Abstract
The aim of this work was to study the effect of some nutritional and environmental factors on the production of cellulases, in particular endoglucanase (CMCase) and exoglucanases (FPase) from Bacillus licheniformis MVS1 and Bacillus sp. MVS3 isolated from an Indian hot spring. The characterization study indicated that the optimum pH and temperature value was 6.5 to 7.0 and 50-55°C, respectively. Maximum cellulases production by both the isolates was detected after 60 h incubation period using wheat and rice straw. The combination of inorganic and organic nitrogen source was suitable for cellulases production. Overall, FPase production was much higher than CMCase production by both of the strains. Between the two thermophiles, the cellulolytic activity was more in B.licheniformis MVS1 than Bacillus sp. MVS3 in varying environmental and nutritional conditions.
Isolation; Thermophiles; Submerged fermentation; Parameter optimization; Cellulases; Hot spring
AGRICULTURE, AGRIBUSINESS AND BIOTECHNOLOGY
Optimization of fermentation conditions for cellulases production by Bacillus licheniformis MVS1 and Bacillus sp. MVS3 isolated from Indian hot spring
Somen Acharya; Anita Chaudhary
Division of Environmental Sciences; Indian Agricultural Research Institute; New Delhi-110012, India, E-mail: someniari@gmail.com
ABSTRACT
The aim of this work was to study the effect of some nutritional and environmental factors on the production of cellulases, in particular endoglucanase (CMCase) and exoglucanases (FPase) from Bacillus licheniformis MVS1 and Bacillus sp. MVS3 isolated from an Indian hot spring. The characterization study indicated that the optimum pH and temperature value was 6.5 to 7.0 and 50-55°C, respectively. Maximum cellulases production by both the isolates was detected after 60 h incubation period using wheat and rice straw. The combination of inorganic and organic nitrogen source was suitable for cellulases production. Overall, FPase production was much higher than CMCase production by both of the strains. Between the two thermophiles, the cellulolytic activity was more in B.licheniformis MVS1 than Bacillus sp. MVS3 in varying environmental and nutritional conditions.
Key words: Isolation, Thermophiles, Submerged fermentation, Parameter optimization, Cellulases, Hot spring
INTRODUCTION
Cellulose, the most abundant organic source of feed, fuel and chemicals (Spano et al. 1975) consists of glucose units linked by β-1,4-glycosidic bonds in a linear mode (Heck et al. 2002). Each year photosynthetic fixation of CO2 yields about 1011 tons of dry plant material worldwide, and almost half of this material consists of cellulose (Eriksson et al. 1990). Although large quantities of presently available cellulosic materials have great potential as a source of renewable energy, it is often considered as a source of voluminous waste (Abdel and Ismail 1995). Cellulases are the complex enzyme systems that hydrolyze the β-1,4 glycosidic bonds in the cellulose to release glucose units (Nishida et al. 2007). The cellulosic enzymes required for the hydrolysis of cellulose include endoglucanases (CMCase), exoglucanases (FPase) and β-glucosidases (cellobiase) (Matsui et al. 2000). In the current industrial processes, cellulolytic enzymes have many useful applications in textile, paper, detergent industries and increasing the utilization efficiency of plant materials in animal feed manufacture (Dienes et al. 2004; Duan et al. 2004). Currently, the production of these enzymes is mostly from Trichoderma and Aspergillus (Bhat 2000). However, a major disadvantage for industrial applications is that most cellulases from the fungal origin lack thermal stability at high temperature. Since most industrial processes are carried out at high temperatures, there is a clear need for thermophilic enzymes. Thermophilic cellulolytic enzymes have been generally isolated from the thermophiles and hyperthermophiles (Huang 2004). Cellulases produced by bacteria are often more effective catalysts. They may also be less inhibited by the presence of material that has already been hydrolyzed (feedback inhibition). In general, bacterial cellulases are constitutively produced, whereas fungal cellulases are inducible in nature (Suto and Tomita 2001). Thermophilic bacterial cellulases have been reported from Bacillus sp. (Mawadza et al. 2000); B. stearothermophilus and B. licheniformis (Hala and Priest 1994). One of the natural habitats of the thermophilic bacteria is the hot or thermal spring. Vajreshwari is one of the hot springs in India which has not been yet explored in details from the microbiological point of view. Since, the aim of this work was to identify effective thermophilic cellulase producers from the hot spring to study the cultivation conditions for optimum yield of cellulases.
