Resumos
Em humanos saudáveis, nove aminoácidos são considerados essenciais, uma vez que não podem ser sintetizados endogenamente e, portanto, devem ser ingeridos por meio da dieta. Dentre os aminoácidos essenciais, se incluem os três aminoácidos de cadeia ramificada, ou seja, leucina, valina e isoleucina. Esses aminoácidos participam da regulação do balanço protéico corporal além de serem fonte de nitrogênio para a síntese de alanina e glutamina. No tocante à regulação da síntese protéica muscular, verifica-se que a leucina age estimulando a fase de iniciação da tradução do RNA-mensageiro em proteína, por mecanismos tanto dependentes quanto independentes de insulina. No que concerne ao exercício físico, supõe-se que esses aminoácidos estejam envolvidos na fadiga central, no balanço protéico muscular, na secreção de insulina, na modulação da imunocompetência, no aumento da performance de indivíduos que se exercitam em ambientes quentes e na diminuição do grau de lesão muscular. Nesse contexto, essa revisão aborda os aspectos atuais do metabolismo e da suplementação de aminoácidos de cadeia ramificada no exercício físico.
Aminoácidos de cadeia ramificada; Exercício físico; Fadiga central; Imunocompetência; Lesão muscular; Síntese protéica
In healthy humans, nine amino acids are considered to be essential once they cannot be endogenously synthesised and must therefore be ingested in the diet. Amongst the essential amino acids are the three branched chain amino acids, namely, leucine, valine and isoleucine. These amino acids participate in the regulation of protein balance in addition to being nitrogen sources for the synthesis of alanine and glutamine. As to the regulation of muscle protein synthesis, leucine acts in the stimulation of initiation of mRNA translation into protein, both through mechanisms that are dependent and independent of insulin. In the physiology of physical exercise, these branched amino acids play a role in central fatigue hypothesis, in muscle protein balance, in the secretion of insulin, in the modulation of the immune response, in performance enhancement of individuals who work out in hot environments, and in avoiding muscle lesion. This review approaches all aspects of the metabolism of and supplementation with branched chain amino acids in physical exercise.
Branched chain amino acids; Physical exercise; Central fatigue; Immune response; Muscle lesion; Protein synthesis
REVISÃO
Aspectos atuais sobre aminoácidos de cadeia ramificada e exercício físico
Current aspects of branched chain amino acid and exercise
Marcelo Macedo Rogero; Julio Tirapegui* * Correspondência: J. Tirapegui Departamento de Alimentos e Nutrição Experimental Av. Prof. Lineu Prestes, 580, Bloco 14 Faculdade de Ciências Farmacêuticas Universidade de São Paulo 05508-900 - São Paulo - SP, Brasil E-mail: tirapegu@usp.br
Departamento de Alimentos e Nutrição Experimental, Faculdade de Ciências Farmacêuticas, Universidade de São Paulo
RESUMO
Em humanos saudáveis, nove aminoácidos são considerados essenciais, uma vez que não podem ser sintetizados endogenamente e, portanto, devem ser ingeridos por meio da dieta. Dentre os aminoácidos essenciais, se incluem os três aminoácidos de cadeia ramificada, ou seja, leucina, valina e isoleucina. Esses aminoácidos participam da regulação do balanço protéico corporal além de serem fonte de nitrogênio para a síntese de alanina e glutamina. No tocante à regulação da síntese protéica muscular, verifica-se que a leucina age estimulando a fase de iniciação da tradução do RNA-mensageiro em proteína, por mecanismos tanto dependentes quanto independentes de insulina. No que concerne ao exercício físico, supõe-se que esses aminoácidos estejam envolvidos na fadiga central, no balanço protéico muscular, na secreção de insulina, na modulação da imunocompetência, no aumento da performance de indivíduos que se exercitam em ambientes quentes e na diminuição do grau de lesão muscular. Nesse contexto, essa revisão aborda os aspectos atuais do metabolismo e da suplementação de aminoácidos de cadeia ramificada no exercício físico.
Unitermos: Aminoácidos de cadeia ramificada. Exercício físico. Fadiga central. Imunocompetência. Lesão muscular. Síntese protéica
ABSTRACT
In healthy humans, nine amino acids are considered to be essential once they cannot be endogenously synthesised and must therefore be ingested in the diet. Amongst the essential amino acids are the three branched chain amino acids, namely, leucine, valine and isoleucine. These amino acids participate in the regulation of protein balance in addition to being nitrogen sources for the synthesis of alanine and glutamine. As to the regulation of muscle protein synthesis, leucine acts in the stimulation of initiation of mRNA translation into protein, both through mechanisms that are dependent and independent of insulin. In the physiology of physical exercise, these branched amino acids play a role in central fatigue hypothesis, in muscle protein balance, in the secretion of insulin, in the modulation of the immune response, in performance enhancement of individuals who work out in hot environments, and in avoiding muscle lesion. This review approaches all aspects of the metabolism of and supplementation with branched chain amino acids in physical exercise.
Uniterms: Branched chain amino acids. Physical exercise. Central fatigue. Immune response. Muscle lesion. Protein synthesis
INTRODUÇÃO
Aminoácidos são as unidades básicas da composição de uma proteína. Em humanos saudáveis, nove aminoácidos são considerados essenciais, uma vez que não podem ser sintetizados endogenamente e, portanto, devem ser ingeridos por meio da dieta. Dentre os aminoácidos essenciais, se incluem os três aminoácidos de cadeia ramificada (ACR), ou seja, leucina, valina e isoleucina, que apresentam, respectivamente, concentração plasmática média de 120, 220 e 63 µmol/L; concentração intramuscular na forma livre média de 133, 253 e 68 µmol/L de água intracelular; e concentração na proteína muscular humana de 59,5, 43,5 e 41,9 mmol/100 g de proteína. A concentração de ACR também difere em relação ao tipo de fibra muscular, sendo 20-30% maior em fibras de contração lenta em comparação àquelas de contração rápida. Os ACR correspondem a cerca de 35% dos aminoácidos essenciais em proteínas musculares e, uma vez que a massa muscular de humanos é de cerca de 40-45% da massa corporal total, verifica-se que grande quantidade de ACR está presente em proteínas musculares (Marchini et al., 1998; Wagenmakers, 1998).
Em indivíduos adultos, ACR são relevantes para a manutenção da proteína corporal além de serem fonte de nitrogênio para a síntese de alanina e glutamina. Existem evidências demonstrando o papel fundamental dos ACR - especialmente a leucina - na regulação de processos anabólicos envolvendo tanto a síntese quanto a degradação protéica muscular. Além disso, ACR apresentam potenciais efeitos terapêuticos, uma vez que esses aminoácidos podem atenuar a perda de massa magra durante a redução de massa corporal; favorecer o processo de cicatrização; melhorar o balanço protéico muscular em indivíduos idosos; e propiciar efeitos benéficos no tratamento de patologias hepáticas e renais (Shimomura et al., 2006a; Tom, Nair, 2006).
No que concerne a nutrição esportiva, os ACR são extensivamente utilizados por atletas baseado na premissa de que esses aminoácidos podem promover anabolismo protéico muscular, atuar em relação à fadiga central, favorecer a secreção de insulina, melhorar a imunocompetência, diminuir o grau de lesão muscular induzido pelo exercício físico e aumentar a performance de indivíduos que se exercitam em ambientes quentes.
METABOLISMO DOS AMINOÁCIDOS DE CADEIA RAMIFICADA
No tocante ao metabolismo dos ACR, inicialmente cabe ressaltar as vias bioquímicas envolvidas no catabolismo desses aminoácidos. Diferentemente de outros aminoácidos, que são oxidados primariamente no tecido hepático, o sistema enzimático mais ativo para a oxidação dos ACR está localizado no músculo esquelético. Apesar do fígado não poder diretamente catabolizar os ACR, o mesmo apresenta um sistema muito ativo para a degradação dos cetoácidos de cadeia ramificada oriundos dos correspondentes ACR. Essa distribuição tecidual específica do catabolismo dos ACR decorre da distribuição única das duas primeiras enzimas envolvidas no catabolismo dos ACR, ou seja, aminotransferase de aminoácidos de cadeia ramificada (ATACR) - que catalisa a transaminação dos ACR, em reação reversível - e o complexo enzimático desidrogenase de cetoácidos de cadeia ramificada (DCCR) - que catalisa a descarboxilação oxidativa dos cetoácidos de cadeia ramificada, em reação irreversível (Shimomura et al., 2006a; Shimomura et al., 2006b; Shimomura, Harris, 2006).
