ABSTRACT
Rumen fungi inhabit the gastro-intestinal tract of ruminants and the most non-ruminant herbivores. Rumen fungi produce highly active plant cell wall degrading enzymes, therefore they have gained scientific interest. In this study, genes encoding xylanase (xynA-7) and cellulase (celA-5) were amplified from Neocallimastix sp. GMLF7 and Orpinomyces sp. GMLF5, respectively, and expressed in Escherichia coli. XynA-7 was found to be active only on xylan, however CelA-5 had activity both on carboxymethyl cellulose and lichenan. Lichenase activity of CelA-5 was found to be higher than carboxymethyl cellulase activity. The optimal conditions were at pH 6.0 and 40 °C for CelA-5 and at pH 6.5 and 50 °C for XynA-7. A coexpression vector was constructed to coproduce the XynA-7 and CelA-5 and then transformed into E. coli. The ability of the transformed E. coli strain to produce CMCase, xylanase and lichenase was evaluated. The transformed E. coli strain acquired the capacity to degrade CMC, xylan and lichenan.
Key words:
Rumen fungi; xylanase; cellulase; coexpression; E. coli
INTRODUCTION
The major components of a plant cell wall are cellulose and xylan 11 - Lee JH, Lim M, Kim M, Heo S, Seo J, Kim Y, et al. Constitutive coexpression of Bacillus endoxylanase and Trichoderma endoglucanase genes in Saccharomyces cerevisiae.J Microbiol Biotechn. 2007; 17(12): 2076. . Cellulose is composed of insoluble linear chains of β-D-1,4-linked glucose units and composed of crystalline and amorphous regions 22 - Howard RL, Abotsi E, Van Rensburg EJ, Howard S. Lignocellulose biotechnology: issues of bioconversion and enzyme production. Afr J Biotechnol. 2004; 2(12): 602-619. . Xylan is a main component of the hemicellulose, and it is a complex polymer consisting of a β-D-1,4-linked xylopyranoside backbone which is partially substituted with acetyl, arabinosyl, and glucuronosyl side chains 33 - Katahira S, Fujita Y, Mizuike A, Fukuda H, Kondo A. Construction of a xylan-fermenting yeast strain through codisplay of xylanolytic enzymes on the surface of xylose-utilizing Saccharomyces cerevisiae cells. Appl Environ Microbiol. 2004; 70(9): 5407-5414.. Many microorganisms are capable of degrading and utilizing cellulose and xylan as carbon and energy sources 44 - Sánchez C. Lignocellulosic residues: biodegradation and bioconversion by fungi.Biotechnol Adv. 2009; 27(2): 185-194.. Cellulase (endo-1,4-β-glucanase, EC 3.2.1.4) randomly attacks the internal O-glycosidic bonds in cellulose, resulting in glucan chains of different lengths 55 - Kuhad RC, Gupta R, Singh A. Microbial cellulases and their industrial applications.Enzyme Research. 2011; Article ID 280696, 1-10.. Xylanase (endo-1,4-β-xylanase, E.C.3.2.1.8) acts on homopolymeric backbone of 1,4-linked β-D-xylopyranose producing xylooligomers66 -J uturu V, Wu JC. Microbial xylanases: engineering, production and industrial applications.Biotechnol Adv. 2012; 30(6): 1219-1227. . Cellulases and xylanases are receiving considerable attention because of their potential applications in several industrial processes.
