Abstract
A suitable chemically defined culture medium was selected and some optimal conditions for the production of the highly immunosuppressive compound, cyclosporin A (Cyc A) are reported. Medium of the following composition was favorable for the production of Cyc A by Fusarium roseum: glucose, 30; NaNO3, 2; KH2PO4, 1; MgSO4.7H2O, 0.5 and KCL, 0.5 (g/l). Maximum productivity of Cyc A was achieved at pH 6.0 when 50 ml of the fermentation medium/250 ml flask, inoculated with five fungal agar discs (6 mm, diameter) of 7-days old F. roseum culture after incubation at 30 ºC at 120 rpm for 7 days.
Fusarium roseum; Cyclosporin A; Fermentation conditions
MICROBIAL PHYSIOLOGY
Some optimal culture conditions for production of cyclosporin a by Fusarium roseum
Ahmed A. Ismaiel* * Corresponding Author. Mailing address: Botany Depart., Faculty of Science, Zagazig University, Zagazig, Egypt..; E-mail: microbiologist_80@yahoo.com ; El-Sayed A. El-Sayed; Asmaa A. Mahmoud
Department of Botany, Faculty of Science, Zagazig University, Zagazig, Egypt
ABSTRACT
A suitable chemically defined culture medium was selected and some optimal conditions for the production of the highly immunosuppressive compound, cyclosporin A (Cyc A) are reported. Medium of the following composition was favorable for the production of Cyc A by Fusarium roseum: glucose, 30; NaNO3, 2; KH2PO4, 1; MgSO4.7H2O, 0.5 and KCL, 0.5 (g/l). Maximum productivity of Cyc A was achieved at pH 6.0 when 50 ml of the fermentation medium/250 ml flask, inoculated with five fungal agar discs (6 mm, diameter) of 7-days old F. roseum culture after incubation at 30 ºC at 120 rpm for 7 days.
Key words:Fusarium roseum, Cyclosporin A, Fermentation conditions.
INTRODUCTION
Cyclosporins are a family of neutral, highly lipophilic, cyclic undecapeptides containing unusual amino acids (36, 37). They can be produced in a fungal fermentation process by aerobic filamentous fungi which were originally classified as Trichoderma polysporum (Link ex Pers.) Rifai (12) and more recently as Tolypocladium inflatum W. Gams (16) or Beauveria nivea Von Arx (35).
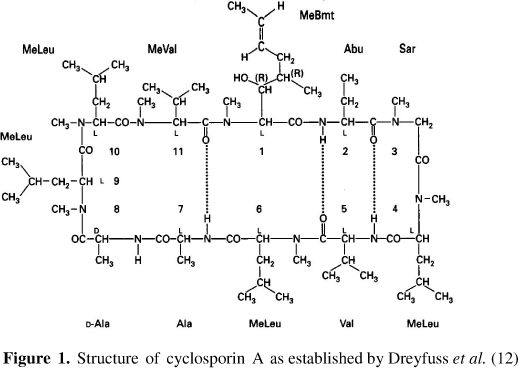
Cyclosporin A (Cyc A, Figure 1), the main representative of the series, is a potent antifungal and immunosuppressive compound (7, 17) that has been widely used to diminish transplant rejection (6) and control some autoimmune diseases (33).
Cyc A biosynthesis is catalyzed by a single multienzyme polypeptide (20), the so-called cyclosporin synthetase which has a molecular mass of about 1400 KDa (30). This enzyme synthesizes Cyc A by a thiotemplate mechanism starting from the precursor amino acids in their unmethylated form (20).
Many attempts have been made to optimize Cyc A production, including immobilization (32) and solid state fermentation (26). However, even though the enzymatic production of cyclosporin has already been established and proven (5), submerged fermentation production is normally used, owing to the complexity involved in enzymatic synthesis. The organisms that are known to produce Cyc A include Tolypocladium inflatum (14), Fusarium solani (29) Neocosmospora varinfecta (24) and Aspergillus terreus (28). In this paper, we briefly describe the optimum fermentation conditions and medium requirements necessary for Cyc A production by Fusarium roseum, a microorganism that has not yet been studied for this target.
MATERIALS AND METHODS
Microorganism
Fusarium roseum CZ1 was locally isolated from a soil sample cultivated with clover (Trifolium alexanderinum) and identified to the species level according to Booth (8) and Domsch et al. (13). Potato dextrose agar (PDA) was used for isolation and maintenance of the experimental fungus.