A primeira reação envolvida no catabolismo dos ACR é a transaminação pelas isoenzimas ATACR - que são enzimas dependentes de piridoxal-fosfato (vitamina B6) -, e que aceitam os três ACR como substratos. No que concerne à atividade tecidual da enzima ATACR (atividade por grama de tecido úmido), verifica-se elevada atividade no coração e rim, atividade intermediária no músculo esquelético e baixa atividade no fígado. Em células de mamíferos, duas ATACR estão presentes, sendo uma mitocondrial e outra citosólica. A partir da reação catalisada pela ATACR, os ACR são convertidos nos seus respectivos cetoácidos, ou seja, a leucina é convertida em α-cetoisocaproato (KIC); a isoleucina em α-ceto-β-metilvalerato (KMV); e a valina em α-cetoisovalerato (KIV). Concomitantemente, verifica-se que na reação catalisada pela ATACR há a conversão de α-cetoglutarato - aceptor de nitrogênio oriundo dos ACR - em glutamato. A partir do glutamato pode ocorrer a síntese de outros aminoácidos, como alanina e glutamina. Desse modo, a transaminação dos ACR fornece mecanismos para transferir o nitrogênio dos ACR de acordo com a necessidade do tecido por glutamato e outros aminoácidos não-essenciais. Além disso, cabe ressaltar que as isoenzimas ATACR em mamíferos são muitos específicas para ACR e glutamato, sendo a preferência de substratos a seguinte: isoleucina > valina >> glutamato (Cynober, Harris, 2006; Harris et al., 2005; Hutson, Sweatt, Lanoue, 2005; Harris, Joshi, Jeoung, 2004).
Posteriormente à reação catalisada pela enzima ATACR e à conseqüente formação dos cetoácidos de cadeia ramificada, esses podem sofrer descarboxilação oxidativa mediada pelo complexo enzimático DCCR - presente na superfície interna da membrana interna mitocondrial. Por meio da reação catalisada pelo complexo DCCR, os cetoácidos de cadeia ramificada KIC, KMV e KIV são convertidos em isovaleril-CoA, 3-metilbutiril-CoA e isobutiril-CoA, respectivamente. A atividade da DCCR é maior no fígado, intermediária no rim e coração, e comparativamente baixa no músculo, tecido adiposo e cérebro (Harper, Miller, Block, 1984).
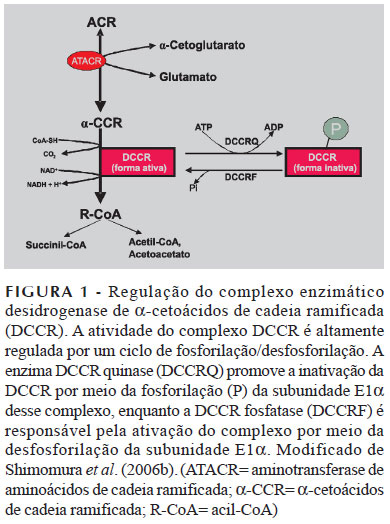
A DCCR é a principal enzima regulatória na via catabólica dos ACR, sendo considerada a etapa controladora do fluxo do catabolismo dos ACR. A atividade do complexo DCCR - diferentemente da atividade da ATACR - é altamente regulada por um ciclo de fosforilação/desfosforilação. A enzima DCCR quinase (DCCRQ) promove a inativação da DCCR por meio da fosforilação da subunidade E1α desse complexo, enquanto a DCCR fosfatase (DCCRF) é responsável pela ativação do complexo por meio da desfosforilação da subunidade E1α (Figura 1) (Harris, Joshi, Jeoung, 2004; Shimomura et al., 2004).
A ativação do complexo DCCR pode ser obtida em curto prazo pela inibição da atividade da enzima DCCRQ por meio do KIC - produto resultante da transaminação da leucina. Análogos estruturais do KIC, incluindo o octanoato, α-cloro-isocaproato e o ácido clofíbrico, também promovem a ativação do complexo DCCR por meio da inibição direta da DCCRQ. Os mecanismos de controle de longo prazo incluem: (i) diminuição da expressão gênica das subunidades da DCCR por meio da baixa ingestão de proteínas; (ii) aumento da expressão da DCCRQ induzida pela dieta com baixo teor de proteínas e por hormônios da tireóide; e (iii) diminuição da expressão da DCCRQ decorrente da alta ingestão de proteínas, jejum, glicocorticóides e clofibrato. A partir desses fatos, verifica-se que a atividade da DCCR é significativamente diminuída em animais alimentados com dietas hipoprotéicas ou tratados com hormônios da tireóide, porém a atividade da DCCR aumenta em animais submetidos ao jejum, diabetes, sepse, câncer, uremia, infecções e doenças inflamatórias causadas por endotoxemia e citocinas (Shiraki et al., 2005; Torres et al., 1998).
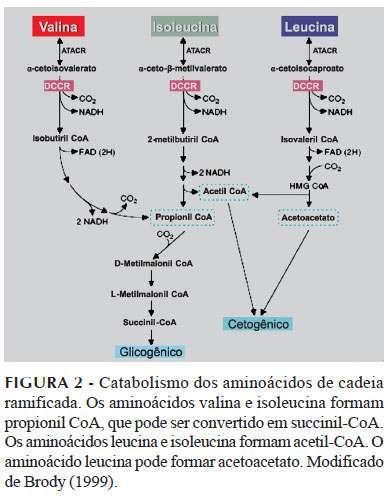
Posteriormente à segunda etapa do catabolismo dos ACR mediada pela DCCR, os produtos dessa reação - derivados de acil-CoA de cadeia ramificada - sofrem oxidação por meio de duas diferentes desidrogenases. Após essa etapa, as vias catabólicas de cada um dos ACR passam a divergir. A leucina é cetogênica, uma vez que forma acetil-CoA e acetoacetato, enquanto a valina é glicogênica, devido ao fato de ser convertida em succinil-CoA - intermediário do ciclo de Krebs. Tanto a isoleucina quanto a valina são metabolizadas para succinato via metilmalonil-CoA. O outro produto do metabolismo da isoleucina é o acetoacetato e, desse modo, a isoleucina pode ser considerada como um aminoácido glicogênico e cetogênico (Figura 2) (Brosnan, Brosnan, 2006; Harper, Miller, Block, 1984).
Aminoácidos de cadeia ramificada e regulação da síntese protéica muscular
ACR são essenciais na dieta e, portanto, relevantes na regulação da síntese protéica muscular. A administração endovenosa de glicose e de várias misturas de aminoácidos, por um período de uma hora, em ratos previamente privados de alimentação, demonstrou que a infusão de ACR e glicose aumenta a síntese protéica no músculo esquelético tão eficientemente quanto uma mistura contendo glicose e todos os aminoácidos. Esse fato sugere que o efeito anabólico de uma mistura completa de aminoácidos pode ser reproduzido pelo fornecimento de uma mistura contendo apenas os três ACR (Garlick, Grant, 1998). Contudo, o efeito da mistura dos três ACR sobre a síntese protéica muscular pode ser atribuído ao aminoácido leucina, uma vez que em estudo com músculo esquelético perfundido, foi verificado que o fornecimento de leucina isoladamente estimula a síntese protéica muscular tão efetivamente como a mistura dos três ACR (Li, Jefferson, 1978).
