might be due to morphological abnormalities as a consequence of the long exposure to 2,4-D (Raghavan, 2004). Such abnormalities could result from defective polar transport of endogenous auxins in response of the exogenous growth regulator (Choi et al., 1997). Insufficient storage products in the cotyledons could also result in lack of shoot development from SE (Chengalrayan et al., 2001). Nevertheless, considering that in some species normal plants can be recovered from abnormal embryos, it is possible that these embryos obtained from C. rosea also could germinate and develop normally. Thus, in spite of the high conversion rate obtained on growth regulator-free MS medium solidified with phytagel, the established protocol could be further optimized to improve the conversion capacity.
An acclimatization rate of 53% was found three months after the transfer of plantlets to ex vitro conditions. Plants presented a normal phenotypical aspect (Fig. 1g), flowered after eight months and produced fruits. The seeds were collected and germinated in nursery conditions originating a second generation of plants with normal features. The present work reached its objective in establishing a methodology for propagation of C. rosea through somatic embryogenesis, showing the ability of 2,4-D to induce this process from stem explants. The system described here represents an alternative approach for clonal propagation and provides a basis for further studies on somatic embryogenesis in Cleome species.
ACKNOWLEDGEMENTS
The authors are grateful to Jeanne A. T. Glória (Universidade do Estado do Rio de Janeiro) for the valuable technical assistance and Dr Claudia F. Barros (Instituto de Pesquisa Jardim Botânico do Rio de Janeiro) for the use of Image Capture System. This work was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Received: October 24, 2008; Revised: February 17, 2009; Accepted: October 21, 2009.
- Albarello, N.; Simões, C.; Rosas, P.F.G.; Castro, T.C.; Gianfaldoni, M.G.; Callado, C.H.; Mansur, E. (2006), In vitro propagation of Cleome spinosa (Capparaceae) using explants from nursery-grown seedlings and axenic plants. In Vitro Cell Dev Biol Plant., 42, 601-606
- Bradford, K.J. (1994), Water stress and the water relations of seed development: a critical review. Crop Sci., 34, 1-11
- Chengalrayan, K.; Hazra, S.; Gallo-Meagher, M. (2001), Histological analysis of somatic embryogenesis and organogenesis induced from mature zygotic embryo-derived leaflets of peanut (Arachis hypogaea L.). Plant Sci., 161, 415-421
- Choi, Y.E.; Kim, H.S.; Soh, W.Y.; Yang, D.C. (1997), Development and structural aspects of somatic embryos formed on medium containing 2,3,5-triiodobenzoic acid. Plant Cell Rep., 16, 738-744
- Choudhury, H.; Kumaria, S.; Tandon, T. (2008), Induction and maturation of somatic embryos from intact megagametophyte explants in Khasi pine (Pinus kesiya Royle ex. Gord.). Curr. Sci., 95 (10), 1433-1438
- Fehér, A.; Pasternak, T.P.; Dudits, D. (2003), Transition of somatic plant cells to an embryogenic state. Plant Cell Tiss. Org. Cult., 74, 201-228
- Fernando, J.A.; Melo, M.; Soares, M.K.M.; Appezzato-da-Glória, B. (2001), Anatomy of somatic embryogenesis of Carica papaya L. Braz. Arch. Biol. Technol., 45 (3), 277-286
- Gaj, M.D. (2004), Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul., 43, 27-47
- Guiderdoni, E.; Demarly, Y. (1988), Histology of somatic embryogenesis in cultured leaf segments of sugarcane plantlets. Plant Cell Tiss. Org. Cult., 14, 71-88
- Guzzo, F.; Baldan, B.; Mariani, P.; Lo Schiavo, F.; Terzi, M. (1994), Studies on the origin of totipotency cells in explants of Daucus carota L. J. Exp. Bot., 45, 1427-1432
- Inamdar, J.A.; Nataraj, M.; Mohan, J.S.S.; Subramanian, R.B. (1990), Somatic embryogenesis from callus cultures of Crataeva nurvala - Buch. Ham. Phytomorphology, 40, 319-322
- Jiménez, V.M. (2005), Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul., 47, 91-110
- Johansen, S. (1940), Plant Microtechnique. Mc Graw-Hill Book Company, New York
- Kartal, M. (2007), Intellectual property protection in the natural product drug discovery, traditional herbal medicine and herbal medicinal products. Phytother. Res. 21, 113-119
- Kitamiya, E.; Suzuki, S.; Sano, T.; Nagata, T. (2000), Isolation of two genes that were induced upon the initiation of somatic embryogenesis on carrot hypocotyls by high concentrations of 2,4-D. Plant Cell Rep., 19, 551-557