The rumen contains a complex microbial ecosystem, which extracts nutrients from lignocellulosic agricultural by-products 77 - Kamra DN. Rumen microbial ecosystem.Curr Sci India. 2005; 89(1): 124-135.. Rumen fungi are obligate anaerobic microorganisms and constitute a significant portion of the rumen microbial biomass 88 -Sirohi SK, Singh N, Dagar SS, Puniya AK. Molecular tools for deciphering the microbial community structure and diversity in rumen ecosystem.Appl Microbiol Biot. 2012; 95(5): 1135-1154. . Rumen fungi rapidly colonize plant cell walls 99 -Borneman WS, Ljungdahl LG, Hartley RD, Akin DE. Isolation and characterization of p-coumaryl esterase from the anaerobic fungus Neocallimastix strain MC-2. Appl Environ Microbiol. 1991; 57: 2377-2344. . They degrade cell wall carbohydrates such as cellulose and hemicellulose from the plant parts where they attached 1010 -Grenet E, Barry P. Colonization of thick-walled plant tissues by anaerobic fungi. Anim Fd Sci Technol. 1988; 19: 25-31.. This degradation is performed by various highly active cellulases, xylanases, glycosidases and xylosidases of rumen fungi 1111 -Comlekcioglu U, Ozkose E, Tutus A, Akyol I, Ekinci MS. Cloning and characterization of cellulase and xylanase coding genes from anaerobic fungus Neocallimastix sp. GMLF1.Int J Agric Biol. 2010; 12(5): 691-696.. The cellulolytic and xylanolytic capacity of rumen fungi was also studied in detail by characterizing the enzyme encoding genes. Several cellulase and xylanase encoding genes have been cloned from Neocallimastix sp., Orpinomyces sp. and Piromyces sp. 1212 -Krause DO, Denman SE, Mackie RI, Morrison M, Rae AL, Attwood GT, et al. Opportunities to improve fiber degradation in the rumen: microbiology, ecology, and genomics.FEMS Microbiol Rev. 2003; 27(5): 663-693.. Recently, 357 glycoside hydrolases, 24 polysaccharide lyases and 92 carbohydrate esterases were reported in the genome of Orpinomyces sp. C1A 1313 -Youssef NH, Couger MB, Struchtemeyer CG, Liggenstoffer AS, Prade RA, Najar FZ, et al. The genome of the anaerobic fungus Orpinomyces sp. strain C1A reveals the unique evolutionary history of a remarkable plant biomass degrader. Appl Environ Microbiol. 2013; 79(15): 4620-4634..
Neocallimastix spp. and Orpinomyces spp. are well studied microorganisms and they have been reported as excellent cellulase and xylanase producers. In this study, we describe the cloning, coexpression and secretion of the xylanase encoding gene xynA-7 from Neocallimastix sp. GMLF7, and cellulase encoding bifunctional gene celA-5 from Orpinomyces sp. GMLF5 in Escherichia coli. We also conducted the partial characterization and examined the activity of the recombinant enzymes.
MATERIALS AND METHODS
Microorganisms and Growth Conditions
The rumen fungi were grown in anaerobic medium at 39 oC. Anaerobic medium was prepared according to Orpin 1414 - Orpin CG. Studies on the rumen flagellate Sphaeromonas communis. J Gen Microbiol. 1976; 94: 270-280. and dispensed into Hungate tubes under strictly anaerobic conditions. Wheat straw was used as energy source for the maintenance media, and glucose was used as energy source to obtain relatively higher amounts of cells for DNA extraction 1515 -Comlekcioglu U, Ozkose E, Akyol I, Yazdic FC, Ekinci MS. Expression of β-(1, 3-1, 4)-glucanase gene of Orpinomyces sp. GMLF18 in Escherichia coli EC1000 and Lactococcus lactis subsp. cremoris MG1363. Turk J Biol. 2011; 35: 405-414.. Escherichia coli strains were grown in Luria-Bertani (LB) broth in a shaking incubator (150 rpm) at 37 oC. The list of fungal and E. coli strains is given in Table 1.
Genomic DNA Extraction and Polymerase Chain Reaction
Rumen fungi were grown for 2 days and the fungal biomass was harvested by centrifugation at 1250 g for 10 min. Biomass was frozen using liquid nitrogen and immediately broken down using a Mixer Mill (Retsch MM301). Genomic DNA extraction was performed according to Chen et al. 1919 -Chen H, Hopper SL, Li XL, Ljungdahl LG, Cerniglia CE. Isolation of extremely AT-rich genomic DNA and analysis of genes encoding carbohydrate-degrading enzymes from Orpinomyces sp. strain PC-2.Curr Microbiol. 2006; 53(5): 396-400. . The xylanase and cellulase encoding genes were amplified from Neocallimastix sp. GMLF7 and Orpinomyces sp. GMLF5, respectively, by using xylanase primers (F: 5’ACTGTTGCTAAGGCCCAATG-3’ and R: 5’-ACCCCATTTACCATCGTCATC-3') and the cellulase primers (F: 5’-ATTATATTCATATTCACTGGTTGAA-3' and R: 5’-TTAGAATGGTGGGTTAGCATTT-3').