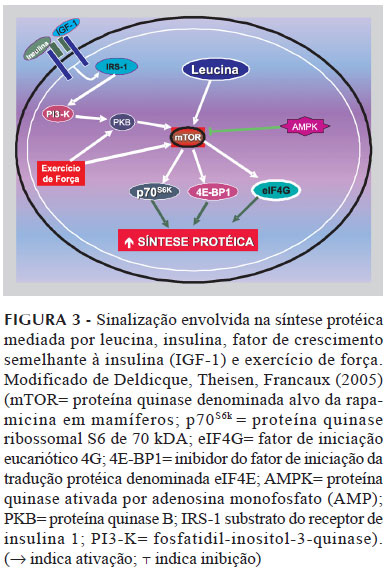
A leucina exerce os seus efeitos em nível pós-transcricional e mais comumente durante a fase de iniciação da tradução do RNA-mensageiro em proteína. O mecanismo pelo qual a leucina estimula a tradução de proteínas está relacionado ao fato do aumento da concentração intracelular desse aminoácido promover a ativação de uma proteína quinase denominada alvo da rapamicina em mamíferos (mammalian Target of Rapamycin - mTOR). O mTOR estimula a síntese protéica principalmente por meio de três proteínas regulatórias chaves: a proteína quinase ribossomal S6 de 70 kDA (p70S6k); a proteína 1 ligante do fator de iniciação eucariótico 4E (4E-BP1); e o fator de iniciação eucariótico 4G (eIF4G) (Figura 3) (Anthony et al., 2001; Anthony et al., 2000).
A 4E-BP1 é uma inibidora do fator de iniciação da tradução protéica conhecido como eIF4E. Quando a 4E-BP1 é fosforilada, o eIF4E é liberado e pode unir-se ao eIF4G - o qual está também sob o controle do mTOR - e ao eIF4A, o que forma o complexo eIF4F. A montagem desse complexo é necessária para a continuação da etapa de iniciação da tradução do RNA-mensageiro em proteína. A mTOR também ativa a p70S6k, que estimula a iniciação da tradução bem como a elongação da síntese protéica por diferentes mecanismos. A p70S6k,quando ativada, fosforila e inativa a enzima quinase do fator de elongação 2 (eEF2K), fato este que permite que o eEF2 seja ativado, o que promove a elongação. Consistente com esses fatos, a administração de leucina para ratos induz hiperfosforilação da 4E-BP1, promove formação do complexo eIF4F, causa hiperfosforilação da p70S6k e estimula a síntese protéica. Similarmente, dietas para ratos contendo 20% de proteína estimulam a síntese protéica hepática e muscular, que é associada ao aumento da fosforilação da 4E-BP1 e à conseqüente redução da ligação do eIF4E para a 4E-BP1, além do aumento da formação do complexo eIF4F. Esses fatos permitem relacionar a resposta anabólica sobre a síntese protéica muscular induzida pela ingestão de proteínas, por meio da capacidade do mTOR detectar alterações na concentração intracelular de leucina (Kimball, Jefferson, 2006a; Kimball, Jefferson, 2006b; Shah et al., 2000).
Leucina, insulina e síntese protéica muscular
A leucina influencia o controle de curto prazo da etapa de tradução da síntese protéica e este efeito é sinérgico com a insulina, que é um hormônio anabólico, com papel crítico na manutenção da síntese protéica muscular. Contudo, a insulina de modo isolado não é suficiente para estimular a síntese protéica muscular no estado pós-absortivo, sendo necessária a ingestão de proteínas ou de aminoácidos para restaurar completamente as taxas de síntese protéica. É proposto que o efeito da insulina na síntese protéica muscular esteja relacionado ao papel desse hormônio em potencializar o sistema de tradução de proteínas, ao invés de regular diretamente tal processo, ou seja, a insulina exerce um efeito permissivo sobre a síntese protéica na presença de aminoácidos (Norton, Layman, 2006). Aliado a isto, cabe ressaltar que a administração oral de leucina produz ligeiro e transitório aumento na concentração de insulina sérica, fato este que age também de modo permissivo para a estimulação da síntese protéica induzida por este aminoácido (Bolster, Jefferson, Kimball, 2004; Anthony et al., 2002a).
Em estudos sobre a interação entre os efeitos estimulatórios da leucina e da insulina sobre a síntese protéica no músculo esquelético, verifica-se que a administração de somatostatina - a qual inibe a secreção de insulina - atenua o aumento induzido pela leucina sobre a fosforilação da 4E-BP1 e da p70S6k, porém não tem efeito sobre a associação do eIF4E e eIF4G. Além disso, estudos em ratos diabéticos demonstram que parte da resposta da leucina sobre a síntese protéica no músculo esquelético ocorre tanto por meio de mecanismos independentes de insulina quanto dependentes de insulina (Kimball, Jefferson, 2006b). Portanto, conclui-se que os efeitos estimulatórios da leucina sobre a síntese protéica muscular ocorrem por mecanismos dependentes de insulina, que incluem a sinalização mediada pela proteína mTOR para a 4E-BP1 e a p70S6k, enquanto os efeitos independentes de insulina são mediados por um mecanismo ainda não totalmente esclarecido, que envolve a fosforilação do eIF4G e/ou sua associação com o eIF4E (Crozier et al., 2005; Deldicque, Theisen, Francaux, 2005; Anthony et al., 2002b).
METABOLISMO DE AMINOÁCIDOS DE CADEIA RAMIFICADA DURANTE O EXERCÍCIO
Durante o exercício físico ocorre a captação de diversos aminoácidos - predominantemente ACR - pelo tecido muscular. Se o exercício físico é prolongado, verifica-se significativa liberação de ACR pelo tecido hepático, aliada à diminuição da concentração plasmática de ACR - por exemplo, a concentração plasmática de leucina diminui entre 11 e 33% (Mero, 1999). Em um estudo (Decombaz et al., 1979) realizado com 11 homens treinados, submetidos a uma corrida com percurso de 100 km, foi verificada diminuição significativa (35-85%) da concentração sérica de ACR em relação aos valores pré-exercício. Em outro estudo com exercício prolongado (ciclismo, duração de 225 minutos, a 50% VO2max), Rennie et al. (1981) verificaram diminuição significativa da concentração plasmática de ACR ao final do exercício.
O músculo esquelético humano pode oxidar ao menos seis aminoácidos (leucina, isoleucina, valina, aspartato, glutamato e asparagina), todavia, durante o exercício físico, os ACR são preferencialmente oxidados (Wagenmakers, 1998). Os ACR são transaminados para os seus respectivos cetoácidos por meio da reação catalisada pela enzima ATACR, com subseqüente oxidação ocorrendo pelo complexo enzimático DCCR. O grupo amino dos ACR é transaminado com o α-cetoglutarato para formar glutamato, o qual é então transaminado com o piruvato - oriundo da via glicolítica - para formar alanina; ou aminado por meio da reação catalisada pela enzima glutamina sintetase, para formar glutamina (Rogero et al., 2006). A enzima DCCR é a enzima limitante do fluxo das reações envolvidas na oxidação dos ACR, com cerca de 5-8% na forma ativa (desfosforilada) no repouso e 20-25% na forma ativa durante o exercício (Tarnopolsky, 2004). A ativação da DCCR é relacionada à concentração de ACR e de cetoácidos de cadeia ramificada na fibra muscular, à depleção do glicogênio muscular e à diminuição do pH e da razão ATP:ADP (Shimomura et al., 1995). A correlação inversa entre ativação do complexo DCCR e concentração muscular de glicogênio sustenta o fato que estratégias de suplementação com carboidratos durante o exercício físico promovem efeito poupador da oxidação de aminoácidos por meio da diminuição da atividade do complexo DCCR. Cabe destacar que o aumento da ativação do complexo DCCR (e da oxidação de leucina) ocorre predominantemente durante o exercício intenso (70-80% VO2max) e prolongado, enquanto que em intensidades de exercício inferiores, o grau de ativação é reduzido (Xu et al., 2001; Shimomura et al., 2001).