MATERIALS AND METHODS
Enrichment, Isolation and Screening of Cellulases Producing Thermophilic Bacteria
Water samples were obtained from the Vajreshwari hot spring (19° 29′ 12″ N and 73° 1′ 33″ E), Maharashtra, India. Water samples were collected aerobically in 500 ml sterile plastic bottles which were filled, sealed and stored at -20°C temperature. For enrichment of the water samples for the isolation of cellulases producing thermophilic bacteria, the basic mineral liquid media (g/l): KH2PO4: 1.36; (NH4)2SO4: 1.0; MgSO4.7H2O: 0.2; FeSO4: 0.001; NaCl: 2.0; yeast extract, 1.0 (Patel et al. 2006) supplemented with 0.3% carboxymethyl cellulose (CMC), was added to the water sample (1:1), mixed and incubated at 50°C for 72 h. The samples were then diluted in sterile distilled water, plated on agar medium consisting of basal medium as above and the plates were incubated at 50°C. After growth was obtained in the agar plates, morphologically distinct colonies were sub- cultured on respective agar plates to get the pure colonies. The screening for high cellulases producers was done by the blood red cellulose stain method (Ranoa et al. 2005). Out of the seven isolates, two isolates having colonies with high diffusible zones were screened as potential cellulases producers for further study. These two isolates were preserved on the same agar medium at 4°C with periodic sub-culturing.
Bacterial Identification through Biochemical Tests and 16S rDNA Gene Sequences
Identification of the isolates were confirmed through Gram staining methods, by a series of biochemical tests as prescribed by Bergey manual (Bergey 1957) and through 16S rDNA gene sequences. For 16S rDNA gene sequence analysis, bacterial genomic DNA was extracted and purified (Minamisawa et al. 1992). 16S rDNA was sequenced and the 16S rDNA gene sequence of the isolates were aligned with reference 16S rDNA sequences of the GenBank using the BLAST algorithm available in NCBI for the identification of the isolates and found to be Bacillus licheniformis and Bacillus sp. The partial 16S rDNA sequence of the isolates has been submitted to GenBank. The accession number for B. licheniformis MVS1 is GU590781.
Cellulase Activity Measurements
The organisms were inoculated in Patel's medium supplemented with 0.3% CMC for 12 h at 50°C for preparation of pre-inoculum. It was further used to inoculate the 100 ml of Patel's medium for cellulase production. Cellulase activities were measured for the cells grown in 100 ml cellulolytic medium, supplemented with either 0.3% CMC or 1% pretreated wheat and rice straw as substrate in 500 ml Erlenmeyer flasks on a incubator shaker at 150 rpm. The flasks were incubated for about 40 h using CMC as substrate and about 80 h when wheat and rice straw were used as substrates. The aliquots drawn from triplicate flasks were centrifuged at 10,000 rpm for 10 minutes and cell-free supernatants were used for different cellulolytic enzyme assays. Rice and wheat straw obtained from the Indian Agricultural Research Institute's (IARI) experimental field were dried at 50°C, grounded to fine powder and passed through 30-mesh sieve. They were pretreated with 4% NaOH for overnight at room temperature and then washed and dried before use as substrates at 1% level for cellulase production.
Optimization of Parameters for Improving Cellulase Production
The most suitable pH of the fermentation medium was determined by adjusting the pH of the culture medium at different levels in the range of pH 5.5-7.5. In order to determine the effective temperature for cellulase production by the selected strain, fermentation was carried out at 5°C intervals in the range of 45 to 65°C. The fermentation was carried out up to 80 h, and the production rate measured at 20 h intervals. To detect the appropriate nitrogen source for cellulase production by the isolates, the fermentation medium was supplemented with two inorganic (ammonium sulphate and sodium nitrate) and two organic (yeast extract and beef extract) nitrogen compounds at 0.2% level, thereby substituting the prescribed nitrogen source of the fermentation medium.