Plasmids and Cloning Strategy
The pCT (Favorgen, Taiwan) and pGEMT-Easy (Promega, USA) vectors were used for construction of xylanase and cellulase expression vectors, respectively. The list of constructed vectors was given in Table 1. The constructed vectors were transformed into E. coli EC1000 and by using CaCl2 method 2020 -Mandel M, Higa A. Calcium-dependent bacteriophage DNA infection. J Mol Biol. 1970; 53: 159-162.. Plasmid isolations were carried out with Plasmid Isolation Kit (Vivantis, Malaysia). Cellulase gene digested from pGEMC with EcoRI and ligated into pCTX7. The resultant plasmid (pCTXC) was transformed into E. coli EC1000 and E. coli MC1022. Xylanase and cellulase positive colonies were screened according to Teather and Wood 2121 -Teather RM, Wood PJ. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol. 1982; 43: 777-780..
Enzyme Assay
For investigation of the cell-associated and extracellular enzyme activities, transformant E. coli strain was transferred to LB broth containing ampicillin (50µg/ml) and incubated at 37 oC for 12h in a shaking incubator (150 rpm). The culture was centrifuged at 6000 rpm for 10 min at 4 oC. The supernatant was collected and used as extracellular enzyme. The cell pellet washed twice with 50 mM sodium-phosphate buffer (pH 6.0) and then broken down by using ball-mill dismembranator (Retsch, Germany) and resuspended in the same buffer. Cellular debris was subsequently removed by centrifugation and the clarified extract used as cell-associated activity. Extracellular and cell-associated enzyme fractions were stored at -20 oC until required. Xylanase, cellulase, avicelase and lichenase activities were determined by measuring the amount of reducing sugar released by enzyme fractions incubated with 0.5% birchwood xylan (Sigma), 0.5% carboxymethyl cellulose (Sigma), 0.2% Avicel (Merck), and 0.1% lichenan (Megazyme), respectively, in 50 mM sodium phosphate buffer (pH 6.0). Reducing sugars released were measured by the dinitrosalicyclic acid reagent 2222 -Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem. 1959; 31: 426-428.. One unite of enzyme activity was defined as the amount of enzyme that releases 1 μmol of reducing sugar per minute in the reaction mixture under the assay conditions. Optimum pH and temperature, pH and temperature stabilities were determined according to Comlekcioglu et al. 2323 -Comlekcioglu U, Tutus A, Cicekler M, Gunes M, Aygan A. Application of recombinant xylanase from Orpinomyces sp. in elemental chlorine-free bleaching of kraft pulps.Rom Biotech Lett. 2014; 19(1): 8941-8949. .
RESULTS
Cloning of xynA-7 and celA-5
The endo-β-1,4-xylanase (xynA-7) and endo-β-1,4-glucanase (celA-5) genes were amplified from Neocallimastix sp. GMLF7 and Orpinomyces sp. GMLF5, respectively, using the genomic DNA as the template. The xynA-7 and celA-5 fragments were about 800 and 1500 bp and cloned into pCT and pGEMT-Easy vectors, respectively. The resultant plasmids were named as pCTX7 and pGEMC, and transferred to E. coli EC1000. Recombinant E. coli pCTX7 and E. coli pGEMC strains were produced XynA-7 and CelA5 enzymes, respectively.
Characterization of XynA-7 and CelA-5 activities
XynA-7 and CelA-5 enzymes were produced from recombinant E. coli strains. The substrate specificities of the recombinant enzymes for xylan, CM-cellulose, lichenan and avicel were determined. The activities were determined for both cell associated and supernatant fraction. The major enzymatic activities of XynA-7 and CelA-5 were found to be cell associated in E. coli. More than 90% of the XynA-7 and CelA-5 activities were recovered from cellular biomass. Therefore, characterization of the enzymes was carried out using the cell associated enzyme in further studies. XynA-7 was found to be active only on xylan. CelA-5 had activity both on CMC and lichenan, however activity of CelA-5 against lichenan was much higher than CMCase activity. Avicelase activity was also tested for CelA-5, however no avicelase activity was detected in CelA-5.