O treinamento físico também influencia a oxidação de ACR e a ativação do complexo DCCR. Nesse contexto, McKenzie et al. (2000) investigaram a oxidação de leucina e a ativação do complexo DCCR durante 90 minutos de exercício, a 65% VO2max, antes e após 38 dias de treinamento de endurance em homens e mulheres. Tanto a oxidação de leucina quanto a atividade do complexo DCCR foram menores durante o exercício físico após o período de treinamento. Entretanto, a atividade total do complexo DCCR foi maior após o treinamento, o que sugere aumento da capacidade absoluta de oxidação dos ACR. Portanto, esses fatos indicam que o treinamento de endurance resulta em efeito poupador da oxidação de proteínas decorrente da redução da atividade do complexo DCCR, apesar do aumento da capacidade total de oxidação de ACR. Desse modo, essas adaptações induzidas pelo treinamento de endurance diminuiriam a contribuição de ACR para o fornecimento de energia; contudo, em período de estresse nutricional (baixa ingestão de energia ou de carboidratos) ou metabólico (treinamento exaustivo ou exercícios prolongados e intensos), a quantidade diária de oxidação de aminoácidos poderia exceder àquela observada em indivíduos sedentários ou em indivíduos com atividade física em nível recreacional.
Exercício de endurance, imunocompetência e aminoácidos de cadeia ramificada
O sistema imune é influenciado agudamente, e em menor extensão, cronicamente, pelo exercício. Dados epidemiológicos e experimentais sugerem que o exercício moderado aumenta a imunocompetência, enquanto que durante o treinamento intenso e após um evento competitivo ocorre aumento da incidência de infecções do trato respiratório superior (ITRS) em atletas. O exercício intenso e prolongado está associado com temporária imunossupressão que afeta macrófagos, neutrófilos e linfócitos. Os mecanismos envolvidos não estão completamente elucidados, porém são multifatoriais, incluindo ações hormonais - por exemplo, catecolaminas e cortisol -, inibição da síntese de citocinas por macrófagos e linfócitos T e diminuição da concentração plasmática de glutamina, que é o aminoácido livre mais abundante no plasma e no tecido muscular, e é utilizado em altas taxas por células de divisão rápida, incluindo leucócitos, para fornecer energia e favorecer a biossíntese de nucleotídeos. Uma vez que o exercício prolongado e intenso causa diminuição das concentrações plasmática e muscular de glutamina, esse fato pode repercutir sobre a imunocompetência do atleta, aumentando a incidência de ITRS (Nieman, Pedersen, 1999; Rogero, Tirapegui, 2000).
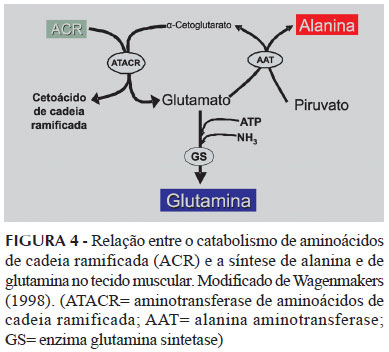
Os ACR podem atuar como precursores da síntese de glutamina no tecido muscular. Esses aminoácidos fornecem grupamentos amino em reações de transaminação, as quais acarretam na formação de glutamato que, posteriormente, na reação catalisada pela enzima glutamina sintetase, participa da síntese de glutamina (Rogero et al., 2006) (Figura 4). Nesse contexto, alguns estudos têm avaliado a efetividade da suplementação com ACR para manter a concentração plasmática de glutamina e modificar a resposta imune frente ao exercício de endurance exaustivo.
No que concerne ao estudo do efeito da suplementação com ACR durante o exercício exaustivo sobre a concentração plasmática de glutamina, Parry-Billings et al. (1992) avaliaram o efeito da suplementação com ACR (4 bebidas, contendo 4 g de ACR diluídos em 100 mL em cada bebida, totalizando 16 g de ACR), que foi oferecida após percorridos 10,5 km, 20,5 km, 32,5 km e 37,5 km ao longo de uma maratona (42,2 km) para indivíduos saudáveis. A suplementação com ACR promoveu aumento da concentração plasmática de ACR, ao mesmo tempo em que manteve a concentração plasmática de glutamina ao final da maratona. Por outro lado, o grupo placebo teve redução significativa da concentração plasmática de glutamina (16%) e de ACR (18%).
Bassit et al. (2000) avaliaram o efeito da suplementação com ACR sobre a resposta imune e a concentração plasmática de glutamina de triatletas, os quais realizaram um triatlo olímpico (natação 1,5 Km, ciclismo 40 km e corrida 10 Km). Os indivíduos foram distribuídos em grupo placebo ou grupo suplementado com ACR 30 dias antes da realização do triatlo. A suplementação com ACR (6 g/dia; leucina 60%, valina 20% e isoleucina 20%) foi ingerida durante os 30 dias que antecederam o triatlo. Uma dose de 3 g de ACR foi ingerida 30 minutos antes do triatlo, bem como uma dose de 3 g de ACR foi ingerida, pela manhã, durante os sete dias posteriores à realização do triatlo. Os autores verificaram que a concentração plasmática de glutamina após o triatlo foi mantida em relação aos valores basais no grupo suplementado com ACR, enquanto houve diminuição significativa no grupo placebo após o triatlo. Quanto à resposta imune, o grupo suplementado apresentou maior síntese in vitro de interleucina (IL)-1, IL-2, fator de necrose tumoral (TNF) e interferon (IFN)-α a partir de células mononucleares do sangue periférico estimuladas com mitógenos, no momento pós-triatlo, em relação ao grupo placebo. Além disso, a suplementação com ACR promoveu maior capacidade de proliferação de linfócitos obtidos do sangue periférico, quando estimulados com mitógenos, em relação ao grupo placebo tanto antes quanto após o triatlo. Paralelamente a esses efeitos, esse estudo também demonstrou redução da incidência de sintomas de infecção (34%) reportada pelos atletas suplementados com ACR no decorrer do período de suplementação - 30 dias antes e na semana posterior ao triatlo.
Desse modo, verifica-se que a manutenção da concentração plasmática de glutamina por meio da suplementação com ACR apresenta efeitos benéficos sobre a imunocompetência de atletas; todavia, estudos com suplementação de glutamina durante e após exercícios de endurance indicam que essa intervenção nutricional não previne a redução da imunocompetência induzida pelo exercício (Krzywkowski et al., 2001a; Krzywkowski et al., 2001b). Sendo assim, não está elucidado qual o mecanismo de ação da suplementação com ACR sobre a imunocompetência, ou seja, se é efeito decorrente da manutenção da concentração plasmática de glutamina, ou se é efeito direto dos ACR.
Exercício de endurance, hipótese da fadiga central e aminoácidos de cadeia ramificada
Fadiga durante o exercício físico pode estar relacionada tanto a fatores periféricos quanto centrais, os quais são influenciados pela intensidade e duração do exercício, ingestão de nutrientes e o nível de treinamento do indivíduo. Estudos têm sido publicados sobre a fadiga periférica e diversas alterações bioquímicas foram identificadas na etiologia dessa fadiga, como depleção da concentração muscular de glicogênio ou de creatina, acúmulo de prótons e falha na transmissão neuromuscular, enquanto fatores neurobiológicos relacionados à fadiga central são bem menos conhecidos. Durante o exercício prolongado de intensidade moderada, a diminuição da concentração de glicose decorrente da depleção dos estoques de glicogênio hepático é um dos fatores conhecidos que afeta o sistema nervoso central (SNC) e causa fadiga (Blomstrand, 2006).
Outro fator relacionado à etiologia da fadiga central está relacionado ao aumento da liberação de neurotransmissores, particularmente a 5-hidroxitriptamina (serotonina). Esta hipótese decorre do fato do exercício exaustivo resultar em gradual aumento da concentração de ácidos graxos livres no plasma, os quais competem com o aminoácido triptofano pela ligação à proteína plasmática albumina. Desse modo, observa-se aumento da concentração de triptofano livre, por meio do deslocamento deste aminoácido a partir da albumina plasmática. Cabe ressaltar que, em condições fisiológicas normais, 70-90% do triptofano que circula no sangue estão ligados à albumina plasmática, enquanto que 10-30% estão na forma livre. Além disso, o exercício intenso e prolongado acarreta em diminuição dos estoques de glicogênio muscular e hepático, o que desencadeia a utilização de outros substratos como fontes de energia, como os ACR, que são captados primariamente pelo tecido muscular e apresentam aumento da sua oxidação no decorrer do exercício físico (Fernstrom, Fernstrom, 2006; Davis, Alderson, Welsh, 2000).