- Klimaszewska, K.; Bernier-Cardou, M.; Cyr, D.R.; Sutton, B.C.S. (2000), Influence of gelling agents on culture medium gel strength, water availability. tissue water potential and maturation response in embryogenesis cultures of Pinus strobus L. In Vitro Cell. Dev. Biol. - Plant, 36, 279-286
- Magioli, C.; Rocha, A.P.M.; Tarré, E.; Santiago-Fernandes, L.D.; Oliveira, D.E.; Krul, W.R.; Mansur, E. (2001), Effect of morphological factors, antibiotics and Agrobacterium co-cultivation in the efficiency of somatic embryogenesis of eggplant (Solanum melogenta L). J. Plant Biotech., 3, 19-25
- Malabadi, R.B.; Choudhury, H.; Tandon, P. (2004), Initiation, maintenance and maturation of somatic embryos from thin apical dome sections in Pinus kesiya (Royle ex. Gord) promoted by partial desiccation and gellan gum. Scientia Horticult., 102, 449-459
- Murashige, T.; Skoog, F. (1962), A revised medium for rapid growth and biossays with tobacco tissue cultures. Physiol. Plant., 15, 473-497
- Myers, J.M.; Simon, P.W. (1999), Regeneration of garlic callus as affected by clonal variation, plant growth regulators and culture conditions over time. Plant Cell Rep., 19, 32-36
- Naseem, M.; Jha, K. K. (1994), Differentiation and regeneration in Cleome leaves culture in vitro. Egypt J Bot.,1, 37-49
- Naseem, M.; Jha, K. K. (1997), Rapid clonal multiplication of Cleome gynandra DC, through tissue culture. Phytomorphology, 47 (4), 405-411
- Nonohay, J.S.; Mariath, J.E.A.; Winge, H. (1999), Histological analysis of somatic embryogenesis in Brazilian cultivars of barley, Hordeum vulgare vulgare, Poaceae. Plant Cell Rep., 18, 929-934
- Padmanabhan, K.; Cantliffe, D.J.; Koch, K.E. (2001), Auxin-regulated gene expression and embryogenic competence in callus cultures of sweet potato, Ipomoea batatas (L.) Lam. Plant Cell Rep., 20, 187-192
- Park, S-Y.; Ahn, J-K.; Lee, W-Y.; Murthy, H.N.; Paek, K-Y. (2005), Mass production of Eleutherococcus koreanum plantlets via somatic embryogenesis from root cultures and accumulation of eleutherosides in regenerants. Plant Sci., 168, 1221-1225
- Quiroz-Figueroa, F.R.; Fuentes-Cerda, C.F.J.; Rojas-Herrera, R.; Loyola-Vargas, V.M. (2002), Histological studies on the developmental stages and differentiation of two different somatic embryogenesis systems of coffea arabica Plant Cell Rep., 20, 1141-1149
- Quiroz-Figueroa, F.R.; Rojas-Herrera, R.; Galaz-Avalos, R.M.; Loyola-Vargas, V.M. (2006), Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tiss. Organ Cult., 86, 285-301
- Raghavan, V. (2004), Role of 2,4-dichlorophenoxy acetic acid (2,4-D) on cultured zygotic embryos of Arabidopsis: cell expansion, cell cycling and morphogenesis during continuous exposure of embryos to 2,4-D. Am. J. Bot., 91 (11), 1743-1756
- Ramachandra Rao, S.; Ravishankar, G.A. (2002), Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv., 20, 101-153
- Saito, T.; Nishimura, S. (1994), Improved culture conditions for somatic embryogenesis using an aseptic ventilative filter in eggplant (Solanum melongena L). Plant Sci., 102, 205-211
- Santos, A.L.W.; Silveira, V.; Steiner, N.; Vidor, M.; Guerra, M.P. (2002), Somatic embryogenesis in parana pine (Araucaria angustifolia (Bert.) O. Kuntze). Braz. Arch. Biol. Technol., 45 (1), 97-106
- Simões, C.; Santos, A.S.; Albarello, N.; Figueiredo, S.F.L. (2004), Shoot organogenesis and plantlet regeneration from stem explants of Cleome rosea Vahl (Capparaceae). J. Plant Biotech., 6 (3), 199-204
- Simões, C.; Mattos, J.C.P.; Sabino, K.C.C.; Caldeira-de-Araújo, A.; Coelho, M.G.P.; Albarello, N.; Figueiredo, S.F.L. (2006), Medicinal potencial from in vivo and acclimatized plants of Cleome rosea Vahl ex DC. (Capparaceae). Fitoterapia, 77, 94-99
- Sharma, S.K.; Millam, S. (2004), Somatic embryogenesis in Solanum tuberosum L.: a histological examination of key developmental stages. Plant Cell Rep., 23, 115-119
- Zamith, L.R.; Scarano, F.R. (2006), Restoration of a restinga sandy coastal plain in Brazil: Survival and growth of planted woody species. Restoration Ecol., 14 (1), 87-94
Publication Dates
-
Publication in this collection
12 Aug 2010 -
Date of issue
June 2010
History
-
Reviewed
17 Feb 2009 -
Received
24 Oct 2008 -
Accepted
21 Oct 2009