The activities of the two enzymes at various temperatures and pH values are shown in Figure 1. XynA-7 and CelA-5 displayed their highest activities at 50 and 40°C, respectively. XynA-7 retained 70% of its activity after preincubation at 50°C for 3 h in the absence of substrate. The maximum thermal stability was obtained at 40 oC for CelA-5 and 59% of activity was still remained after 3 h incubation, although the CelA-5 activity diminished at 50°C after 1 h. The maximum activities were at pH 6.0 for CelA-5 and at pH 6.5 for XynA-7. Enzyme activities greater than 60% of the highest activity were attained at pH 5.0 to 7.0 for CelA-5 and at pH 4.0 to 8.0 for XynA-7. After preincubation at different pH values at optimum temperature for 30 min, both enzymes lost more than 60% of the maximal activity above pH 7.0.

Effects of temperature (A) and pH (B) on the activities of XynA-7 (solid line) and CelA-5 (dashed line). (C) Thermal stability at 40 oC (() and 50 oC ((). (D) pH stability after incubation for 30 min at optimum temperature.
Construction of co-expression vector
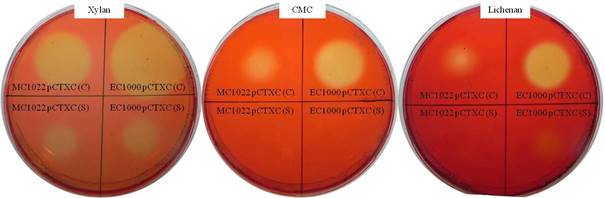
To coexpress the xynA-7 gene of Neocallimastix sp. GMLF7 (800 bp fragment) and celA-5 gene of Orpinomyces sp. GMLF5 (1500 bp fragment), celA-5 were cloned into the vector pCTX7, generating pCTXC. celA-5 fragment was obtained by digestion from the pGEMC vector using EcoRI, and ligated to a similarly digested pCTX7 (Fig. 2). The resultant plasmid (pCTXC) was then transformed into E. coli EC1000 and E. coli MC1022 as hosts. The presence of the pCTXC in the E .coli transformants was investigated by screening the CMCase activity using Congo red plate assay. A CMCase positive colony was selected and the recombinant plasmid pCTXC was isolated. pCTXC was verified by EcoRI digestion, which produced the celA-5 fragment (1500 bp) (Figure 3). A Congo red plate assay was also used to confirm the xylanase and lichenase activities of the selected CMCase positive transformant. The cell lysates of the E. coli pCTXC strains produced yellow halo zones on the substrate containing plate, indicating that xynA-7 and bifunctional celA-5 were functionally coexpressed by pCTXC containing E. coli transformants (Fig. 4). Enzyme production of E. coli EC1000/pCTXC was found to be higher than E. coli MC1022/pCTXC, hence pCTXC harboring E. coli EC1000 was used in further studies.

Schematic diagram of xylanase gene containing pCTX7 and bifunctional gene containing pGEMC. pCTX7 and pGEMC were digested with EcoRI, celA-5-EcoRI fragment was isolated from the gel and ligated to pCTX7-EcoRI fragment. Lane 1, 100 bp ladder; lane 2, EcoRI digestion of pCTX7; lane 3, EcoRI digestion of pGEMC.

Confirmation of celA-5 gene in pCTXC by EcoRI digestion. Lane 1, 100 bp ladder; lane 2, EcoRI digestion of pGEMC; lane 3, EcoRI digestion of pCTX7; lane 4, EcoRI digestion of pCTXC.

Plate tests for xylanase, CMCase and lichenase activities of pCTXC harboring E. coli EC1000 and MC1022. Extracellular (S) and cell associated (C) enzyme fractions were pipetted onto xylan, CMC and lichenan containing agarose plates, allowed to incubate and stained with Congo red.