Os ACR competem com o triptofano livre pela ligação ao mesmo transportador de aminoácidos neutros na barreira hemato-encefálica. Desse modo, a entrada de triptofano no sistema nervoso central (SNC) é regulada pela razão plasmática triptofano livre:ACR e favorecida pela diminuição da concentração de ACR no sangue, decorrente do aumento da sua taxa de oxidação. Sendo assim, a diminuição dos estoques de glicogênio, o aumento da oxidação de ACR e a elevação da concentração de ácidos graxos plasmáticos atuam como fatores relevantes no aumento da síntese do neurotransmissor serotonina no SNC, fato este dependente da disponibilidade de triptofano - precursor da serotonina - no SNC (Figura 5). O aumento da síntese de serotonina durante o exercício físico pode estar relacionado ao desenvolvimento da fadiga central, porquanto este neurotransmissor possui diversas funções fisiológicas, uma vez que atua no humor, letargia, comportamento individual, regulação do sono, da temperatura corporal e da pressão arterial, supressão do apetite e alterações na percepção de esforço físico (Blomstrand, 2006; Fernstrom, Fernstrom, 2006; Davis, Alderson, Welsh, 2000).
Tem sido proposto que a ingestão de ACR durante o exercício prolongado poderia atenuar o aumento da razão entre as concentrações de triptofano livre e ACR, o que retardaria o início da fadiga central e, conseqüentemente, aumentaria a performance. Contudo, um potencial efeito prejudicial da suplementação com ACR, dependendo da dose administrada, é o aumento das concentrações plasmática e muscular de amônia, que atua como agente de fadiga. Além disso, de acordo com diversos estudos, evidencia-se que a suplementação com ACR não aumenta a performance quando comparada à suplementação de carboidratos (Calders, Matthys, Derave, 1999; Davis et al., 1999; van Hall et al., 1995).
Dentre os estudos que investigaram o efeito da suplementação com ACR e a hipótese da fadiga central, destaca-se a pesquisa de van Hall et al. (1995), no qual 10 atletas de endurance foram avaliados durante exercício em ciclo ergômetro, realizado em intensidade entre 70 e 75% da força máxima, enquanto ingeriram bebidas contendo 6% de sacarose, 6% de sacarose acrescida de triptofano (3 g por litro), 6% de sacarose e baixa dose de ACR (6 g por litro) ou 6% de sacarose e alta dose de ACR (18 g por litro). O estudo foi delineado de modo randomizado e duplo cego. O tempo de tolerância ao esforço (122 ± 3 minutos) não diferiu entre os quatro grupos avaliados, apesar de os tratamentos terem significativamente aumentado a concentração plasmática do respectivo aminoácido suplementado. Nesse estudo, foi calculada a taxa de transporte unidirecional de triptofano através da barreira hematoencefálica (influxo), sendo estimado que a suplementação com ACR reduziu a captação de triptofano na exaustão em cerca de 8-12%, enquanto a ingestão de triptofano causou aumento da sua captação pelo cérebro de 7-20 vezes. Desse modo, esses resultados indicam que a manipulação do fornecimento de triptofano para o SNC não têm efeito adicional sobre a atividade serotoninérgica durante o exercício prolongado e exaustivo ou que a manipulação da atividade serotoninérgica funcionalmente não contribui para os mecanismos de fadiga.
Davis et al. (1999) verificaram o efeito da suplementação com ACR sobre a performance durante a realização de exercício (corrida) de alta intensidade e intermitente. Para tanto, indivíduos ingeriram, 1 hora antes do exercício, bebidas contendo carboidrato (5 mL/kg; concentração de 20% de carboidrato) (grupo CHO), ou a mesma bebida adicionada com 7 g de ACR (grupo CHO+ACR), ou água flavorizada (grupo P). Imediatamente antes do início do exercício, os indivíduos do grupo CHO ingeriram bebida contendo carboidrato (5 mL/kg; concentração de 6% de carboidrato); o grupo CHO+ACR ingeriu essa mesma bebida adicionada de 7 g de ACR, enquanto o grupo P ingeriu água flavorizada. O tempo de tolerância ao esforço e as concentrações de glicose e de insulina foram maiores nos grupos CHO e CHO+ACR em relação ao grupo P, o que confirma o efeito benéfico da suplementação com carboidratos sobre a performance em protocolo de exercício com modelo de atividade similar àquele observado em esportes, tais como futebol, basquete e hóquei. Entretanto, esse estudo não sustenta a hipótese de benefício adicional da suplementação com ACR sobre a performance.
Portanto, diante dos estudos relacionados à performance, fadiga central e suplementação de nutrientes, é fundamental destacar que a suplementação com carboidratos pode ser uma estratégia mais efetiva, uma vez que atenua tanto o aumento da concentração plasmática de ácidos graxos livres quanto da razão plasmática triptofano livre:ACR durante o exercício prolongado e exaustivo (Davis, Alderson, Welsh, 2000). Além disso, a ingestão de carboidratos também reduz o aumento da concentração de amônia no plasma e no tecido muscular durante o exercício físico, aliado ao fato da inclusão de ACR em soluções contendo carboidratos não acarretar em benefícios adicionais sobre a performance.
Exercício de endurance em ambientes quentes, performance e aminoácidos de cadeia ramificada
A diminuição da performance em exercícios de endurance realizados em ambientes quentes tem sido atribuída à redução do volume plasmático ou sangüíneo, ao aumento da taxa de glicogenólise muscular, à redução do fluxo sangüíneo muscular e ao acúmulo de lactato no sangue. Contudo, tem sido proposto que a fase inicial da fadiga durante o exercício de endurance realizado em ambiente quente, quando comparado ao mesmo exercício realizado em ambiente frio, não está associada com os fatores acima citados. Desse modo, tem sido sugerido que o estresse térmico per se pode acelerar a ocorrência da fadiga por meio da redução da função de centros motores no cérebro, alteração do recrutamento de unidades motoras ou diminuição da motivação para a atividade. A observação desses fatos, aliada ao conhecimento que ACR estão relacionados com a hipótese da fadiga central, têm acarretado em estudos que visam avaliar o efeito da suplementação com ACR sobre a performance de indivíduos submetidos a exercício de endurance em ambientes quentes (Nielsen et al., 1990; Savard et al., 1988).
Nesse contexto, Mittleman et al. (1998) investigaram o efeito da suplementação com ACR sobre a performance em indivíduos submetidos a exercício de endurance em ciclo ergômetro, em intensidade de 40% VO2max e temperatura ambiente de 34,4 ºC. Os indivíduos ingeriram 5 mL/kg de massa corporal de uma bebida contendo 5,88 g/L de ACR (54% leucina, 19% isoleucina e 27% valina) ou 5,88 g/L de polidextrose, a cada 60 minutos, durante o repouso, e a cada 30 minutos, durante o exercício. Aumento de 12% no tempo de tolerância ao esforço físico foi verificado após a suplementação com ACR comparado ao grupo placebo, aliado à redução de 2-3 vezes na razão plasmática triptofano livre: ACR. Por outro lado, Watson et al. (2004) verificaram que a suplementação com ACR - aproximadamente 20 g de ACR consumidos antes e durante o exercício - não teve qualquer efeito sobre a performance ou sobre os escores de percepção de esforço em indivíduos submetidos a exercício de endurance (ciclismo) em ambiente quente (30 ºC) e intensidade de 50% VO2max.
Exercício de força, balanço protéico muscular e aminoácidos de cadeia ramificada
A hipertrofia muscular ocorre apenas a partir do saldo de síntese de proteínas, ou seja, quando a síntese protéica muscular excede a degradação protéica muscular (Rennie, Tipton, 2000). É notório que o exercício, especialmente o exercício de força, tem profundo efeito sobre o metabolismo protéico muscular, freqüentemente resultando em crescimento muscular. Agudamente, o exercício de força pode resultar em melhora do balanço protéico muscular (síntese - degradação); porém, na ausência da ingestão de alimentos, o balanço ainda permanece negativo. Portanto, os efeitos interativos entre o exercício de força e as diferentes estratégias nutricionais devem ser considerados no estudo do metabolismo protéico muscular. Nesse contexto, verifica-se que a ingestão de aminoácidos isoladamente aumenta a taxa de síntese protéica muscular. Contudo, o mais potente iniciador dessa síntese é a combinação de exercício de força com aumento da disponibilidade de aminoácidos (Tipton, Wolfe, 2004; Houston, 1999).