Enzyme Assays
All enzyme assays were carried out in 50 mM sodium citrate buffer (pH 4.8) unless otherwise stated. CMCase activity was determined in accordance with the Ghose's procedure (Ghose 1987), with 1% solution of CMC as substrate. The release of reducing sugars in 30 min at 50°C was measured by the DNSA method (Miller 1959). FPase activity was assayed (Mandels et al. 1976) in a manner similar to that used to determine CMCase activity, by taking 50 mg of Whatman No. 1 filter paper in 50 mM sodium citrate buffer (pH 4.8) as the substrate and the concentration of glucose released by enzyme was estimated by DNSA method. A unit of activity was defined as the amount of enzyme required to liberate 1μmol of glucose per minute under the assay conditions.
Statistical Analysis
The data were statistically analyzed by Duncan's multiple range test at the 0.05 probability level (p <0.05) using SPSS statistical software (SPSS for Windows, Release 12).
RESULTS AND DISCUSSION
In the present experiment, the two cellulolytic hot spring isolates were identified as B. licheniformis MVS1 (GU590781) and Bacillus sp. MVS3. Biochemical test results of the isolated bacteria are shown in Table 1.
Effect of Different Nitrogen Sources on Cellulase Production
The results of the effect of various nitrogen sources on cellulase production by B. licheniformis MVS1 and Bacillus sp. MVS3 is illustrated in Table 2. Among all the nitrogen sources tested, in case of B. licheniformis MVS1, beef extract gave maximum FPase activity (0.542 IU/ml) when wheat straw was used as sole carbon source but in case of rice straw, control, i.e., cellulolytic medium was found to be better (0.471 IU/ml) (Table 2).
But for Bacillus sp. MVS3, the nitrogen source in the control was better over other treatments for both CMCase and FPase production for both wheat and rice straw. The present results showed lower cellulase activity with inorganic nitrogen sources which suggested that reduced utilization of inorganic nitrogen by aerobic bacteria. These data were in accordance with the results of Ray et al. (2007) who reported that organic nitrogen sources were more suitable for optimizing the cellulase production by B. subtilis and B. circulans than inorganic sources. On the contrary, Spiridonov and Wilson (1998) found that NH4 compounds were the most favourable nitrogen sources for cellulase synthesis.
Effect of pH on Cellulase Production
From the Table 3, it could be seen that the CMCase and FPase production were best at slightly acidic range of pH (pH 6.5) for B. licheniformis MVS1 but for Bacillus sp.
MVS3 FPase production was optimum at pH 7.0. There was decreased production of enzymes at alkaline range of pH. Most microorganisms grow optimally within a wide pH range. Immanuel et al. (2006) reported that the cellulolytic enzyme, endoglucanase from Cellulomonas, Bacillus, and Micrococcus sp., isolated from the estuarine coir netting effluents hydrolyzes substrate in the pH range of 4.0 to 9.0, with maximum activity at pH 7.0. Contrary to that, Song et al. (1985) observed optimal cellulase production at pH 9.0 by Clostridium acetobutylium.
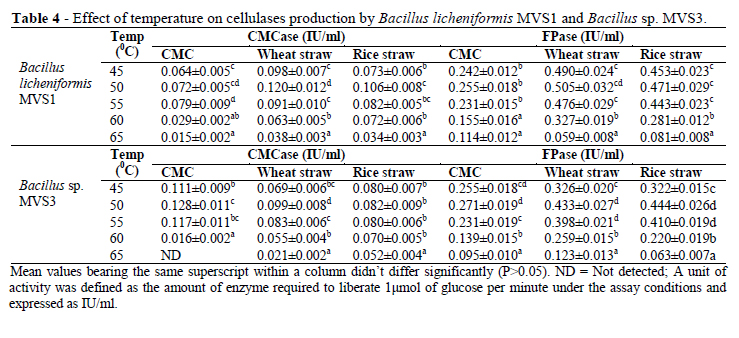
Effect of Temperature on Cellulase Production
Like pH, temperature is one of the most important parameters essential for the success of a fermentation reaction. For cellulase production by the two isolates, 50°C was most effective (Table 4). The production started declining after further increase in temperature and beyond 60°C, it declined significantly (p<0.05) (Table 4).
Souichiro et al. (2004) also reported the optimum temperature for the growth and cellulose degradation by Clostridium straminisolvens as 50-55°C. Immanuel et al. (2006) recorded maximum endoglucanase activity in Cellulomonas, Bacillus and Micrococcus sp. at 40°C at neutral pH.