Cultivation Conditions
To select the proper fermentation medium which supports successful Cyc A production, a total of six types of media (Table 2) differing in the composition of their constituents were tested. A fungal agar disc (6 mm, diameter) of 7-days old culture was introduced into a 250 ml flask containing 50 ml of the autoclaved broth medium. The culture flasks were incubated at 30 ºC for 7 days. In the effort to increase Cyc A yields, several experimental conditions were tried. The effect of physical factors including agitation rates of incubation at 120 and 200 rpm, different fermentation periods varied from 2 to 16 days, a range of initial pH values of the medium varied from 2.0 to 9.0, different numbers of inoculated fungal agar discs varied from 1.0 to 5.0 and different volumes of the cultivation medium at 25 to 100 ml in 250 ml capacity flask, were investigated.
In the experimental series, ten different compounds were tried individually as carbon sources. These compounds were arabinose, glucose, galactose, glycerol, mannitol, maltose, lactose, sucrose, cellulose, and starch. All were used at 3% concentration, the same as that of sucrose in the original yeast extract-supplemented Czapek-Dox's medium. Different concentrations of the most productive carbon sources varied from 2% to 6% (w/v) were tested. Eight different nitrogen sources including organic and inorganic ones were added individually to the medium as substitutes for yeast extract. Substitution of yeast extract by nitrogenous sources was done in an equal nitrogen basis. These nitrogen sources were peptone, casein, soybean meal, yeast extract, ammonium chloride, ammonium sulfate, sodium nitrate, and potassium nitrate. Different concentrations of the most productive nitrogen source were also investigated.
Analytical Methods
Determination of Dry Cell Weight: Culture broth (50 ml) was filtered through preweighted Whatman No.1 filter papers. The cells were then dried at 80 ºC to a constant weight prior to measuring dry weight.
Extraction of Cyc A from F. roseum Culture: The methods of extraction and analysis were adopted according to Sawai et al. (29), which can be summarized, as follows: the culture filtrate was extracted three times with equal volumes of ethyl acetate, shaked for about 30 minutes and allowed to stand for 30 minutes. The ethyl acetate layer washed three times with 5 % sodium bicarbonate solution and then twice with water. The solvent layer was taken and dried over anhydrous Na2SO4, then evaporated under vaccum till dryness. The resultant dried crud extract was dissolved in methanol and undergone the necessary chromatographic analysis.
Qualitative and Quantitative Determination of CycA: Cyc A was determined qualitatively and quantitatively according to the method described by Dreyfuss et al. (12) using precoated thin layer chromatographic (TLC) plates with silica gel GF-254. The methanol extract samples and authentic Cyc A (Sandimmun, Novartis Pharma AG, Basle, Switzerland) were spotted on the chromatograms and developed in n-hexane solution then redeveloped in the solvent system of benzene-isopropanol (9:1). Cyc A spots (Rf = 0.37) give violet influorescence under short wave length ultraviolet rays. Detection of spots was more clearly after staining with iodine vapour. The sopts were scraped and then dissolved in a known volume of methanol and the absorption was then measured using an ultraviolet spectrophotometer (Spectronic 21 D-Molton Roy) at 230 nm. Amounts of Cyc A were then estimated from a standard curve. The identity of Cyc A was also confirmed by 1H NMR spectroscopic analysis which was kindly performed in the Microanalytical Unit of the Cairo University, Giza, Egypt. The spectra were recorded in CDCL3 at 25 ºC with a Varian Gemini-200 spectrometer (Varian, UK) at 200 MHz using tetramethylsilane as an internal reference. Chemical shifts are expressed in ppm (δ) units.
All experemints were conducted in triplicates and the mean ± standard deviation (SD) of these triplicates was calculated.
RESULTS
1H NMR Spectra
In the present study, the resultant crystalline Cyc A from F. roseum cultures was subjected to 1H NMR analysis after submerged cultivation of the experimental fungus, followed by extraction of the filtrate with ethyl acetate and chromatographic separation. The chemical shifts are expressed in ppm (δ) units, recorded in Table 1. The 1H NMR spectrum of Cyc A in CDCL3 showed four N-H amide protons as doublets at 7.26 ppm, seven N-methyl signals (two are singlets at 3.47 and 3.37 ppm and five are doublets between 2.98 and 3.29 ppm) and one formed O-H group as doublet at 3.647 ppm. The aliphatic α-protons resonate between 3.60 to 3.70 ppm, the β-protons in the range of 1.12 to 3.68 ppm, while δ- and γ- protons resonate in the range of 0.841 to 3.66 ppm.