A ingestão de uma mistura de aminoácidos ou de um hidrolisado de proteínas após uma sessão de exercício de força estimula a taxa de síntese protéica em músculo humano e promove balanço protéico muscular positivo. Diferentes teorias tentam explicar a ocorrência desse efeito, como o aumento da disponibilidade de aminoácidos promovendo o aumento do transporte dos mesmos para dentro da célula muscular, o que estimula a síntese protéica (Tipton et al., 2004; Miller et al., 2003). Outra possibilidade é que esse efeito decorre de um grupo de aminoácidos, como os ACR, ou de um único aminoácido, como a leucina. No que concerne à leucina, esta aumenta a fosforilação de proteínas envolvidas na regulação da síntese protéica, incluindo a p70S6k e a 4E-BP1, no músculo esquelético de humanos. Aliado a esse fato, observa-se que a atividade da p70S6k induzida pelo exercício correlaciona-se com o aumento da massa muscular após seis semanas de treinamento de força. Desse modo, alterações na fosforilação da p70S6k no músculo esquelético pós-exercício podem refletir em ativação de vias de sinalização, as quais podem responder pelo aumento da síntese protéica durante a fase inicial da recuperação pós-exercício. Esse fato é relevante, uma vez que a ingestão de leucina aumenta a fosforilação de proteínas envolvidas na regulação da síntese protéica muscular, incluindo a p70S6k (Figura 3) (Blomstrand et al., 2006; Kimball, Jefferson, 2006a; Norton, Layman, 2006; Anthony et al., 2000).
Koopman et al. (2005) verificaram que a adição de leucina em bebida contendo hidrolisado protéico e carboidratos promoveu maior estimulação da síntese protéica corporal total após a realização de uma sessão de exercício de força quando comparada à ingestão de carboidrato ou de carboidrato com hidrolisado protéico. Além disso, a ingestão combinada de carboidrato, hidrolisado protéico e leucina aumentou a síntese protéica muscular em relação à ingestão isolada de carboidrato. Os resultados desse estudo indicam que a adição de leucina na forma livre em combinação com proteínas e carboidratos representa uma estratégia efetiva na promoção do anabolismo protéico muscular pós-exercício de força.
Karlsson et al. (2004) investigaram o efeito do exercício de força isolado ou em combinação com a ingestão oral de ACR sobre a fosforilação da p70S6k no músculo esquelético. Sete indivíduos executaram uma sessão de exercício de força (músculo quadríceps; 4 x 10 repetições; 80% de uma repetição máxima) em duas condições, ou seja, com a ingestão de solução contendo ACR (45% leucina, 30% valina e 25% isoleucina) ou placebo (água flavorizada) durante e após o exercício. A ingestão de ACR acarretou no aumento da concentração plasmática dos três ACR durante o exercício e o período de recuperação (2 horas). O exercício de força promoveu significativo aumento da fosforilação da p70S6k, que persistiu 1 e 2 horas pós-exercício, enquanto a ingestão com ACR aumentou 3,5 vezes a fosforilação da p70S6k durante a recuperação. Além disso, a fosforilação da proteína ribossomal S6 - substrato da p70S6k - foi aumentada durante o período de recuperação pós-exercício de força apenas no grupo que ingeriu ACR. Desse modo, ACR - ingeridos durante e após o exercício de força - podem aumentar a síntese protéica no músculo esquelético pós-exercício de força por meio da cascata de sinalização dependente da p70S6k.
Lesão muscular induzida pelo exercício físico e aminoácidos de cadeia ramificada
Shimomura et al. (2006a) investigaram em doze mulheres saudáveis os efeitos da suplementação com ACR sobre a dor muscular de início tardio (DMIT) e a fadiga muscular induzidas pelo exercício. A composição das soluções testes utilizadas foi: (i) solução ACR (200 mL) que continha 5,5 g de ACR (isoleucina:leucina:valina = 1:2,3:1,2); e (ii) solução placebo (200 mL) contendo 5,5 g de dextrina em substituição aos ACR. Na manhã do exercício físico, os indivíduos ingeriram a solução de ACR (0,1 g/kg de peso) ou a solução de dextrina (0,1 g/kg de peso) 15 minutos antes do exercício de agachamento, que consistiu em 7 séries, de 20 agachamentos/série, com 3 minutos de intervalo entre cada série. Durante cada série, os agachamentos foram realizados a cada 2 segundos. A dor muscular foi maior nos 2º e 3º dias no grupo placebo, indicando a ocorrência de DMIT. Contudo, apesar da DMIT também ter ocorrido no grupo suplementado com ACR, o pico de dor ocorreu apenas no 2º dia e foi significativamente menor em relação àquele observado no grupo controle. A DMIT entre o 3º e o 5º dias foi também significativamente menor no grupo ACR em comparação ao grupo placebo. Portanto, os resultados obtidos nesse estudo demonstram que a ingestão de 5 g de ACR previamente ao exercício físico pode reduzir a DMIT e a fadiga muscular por diversos dias pós-exercício. Dentre os possíveis mecanismos relacionados a esses resultados destacam-se a possibilidade dos ACR atenuarem a degradação protéica pós-exercício e o fato da leucina poder estimular a síntese protéica muscular.
Em outro estudo (Coombes, Mcnaughton, 2000) foi avaliado o efeito da suplementação com ACR sobre a concentração sérica de creatina quinase (CK) e lactato desidrogenase (LDH) - parâmetros indicativos de lesão muscular - após a realização de exercício prolongado. Para tanto, 16 homens foram distribuídos em dois grupos, sendo um grupo suplementado com 12 g de ACR por dia, durante 14 dias, juntamente com a dieta normal, e um grupo controle (dieta normal apenas). O teste de exercício físico foi realizado no 7º dia do estudo e consistiu de exercício de ciclismo realizado em ciclo ergômetro, em intensidade de aproximadamente 70% VO2max. As amostras de sangue foram coletadas uma semana antes do teste (ciclismo) e 1, 2, 3 e 4 horas, 1, 3, 5 e 7 dias após o exercício. Os valores basais de CK e LDH não diferiram entre os grupos 7 dias previamente ao teste. Contudo, verificou-se significativo aumento entre os valores pré-exercício e pós-exercício para LDH e CK até 5 dias após o exercício. Cabe ressaltar que a suplementação com ACR, significativamente, reduziu essa alteração na concentração de LDH entre 2 h e 5 dias pós-exercício e de CK entre 4 h e 5 dias pós-exercício, o que indica que a suplementação com ACR pode reduzir a lesão muscular associada com o exercício de endurance.
CONSIDERAÇÕES FINAIS
A suplementação com ACR estimula o processo de tradução de RNA mensageiros específicos no tecido muscular durante o período de recuperação pós-exercício de força; contudo, há escassez de correspondentes estudos sobre o efeito da suplementação com ACR em humanos submetidos a exercícios de endurance. Não existem evidências comprovando o efeito dos ACR no aumento da performance em indivíduos engajados em exercícios de endurance, todavia, essa intervenção nutricional pode atenuar a lesão muscular induzida pelo exercício. Além disso, estudos são necessários para elucidar qual o mecanismo de ação da suplementação com ACR sobre a imunocompetência de atletas.
Recebido para publicação em 06 de setembro de 2007
Aceito para publicação em 02 de outubro de 2008
- ANTHONY, J.C.; ANTHONY, T.G.; KIMBALL, S.R.; JEFFERSON, L.S. Signaling pathways involved in translational control of protein synthesis in skeletal muscle by leucine. J. Nutr., v.131, n.3, p.856S-860S, 2001.
- ANTHONY, J.C.; LANG, C.H.; CROZIER, S.J.; ANTHONY, T.G.; MACLEAN, D.A.; KIMBALL, S.R.; JEFFERSON, L.S. Contribution of insulin to the translational control of protein synthesis in skeletal muscle by leucine. Am. J. Physiol. Endocrinol. Metab., v.282, n.5, p.E1092-E1101, 2002a.