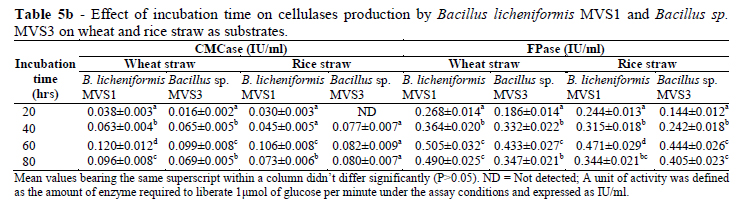
Effect of Incubation Period on Cellulase Production
The effect of incubation time on enzyme production was studied from 8-48 h (Table 5a) and 20-80 h (Table 5b) using CMC and rice and wheat straw as substrate, respectively. The production increased with increase in fermentation period and for B. licheniformis MVS1, CMCase production reached maximum (0.079 IU, 0.106 IU and 0.120 IU) at 8 h and 60 h after incubation when CMC, rice and wheat straw were used as substrates, respectively.
For Bacillus sp. MVS3, the trend was similar with B. licheniformis MVS1 (0.128 IU, 0.082 IU and 0.099 IU, respectively). Further increase in the incubation period resulted in the decreased production of CMCase. But FPase production reached maximum after 16 and 24 h of incubation period for B. licheniformis MVS1 and Bacillus sp. MVS3, respectively when grown on CMC and 60 h when grown on agricultural wastes as substrates.
At the same incubation period (24 h), Ariffin et al. (2006) recorded much lesser FPase (0.011 IU/ml) and CMCase (0.079IU/ml) activities by B. pumilus EB3. Decrease in enzymatic activity with time might be due to the depletion of nutrients and production of other by-products in the fermentation medium (Haq et al. 2005). Pothiraj et al. (2006) recorded maximum FPase activity (0.46 IU/ml) after 10 days of incubation period by Rhizopus stolonifer on cassava waste. The production of cellulases at comparatively earlier stages of fermentation for the isolated strains suggested the usefulness of these strains for enzyme production. Comparison of CMCase and FPase yield by the two isolates indicated most effectiveness of wheat straw as compared to rice straw and CMC treatment.
CONCLUSIONS
In conclusion, it could be said that hot springs appeared to be the potential natural source of thermophilic cellulolytic bacteria. The present study suggested that the nutritional and environmental factors significantly affected the cellulase production. The results also showed the possibility of wheat and rice straw as cheap source substrates for large-scale production of cellulase enzyme.
ACKNOWLEDGEMENT
The financial assistance provided by Indian Agricultural Research Institute (IARI) during the research is gratefully acknowledged.
Received: November 03, 2010;
Revised: November 21, 2011;
Accepted: March 05, 2012
- Abdel FAF, Ismail AM. Utilization of water hyacinth cellulose for production of cellulases by Trichoderma viride-100. Cytobios. 1995; 82(330): 151-7.
- Ariffin H, Abdullah N, Umi Kalsom MS, Shirai Y, Hassan MA. Production and characterisation of cellulase by Bacillus pumilus EB3. Int J Eng Technol. 2006; 3(1): 47-53.
- Bergey D. Manual of Determinative Bacteriology. American Society for Microbiology. Williams & Willkins Co. Publishers, Baltimore, USA. 1957; VII Edition.
- Bhat MK. Cellulases and related enzymes in biotechnology. Biotechnol Adv. 2000; 18: 355-383.
- Dienes D, Egyházi A, Réczey K. Treatment of recycled fiber with Trichoderma cellulases. 2004; Ind Crop Prod. 20: 11-21.
- Duan XY, Liu SY, Zhang WC. Volumetric productivity improvement for endoglucanase of Trichoderma pseudokoingii S-38. J Appl Microbiol. 2004; 96: 772-6.
- Eriksson K, Blanchette RA, Ander P. Microbial and enzymatic degradation of wood and wood components. Berlin: Springer-Verlag; 1990.
- Ghose TK. Measurement of cellulase activities. Pure Appl Chem. 1987; 59: 257-68.
- Hala KH, Priest FG. Thermotolerant varieties of Bacillus lichenifomis isolated from desert environments. J Appl Bacteriol. 1994; 77: 392-400.
- Haq IU, Hameed K, Shahzadi MM, Javed SA, Qadeer MA. Cotton Saccharifying activity of cellulases by Trichoderma harzianum UM-11 in shake flask. Int J Bot. 2005; 1: 19-22.