Enzyme production of E. coli pCTXC strains
When cultured in the LB medium at 37°C, E. coli pCTXC reached stationary growth after 12 h, which was similar to that observed for E. coli EC1000 (data not shown). The specific activities of xynA-7 and celA-5 were determined in the LB medium. The host strain E. coli EC1000 did not show any activity. The highest activities observed for xylanase, CMCase and lichenase of E. coli pCTXC were 30.88, 1.83 and 11.48 U ml-1, respectively. Xylanase and lichenase activities of pCTXC were similar with XynA-7 and CelA-5 expressed from separate promoters. As shown in Table 2, the xylanase and lichenase activity in the extracellular fraction of the E. coli pCTXC culture were much higher than that of the cell associated fraction. However, CMCase activity of cell associated fraction of E. coli pCTXC was lower from the extracellular fraction.
DISCUSSION
Enzyme supplementation to improve the feed utilization by ruminants has an increasing attention. Supplementing dairy cow and feedlot cattle with fibrolytic enzymes has potential for improving the efficiency of feed utilization and animal performance 2424 -Beauchemin KA, Colombatto D, Morgavi DP, Yang WZ. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J Anim Sci. 2003; 81: E37-E47.. Fibrolytic enzyme mixture is reported to enhance the milk yield of cows in early lactation without changing feed intake 2525 -Rode LM, Yang WZ, Beauchemin KA. Fibrolytic enzyme supplements for dairy cows in early lactation.J Dairy Sci. 1999; 82(10): 2121-2126. . Complete degradation of cellulose and hemicellulose in feeds requires a number of enzymes 2626 -McAllister TA, Hristov AN, Beauchemin KA, Rode LM, Cheng KJ. Enzymes in ruminant diets. In: Bedford MR, Partridge GG. editors. Enzymes in Farm Nutrition. London: Oxon: Cab International; 2001. p. 273-298.. Recently, recombinant DNA technology has enabled the bacteria to coexpress the specific genes. Genes encoding superior genes can be combined in a vector and transferred into well characterized microbial hosts.
Rumen fungi produce a wide range of plant cell wall degrading enzymes that enable them to utilize fibrous plant biomass efficiently. Studies on the functional transfer of rumen fungal genes to tractable bioprocessing hosts are rapidly emerging2727 -Haitjema CH, Solomon KV, Henske JK, Theodorou MK, O'Malley MA. Anaerobic gut fungi: Advances in isolation, culture, and cellulolytic enzyme discovery for biofuel production.Biotechnol Bioeng. 2014; 111(8): 1471-1482. . In this study, xylanase encoding xynA-7 and cellulase and lichenase encoding bifunctional celA-5 were cloned into E. coli and we performed partial characterization of XynA-7 and CelA-5. Both enzymes showed maximum activity in the range of 40-50°C and pH 6 to 6.5. Optimum pH and temperature values are found to be similar with other rumen fungal enzymes 2828 - Li XL, Chen H, Ljungdahl LG. Two cellulases, CelA and CelC, from the polycentric anaerobic fungus Orpinomyces strain PC-2 contain N-terminal docking domains for a cellulase-hemicellulase complex.Appl Environ Microbiol. 1997; 63(12): 4721-4728. ,2929 - Huang YH, Huang CT, Hseu RS. Effects of dockerin domains on Neocallimastix frontalis xylanases.FEMS Microbiol Lett. 2005; 243(2): 455-460.,1111 -Comlekcioglu U, Ozkose E, Tutus A, Akyol I, Ekinci MS. Cloning and characterization of cellulase and xylanase coding genes from anaerobic fungus Neocallimastix sp. GMLF1.Int J Agric Biol. 2010; 12(5): 691-696.. The similar optimum activities of XynA-7 and CelA-5 are a prominent property for the simultaneous production of both enzymes in the same host.
E. coli has a number of benefits for recombinant protein expression, such as low production cost, rapid growth and high yields to produce desired products 3030 -Liu M, Yu H. Co-production of a whole cellulase system in Escherichia coli.Biochem Eng J. 2012; 69: 204-210.. While numerous cellulose and xylan degrading enzymes have been cloned in E. coli, to the best of our knowledge, there are few reports representing the co-expression of cellulase and xylanase in E. coli3131 - Adlakha N, Rajagopal R, Kumar S, Reddy VS, Yazdani SS. Synthesis and characterization of chimeric proteins based on cellulase and xylanase from an insect gut bacterium.Appl Environ Microbiol. 2011; 77(14): 4859-4866.. In this study, two E. coli strains, E. coli EC1000 and MC1022, were tested for the coexpression of xynA-7 and celA-5, and E. coli EC1000 harboring the pCTXC plasmid had more enzyme activity than that of E. coli MC1022 containing the pCTXC. These results indicate that difference in E. coli strains are a important factor for high production of coexpressed enzymes. Simultaneous expression of genes can be conducted using either single or multiple plasmids in E. coli, and in the case of a single plasmid, genes can be controlled by single or seperate promoter 3030 -Liu M, Yu H. Co-production of a whole cellulase system in Escherichia coli.Biochem Eng J. 2012; 69: 204-210..