- ANTHONY, J.C.; REITER, A.K.; ANTHONY, T.G.; CROZIER, S.J.; LANG, C.H.; MACLEAN, D.A. KIMBALL, S.R.; JEFFERSON, L.S. Orally administered leucine enhances protein synthesis in skeletal muscle of diabetic rats in the absence of increases in 4E-BP1 or S6K1 phosphorylation. Diabetes, v.51, n.4, p.928-936, 2002b.
- ANTHONY, J.C.; YOSHIZAWA, F.; ANTHONY, T.G.; VARY, T.C.; JEFFERSON, L.S.; KIMBALL, S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr., v.130, n.10, p.2413-2419, 2000.
- BASSIT, R.A.; SAWADA, L.A.; BACURAU, R.F.; NAVARRO, F.; COSTA ROSA, L.F.The effect of BCAA supplementation upon the immune response of triathletes. Med. Sci. Sports Exerc., v.32, n.7, p.1214-1219, 2000.
- BLOMSTRAND, E. A role for branched-chain amino acids in reducing central fatigue. J. Nutr., v.136, n.2, p.544S-547S, 2006.
- BLOMSTRAND, E.; ELIASSON, J.; KARLSSON, H.K.; KOHNKE, R. Branched-chain amino acids activate key enzymes in protein synthesis after physical exercise. J. Nutr., v.136, p.269S-273S, 2006.
- BOLSTER, D.R.; JEFFERSON, L.S.; KIMBALL, S.R. Regulation of protein synthesis associated with skeletal muscle hypertrophy by insulin-, amino acid- and exercise-induced signalling. Proc. Nutr. Soc., v.63, n.2, p.351-356, 2004.
- BRODY, T. Nutritional biochemistry. 2. ed. San Diego: Academic Press, 1999, 658p.
- BROSNAN, J.T.; BROSNAN, M.E. Branched-chain amino acids: enzyme and substrate regulation. J. Nutr., v.136, p.207S-211S, 2006.
- CALDERS, P.; MATTHYS, D.; DERAVE, W. Effect of branched-chain amino acids (BCAA), glucose, and glucose plus BCAA on endurance performance in rats. Med. Sci. Sports Exerc., v.31, n.4, p.583-587, 1999.
- COOMBES, J.S.; MCNAUGHTON, L.R. Effects of branched-chain amino acid supplementation on serum creatine kinase and lactate dehydrogenase after prolonged exercise. J. Sports Med. Phys. Fitness, v.40, n.3, p.240-246, 2000.
- CROZIER, S.J.; KIMBALL, S.R.; EMMERT, S.W.; ANTHONY, J.C.; JEFFERSON, L.S. Oral leucine administration stimulates protein synthesis in rat skeletal muscle. J. Nutr., v.135, n.3, p.376-382, 2005.
- CYNOBER, L.; HARRIS, R.A. Symposium on branched-chain amino acids: conference summary. J. Nutr., v.136, p.333S-336S, 2006.
- DAVIS, J.M.; ALDERSON, N.L.; WELSH, R.S. Serotonin and central nervous system fatigue: nutritional considerations. Am. J .Clin. Nutr., v.72, p.573S-578S, 2000.
- DAVIS, J.M.; WELSH, R.S.; DE VOLVE, K.L.; ALDERSON, N.A. Effects of branched-chain amino acids and carbohydrate on fatigue during intermittent, high-intensity running. Int. J. Sports Med., v.20, n.5, p.309-314, 1999.
- DECOMBAZ, J.; REINHARDT, P.; ANANTHARAMAN, K.; VON GLUTZ, G.; POORTMANS, J.R. Biochemical changes in a 100 km run: free amino acids, urea, and creatinine. Eur. J. Appl. Physiol. Occup. Physiol., v.41, n.1, p.61-72, 1979.
- DELDICQUE, L.; THEISEN, D.; FRANCAUX, M. Regulation of mTOR by amino acids and resistance exercise in skeletal muscle. Eur. J. Appl. Physiol., v.94, n.1-2, p.1-10, 2005.
- FERNSTROM, J.D.; FERNSTROM, M.H. Exercise, serum free tryptophan, and central fatigue. J. Nutr., v.136, n.2, p.553S-559S, 2006.
- GARLICK, P.J.; GRANT, I. Amino acid infusion increases the sensitivity of muscle protein synthesis in vivo to insulin. Effect of branched-chain amino acids. Biochem. J., v.254, n.2, p.579-584, 1998.
- HARPER, A.E.; MILLER, R.H.; BLOCK, K.P. Branched-chain amino acid metabolism. Annu. Rev. Nutr., v.4, p.409-454, 1984.
- HARRIS, R.A.; JOSHI, M.; JEOUNG, N.H. Mechanisms responsible for regulation of branched-chain amino acid catabolism. Biochem. Biophys. Res. Commun., v.313, n.2, p.391-396, 2004.
- HARRIS, R.A.; JOSHI, M.; JEOUNG, N.H.; OBAYASHI, M. Overview of the molecular and biochemical basis of branched-chain amino acid catabolism. J. Nutr., v.135, p.1527S-1530S, 2005.
- HOUSTON, M.E. Gaining weight: the scientific basis of increasing skeletal muscle mass. Can. J. Appl. Physiol., v.24, n.4, p.305-316, 1999.
- HUTSON, S.M.; SWEATT, A.J.; LANOUE, K.F. Branched-chain amino acid metabolism: implications for establishing safe intakes. J. Nutr., v.135, p.1557S-1564S, 2005.
- KARLSSON, H.K.; NILSSON, P.A.; NILSSON, J.; CHIBALIN, A.V.; ZIERATH, J.R.; BLOMSTRAND, E. Branched-chain amino acids increase p70S6k phosphorylation in human skeletal muscle after resistance exercise. Am. J. Physiol. Endocrinol. Metab., v.287, n.1, p.E1-E7, 2004.
- KIMBALL, S.R.; JEFFERSON, L.S. New functions for amino acids: effects on gene transcription and translation. Am. J. Clin. Nutr., v.83, n.2, p.500S-507S, 2006a.
- KIMBALL, S.R.; JEFFERSON, L.S. Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J. Nutr., v.136, p.227S-231S, 2006b.
- KOOPMAN, R.; WAGENMAKERS, A.J.; MANDERS, R.J.; ZORENC, A.H.; SENDEN, J.M.; GORSELINK, M.; KEIZER, H.A.; VAN LOON, L.J. Combined ingestion of protein and free leucine with carbohydrate increases postexercise muscle protein synthesis in vivo in male subjects. Am. J. Physiol. Endocrinol. Metab., v.288, n.4, p.E645-E653, 2005.
- KRZYWKOWSKI, K.; PETERSEN, E.W.; OSTROWSKI, K.; AMSTER, H.L.; BOZA, J.; KRISTENSEN, J.H.; PEDERSEN, B.K. Effect of glutamine supplementation and protein supplementation on exercise-induced decreases in salivary IgA. J. Appl. Physiol., v.91, p.832-838, 2001a.
- KRZYWKOWSKI, K.; PETERSEN, E.W.; OSTROWSKI, K.; KRISTENSEN, J.H.; BOZA, J.; PEDERSEN, B.K. Effect of glutamine supplementation on exercise-induced changes in lymphocyte function. Am. J. Physiol. Cell Physiol., v.281, p.C1259-C1265, 2001b.
- LI, J.B.; JEFFERSON, L.S. Influence of amino acid availability on protein turnover in perfused skeletal muscle. Biochim. Biophys. Acta., v.544, n.2, p.351-359, 1978.
- MARCHINI, J.S.; MORIGUTI, J.C.; PADOVAN, G.J.; NONINO, C.B.; VIANNA, S.M.L.; OLIVEIRA, J.E.D. Métodos atuais de investigação do metabolismo protéico: Aspectos básicos e estudos experimentais e clínicos. Medicina, v.31, n.1, p.22-30, 1998.
- MCKENZIE, S.; PHILLIPS, S.M.; CARTER, S.L.; LOWTHER, S.; GIBALA, M.J.; TARNOPOLSKY, M.A. Endurance exercise training attenuates leucine oxidation and BCOAD activation during exercise in humans. Am. J. Physiol. Endocrinol. Metab., v.278, n.4, p.E580-E587, 2000.