- Heck JX, Hertz PF, Ayub MAZ. Cellulase and xylanase production by isolated Amazon Bacillus strains using soya been industrial residue based solid-state cultivation. Braz J Microbiol. 2002; 33: 213-218.
- Huang XP. Purification and characterization of a cellulase (CMCase) from a newly isolated thermophilic aerobic bacterium Caldibacillus cellulovorans gen. nov. sp. World J Microbiol Biotechnol. 2004; 20: 85-92.
- Immanuel G, Dhanusha R, Prema P, Palavesam A. Effect of different growth parameters on endoglucanase enzyme activity by bacteria isolated from coir retting effluents of estuarine environment. Int J Env Sci Technol. 2006; 3: 25-34.
- Mandels M, Andreotti R, Roche C. Measurement of saccharifyng cellulase. Biotechnol Biogen Symp. 1976; 6: 21-31.
- Matsui I, Sakai Y, Matsui E, Kikuchi H, Kawarabayasi Y, Honda K. Novel substrate specificity of a membrane-bound β-glycosidase from the hyperthermophilic archaeon Pyrococcus horikoshi. FEBS Lett. 2000; 467: 195-200.
- Mawadza C, Hatti-Kaul R, Zvauya R, Mattiasson B. Purification and characterization of cellulases produced by two Bacillus strains. J Biotechnol. 2000; 83(3): 177-187.
- Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar, Analytical Chem. 1959; 31: 426-8.
- Minamisawa K, Takayuki S, Shingo O, Masatsugu K, Teruo A. Genetic relatedness of Bradyrhizobium japonicum field isolates as revealed by repeated sequences and various other characteristics. Appl Envi Microbiol. 1992; 58(9): 2832-9.
- Nishida Y, Suzuki KI, Kumagai Y, Tanaka H, Inoue A, Ojima T. Isolation and primary structure of a cellulase from the Japanese sea urchin Strongylocentrotus nudus. Biochimie. 2007; 1-10.
- Patel MA, Mark SO, Roberta H, Henry CA, Marian LB, Lonnie OI et al. Isolation and characterization of acid tolerant, thermophilic bacteria for effective fermentation of biomass-derived sugars to lactic acid. Appl Env Microbiol. 2006; 72(5): 3228-3235.
- Pothiraj C, Balaji P, Eyini M. Enhanced production of cellulases by various fungal cultures in solid state fermentation of cassava waste. Afr J Biotechnol. 2006; 5(20): 1882-5.
- Ranoa DRE, Lumibao CY, Roxas JL, San Luis BB, Hedreyda CT. Comparison of conventional plate assays with DNA-based screening protocols for protease and cellulase production from putative Bacillus isolates. Sci Diliman. 2005; 17(1): 31-6.
- Ray AK, Bairagi A, Ghosh KS, Sen SK. Optimization of fermentation conditions for cellulase production by Bacillus subtilis CY5 and Bacillus circulans TP3 isolated from fish gut. Acta Ichthyologica ET Piscatoria. 2007; 37(1): 47-53.
- Song FL, Forsberg CW, Gibbins LN. Cellulolytic activity of Clostridium acetobutylium. Appl Env Microbiol. 1985; 50: 220-8.
- Souichiro K, Shin H, Zong JC, Masaharu I, Akira Y, Yasuo I. Clostridium straminisolvens sp. nov., a moderately thermophilic, aerotolerant and cellulolytic bacterium isolated from a cellulose-degrading bacterial community. Int J Sys Evol Microbiol. 2004; 54: 2043-7.
- Spano L, Medeiros J, Mandels M. In: Enzymatic hydrolysis of cellulosic waste to glucose (Pollution Abatement Div., Food Svcs. Lab, US Army Natick Labs). Natick, MA; 1975.
- Spiridonov NA, Wilson DB. Regulation of biosynthesis of individual cellulases in Thermomonospora fusca. J Bacteriol. 1998; 180: 3529-3532.
- Suto M, Tomita F. Induction and catabolite repression mechanisms of cellulase in fungi. J Biosci Bioeng. 2001; 92(4): 305-311.
Publication Dates
-
Publication in this collection
08 Aug 2012 -
Date of issue
Aug 2012
History
-
Received
03 Nov 2010 -
Accepted
05 Mar 2012 -
Reviewed
21 Nov 2011