CONCLUSION
We have established a coexpression system for the production of XynA-7 and bifunctional CelA-5 in E. coli. Coexpressed xylanase and bifunctional cellulase enzymes exhibited properties similar to those of the individually expressed enzymes. In addition to XynA-7 and CelA-5, other polysaccharide degrading enzymes used in animal nutrition, waste treatment, paper manufacturing and fuel production could also be combined in co-expression systems. The use of a single host cell for the production of enzymes may allow for cost-effective enzyme formulations. Additionally, coexpression vectors could also be designed with different strategies, such as using other genes, various promoters and selection markers in order to obtain high production and specifity.
ACKNOWLEDGEMENT
This work was supported by Kahramanmaraş Sutcu Imam University (Grants: 2012/2-13YLS and 2011/4-14YLS). We gratefully thank Dr. Ismail Akyol, Dr. Emin Ozkose and Dr. M. Sait Ekinci for their valuable contributions.
REFERENCES
-
1- Lee JH, Lim M, Kim M, Heo S, Seo J, Kim Y, et al. Constitutive coexpression of Bacillus endoxylanase and Trichoderma endoglucanase genes in Saccharomyces cerevisiaeJ Microbiol Biotechn 2007; 17(12): 2076.
-
2- Howard RL, Abotsi E, Van Rensburg EJ, Howard S. Lignocellulose biotechnology: issues of bioconversion and enzyme production. Afr J Biotechnol 2004; 2(12): 602-619.
-
3- Katahira S, Fujita Y, Mizuike A, Fukuda H, Kondo A. Construction of a xylan-fermenting yeast strain through codisplay of xylanolytic enzymes on the surface of xylose-utilizing Saccharomyces cerevisiae cells. Appl Environ Microbiol 2004; 70(9): 5407-5414.
-
4- Sánchez C. Lignocellulosic residues: biodegradation and bioconversion by fungi.Biotechnol Adv 2009; 27(2): 185-194.
-
5- Kuhad RC, Gupta R, Singh A. Microbial cellulases and their industrial applications.Enzyme Research 2011; Article ID 280696, 1-10.
-
6-J uturu V, Wu JC. Microbial xylanases: engineering, production and industrial applications.Biotechnol Adv 2012; 30(6): 1219-1227.
-
7- Kamra DN. Rumen microbial ecosystem.Curr Sci India 2005; 89(1): 124-135.
-
8-Sirohi SK, Singh N, Dagar SS, Puniya AK. Molecular tools for deciphering the microbial community structure and diversity in rumen ecosystem.Appl Microbiol Biot 2012; 95(5): 1135-1154.
-
9-Borneman WS, Ljungdahl LG, Hartley RD, Akin DE. Isolation and characterization of p-coumaryl esterase from the anaerobic fungus Neocallimastix strain MC-2. Appl Environ Microbiol 1991; 57: 2377-2344.
-
10-Grenet E, Barry P. Colonization of thick-walled plant tissues by anaerobic fungi. Anim Fd Sci Technol 1988; 19: 25-31.
-
11-Comlekcioglu U, Ozkose E, Tutus A, Akyol I, Ekinci MS. Cloning and characterization of cellulase and xylanase coding genes from anaerobic fungus Neocallimastix sp. GMLF1.Int J Agric Biol 2010; 12(5): 691-696.
-
12-Krause DO, Denman SE, Mackie RI, Morrison M, Rae AL, Attwood GT, et al. Opportunities to improve fiber degradation in the rumen: microbiology, ecology, and genomics.FEMS Microbiol Rev 2003; 27(5): 663-693.