- MERO, A. Leucine supplementation and intensive training. Sports Med., v.27, n.6, p.347-358, 1999.
- MILLER, S.L.; TIPTON, K.D.; CHINKES, D.L.; WOLF, S.E.; WOLFE, R.R. Independent and combined effects of amino acid and glucose after resistance exercise. Med. Sci. Sports Exerc., v.35, n.3, p.449-455, 2003.
- MITTLEMAN, K.D.; RICCI, M.R.; BAILEY, S.P. Branched-chain amino acids prolong exercise during heat stress in men and women. Med. Sci. Sports Exerc., v.30, n.1, p.83-91, 1998.
- NIELSEN, B.; SAVARD, G.; RICHTER, E.A.; HARGREAVES, M.; SALTIN, B. Muscle blood flow and muscle metabolism during exercise and heat stress. J. Appl. Physiol., v.69, n.3, p.1040-1046, 1990.
- NIEMAN, D.C.; PEDERSEN, B.K. Exercise and immune function. Recent developments. Sports Med., v.27, n.2, p.73-80, 1999.
- NORTON, L.E.; LAYMAN, D.K. Leucine regulates translation initiation of protein synthesis in skeletal muscle after exercise. J. Nutr., v.136, n.2, p.533S-537S, 2006.
- PARRY-BILLINGS, M.; BUDGETT, R.; KOUTEDAKIS, Y.; BLOMSTRAND, E.; BROOKS, S.; WILLIAMS, C.; CALDER, P.C.; PILLING, S.; BAIGRIE, R.; NEWSHOLME, E.A. Plasma amino acid concentration in the overtraining syndrome: possible effects on the immune system. Med. Sci. Sport. Exerc., v.24, n.12, p.1353-1358, 1992.
- RENNIE, M.J.; EDWARDS, R.H.T.; KRYWAWYCH, S.; DAVIES, C.T.; HALLIDAY, D.; WATERLOW, J.C.; MILLWARD, D.J. Effect of exercise on protein turnover in man. Clin. Sci., v.61, n.5, p.627-639, 1981.
- RENNIE, M.J.; TIPTON, K.D. Protein and amino acid metabolism during and after exercise and the effects of nutrition. Annu. Rev. Nutr., v.20, p.457-483, 2000.
- ROGERO, M.M.; TIRAPEGUI, J. Aspectos atuais sobre glutamina, atividade física e sistema imune. Rev. Bras. Cien. Farm., v.36, n.2, p.201-212, 2000.
- ROGERO, M.M.; TIRAPEGUI, J.; PEDROSA, R.G.; CASTRO, I.A.; PIRES, I.S. Effect of alanyl-glutamine supplementation on plasma and tissue glutamine concentrations in rats submitted to exhaustive exercise. Nutrition, v.22, n.5, p.564-571, 2006.
- SAVARD, G.K.; NEILSEN, B.; LASZCZYNSKA, I.; LARSEN, B.E.; SALTIN, B. Muscle blood flow is not reduced in humans during moderate exercise and heat stress. J. Appl. Physiol., v.64, n.2, p.649-657, 1988.
- SHAH, O.J.; ANTHONY, J.C.; KIMBALL, S.R.; JEFFERSON, L.S. 4E-BP1 and S6K1: translational integration sites for nutritional and hormonal information in muscle. Am. J. Physiol. Endocrinol. Metab., v.279, n.4, p.E715-E729, 2000.
- SHIMOMURA, Y.; FUJII, H.; SUZUKI, M.; MURAKAMI, T.; FUJITSUKA, N.; NAKAI, N. Branched-chain alpha-keto acid dehydrogenase complex in rat skeletal muscle: regulation of the activity and gene expression by nutrition and physical exercise. J. Nutr., v.125, n. p.1762S-1765S, 1995.
- SHIMOMURA, Y.; HARRIS, R.A. Metabolism and physiological function of branched-chain amino acids: discussion of session 1.J. Nutr., v.136, n. p.232S-233S, 2006.
- SHIMOMURA, Y.; HONDA, T.; SHIRAKI, M.; MURAKAMI, T.; SATO, J.; KOBAYASHI, H.; MAWATARI, K.; OBAYASHI, M.; HARRIS, R.A. Branched-chain amino acid catabolism in exercise and liver disease. J. Nutr., v.136, n. p.250S-253S, 2006b.
- SHIMOMURA, Y.; MURAKAMI, T.; NAKAI, N.; NAGASAKI, M.; HARRIS, R.A. Exercise promotes BCAAcatabolism: effects of BCAA supplementation on skeletal muscle during exercise. J. Nutr., v.134, n. p.1583S-1587S, 2004.
- SHIMOMURA, Y.; OBAYASHI, M.; MURAKAMI, T.; HARRIS, R.A. Regulation of branched-chain amino acid catabolism: nutritional and hormonal regulation of activity and expression of the branched-chain alpha-keto acid dehydrogenase kinase. Curr. Opin. Clin. Nutr. Metab. Care, v.4, n.5, p.419-423, 2001.
- SHIMOMURA, Y.; YAMAMOTO, Y.; BAJOTTO, G.; SATO, J.; MURAKAMI, T.; SHIMOMURA, N.; KOBAYASHI, H.; MAWATARI, K. Nutraceutical effects of branched-chain amino acids on skeletal muscle. J. Nutr., v.136, n.2, p.529S-532S, 2006a.
- SHIRAKI, M.; SHIMOMURA, Y.; MIWA, Y.; FUKUSHIMA, H.; MURAKAMI, T.; TAMURA, T.; MORIWAKI, H. Activation of hepatic branched-chain alpha-keto acid dehydrogenase complex by tumor necrosis factor-alpha in rats. Biochem. Biophys. Res. Commun., v.328, n.4, p.973-978, 2005.
- TARNOPOLSKY, M. Protein requirements for endurance athletes. Nutrition, v.20, n.7-8, p.662-668, 2004.
- TIPTON, K.D.; ELLIOTT, T.A.; CREE, M.G.; WOLF, S.E.; SANFORD, A.P.; WOLFE, R.R. Ingestion of casein and whey proteins result in muscle anabolism after resistance exercise. Med. Sci. Sports Exerc., v.36, n.12, p.2073-2081, 2004.
- TIPTON. K.D.; WOLFE, R.R. Protein and amino acids for athletes. J. Sports Sci., v.22, n.1, p.65-79, 2004.
- TOM, A.; NAIR, K.S. Assessment of branched-chain amino acid status and potential for biomarkers. J. Nutr., v.136, n. p.324S-330S, 2006.
- TORRES, N.; LOPEZ, G.; DE SANTIAGO, S.; HUTSON, S.M.; TOVAR, A.R. Dietary protein level regulates expression of the mitochondrial branched-chain aminotransferase in rats. J. Nutr., v.128, n.8, p.1368-1375, 1998.
- VAN HALL, G.; RAAYMAKERS, J.S.; SARIS, W.H.; WAGENMAKERS, AJ. Ingestion of branched-chain amino acids and tryptophan during sustained exercise in man: failure to affect performance. J. Physiol., v.486, n. p.789-794, 1995.
- WAGENMAKERS, A.J. Muscle amino acid metabolism at rest and during exercise: role in human physiology and metabolism. Exerc. Sport Sci. Rev., v.26, n. p.287-314, 1998.
- WATSON, P.; SHIRREFFS, S.M.; MAUGHAN, R.J. The effect of acute branched-chain amino acid supplementation on prolonged exercise capacity in a warm environment. Eur. J. Appl. Physiol., v.93, n.3, p.306-314, 2004.
- XU, M.; NAGASAKI, M.; OBAYASHI, M.; SATO, Y.; TAMURA, T.; SHIMOMURA, Y. Mechanism of activation of branched-chain alpha-keto acid dehydrogenase complex by exercise. Biochem. Biophys. Res. Commun, v.287, n.3, p.752-756, 2001.
Datas de Publicação
-
Publicação nesta coleção
27 Jan 2009 -
Data do Fascículo
Dez 2008
Histórico
-
Aceito
02 Out 2008 -
Recebido
06 Set 2007