-
13-Youssef NH, Couger MB, Struchtemeyer CG, Liggenstoffer AS, Prade RA, Najar FZ, et al. The genome of the anaerobic fungus Orpinomyces sp. strain C1A reveals the unique evolutionary history of a remarkable plant biomass degrader. Appl Environ Microbiol 2013; 79(15): 4620-4634.
-
14- Orpin CG. Studies on the rumen flagellate Sphaeromonas communis J Gen Microbiol 1976; 94: 270-280.
-
15-Comlekcioglu U, Ozkose E, Akyol I, Yazdic FC, Ekinci MS. Expression of β-(1, 3-1, 4)-glucanase gene of Orpinomyces sp. GMLF18 in Escherichia coli EC1000 and Lactococcus lactis subsp. cremoris MG1363. Turk J Biol 2011; 35: 405-414.
-
16-Çömlekçioğlu U, Akyol İ, Kar B, Özköse E, Ekinci MS. Anaerobik rumen funguslarının izolasyonu, tanımlanması ve kültür koleksiyonunun oluşturulması.Hayvansal Üretim 2008; 49(2): 29-35.
-
17- Leenhouts KJ, Buist G, Bolhuis A, ten Berge A, Kiel J, Mierau I, et al. A general system for generating unlabeled gene replacements in bacterial chromosomes. Mol Gen Genet 1996; 253: 217-224.
-
18-Casadaban MJ, Cohen SN. Analysis of gene control signals by DNA fusion and cloning in Escherichia coliJ Mol Biol 1980; 138(2): 179-207.
-
19-Chen H, Hopper SL, Li XL, Ljungdahl LG, Cerniglia CE. Isolation of extremely AT-rich genomic DNA and analysis of genes encoding carbohydrate-degrading enzymes from Orpinomyces sp. strain PC-2.Curr Microbiol 2006; 53(5): 396-400.
-
20-Mandel M, Higa A. Calcium-dependent bacteriophage DNA infection. J Mol Biol 1970; 53: 159-162.
-
21-Teather RM, Wood PJ. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol 1982; 43: 777-780.
-
22-Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem 1959; 31: 426-428.
-
23-Comlekcioglu U, Tutus A, Cicekler M, Gunes M, Aygan A. Application of recombinant xylanase from Orpinomyces sp. in elemental chlorine-free bleaching of kraft pulps.Rom Biotech Lett 2014; 19(1): 8941-8949.
-
24-Beauchemin KA, Colombatto D, Morgavi DP, Yang WZ. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J Anim Sci 2003; 81: E37-E47.
-
25-Rode LM, Yang WZ, Beauchemin KA. Fibrolytic enzyme supplements for dairy cows in early lactation.J Dairy Sci 1999; 82(10): 2121-2126.
-
26-McAllister TA, Hristov AN, Beauchemin KA, Rode LM, Cheng KJ. Enzymes in ruminant diets. In: Bedford MR, Partridge GG. editors. Enzymes in Farm Nutrition. London: Oxon: Cab International; 2001. p. 273-298.
-
27-Haitjema CH, Solomon KV, Henske JK, Theodorou MK, O'Malley MA. Anaerobic gut fungi: Advances in isolation, culture, and cellulolytic enzyme discovery for biofuel production.Biotechnol Bioeng 2014; 111(8): 1471-1482.
-
28- Li XL, Chen H, Ljungdahl LG. Two cellulases, CelA and CelC, from the polycentric anaerobic fungus Orpinomyces strain PC-2 contain N-terminal docking domains for a cellulase-hemicellulase complex.Appl Environ Microbiol 1997; 63(12): 4721-4728.
-
29- Huang YH, Huang CT, Hseu RS. Effects of dockerin domains on Neocallimastix frontalis xylanases.FEMS Microbiol Lett 2005; 243(2): 455-460.
-
30-Liu M, Yu H. Co-production of a whole cellulase system in Escherichia coliBiochem Eng J 2012; 69: 204-210.
-
31- Adlakha N, Rajagopal R, Kumar S, Reddy VS, Yazdani SS. Synthesis and characterization of chimeric proteins based on cellulase and xylanase from an insect gut bacterium.Appl Environ Microbiol 2011; 77(14): 4859-4866.
Publication Dates
-
Publication in this collection
2017
History
-
Received
03 Feb 2016 -
Accepted
14 July 2016