Optimization of Cultural Conditions
Suitability of the Cultivation Medium: The results given in Table 2 indicate that Czapek-Dox's broth medium supplemented with 0.5 % yeast extract proved to be the most suitable and conductive medium for the production process where it yeilded 11.45 mg Cyc A/ l. However, the maximum mycelium growth (7.58 g/ l) was recorded on yeast-sucrose broth medium. Therefore, yeast extract supplemented Czapek-Dox's medium was used as a working medium throughout the present study.
Our results indicated that high concentration of the synthesized Cyc A by the experimental organism was produced in the culture filtrate (11.45 mg/ l) and a lower level was produced in the fungal mycelia (1.51 mg/g biomass) after cultivation of the fungus in yeast extract supplemented Czapek-Dox's medium for 7 days at 30 ºC. Hence, the present study was directed to optimize the production of Cyc A from the culture filtrate of the experimental organism.
Effects of Static and Agitation Conditions of Incubation: With regard to the nature of the fungus growth, it was observed that the agitation rate at 200 rpm showed an increase in fungal pellets more than those recorded in case of the agitation rate at 120 rpm. The results showed that the dry cell weights produced either at the agitation rate 120 rpm or 200 rpm were higher than those produced at static conditions (data not shown). The best Cyc A yield (18.15 mg/ l) was obtained at 120 rpm, however no detectable amount of Cyc A was determined at 200 rpm. The static conditions produced approximately 59 % of the concentration of Cyc A produced at 120 rpm.
Time Course of Cyc A Production:F. roseum was cultured in Czapek-Dox's broth medium supplemented with 0.5 % yeast extract and incubated at 30 ºC in a shaking incubator at 120 rpm. The time course profile of Cyc A production is shown in Figure 2. Any increase in the growth period was accompanied with a clear increase in the values of both dry cell weights and Cyc A till maximum values (11.16 g dry cell weight / l and 18.43 mg Cyc A / l) were obtained at 14 and 7 days, respectively; then a decline was obtained in both parameters with increasing of the growth periods. So that, the experimental fungus was grown for 7 days in the subsequent experiments throughout the present work.
Effects of Incubation Temperature: The results given in Figure 3 show that the optimum incubation temperature for maximum values of dry cell growth (8.54 g/ l) and Cyc A (18.94 mg/ l) was 30 ºC, afterwhich a gradual decreases were obtained in both parameters with the increase of incubation temperature to 35 and 40 ºC.
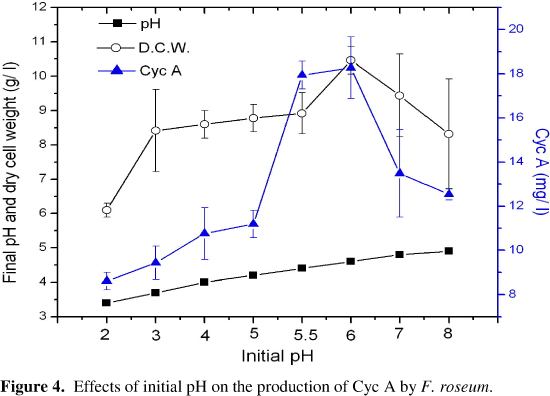
Effects of Initial pH: The initial pH of the medium was adjusted to a range of 2.0 - 9.0 by adding varying amounts of 1 N HCL and 1 N NaOH. In most cases, final pH values were between 3.4 and 4.6. It is obvious that the values of dry cell weights and Cyc A increased with the increase of initial pH values, reaching a maximum for both parameters at pH 6.0 and then decreased at the higher pH values, (Figure 4). Culture medium of pH 6.0 was selected for further investigation throughout this work.
Numbers of Inoculum Culture Discs: Agar discs (6 mm, diameter) of 7 days old F. roseum culture were inoculated in different numbers varied from 1 to 6 into yeast extract supplemented Czapek-Dox's broth. In particular, the results showed that the increases in the numbers of inoculum discs were associated with increases in the productivity of Cyc A (data not shown). The maximum concentration of Cyc A obtained was 21.7 mg/l ~ 1.09 mg/50 ml when the medium was inoculated with 5 fungal agar discs, after which the Cyc A productivity began to decrease when the medium was inoculated with 6 fungal agar discs. The highest dry cell weights (10.46 g/l ~ 0.523 mg/50 ml) were obtained when the medium was inoculated with single fungal agar disc after which, the values were decreased gradually with the increase of numbers of inoculum discs.
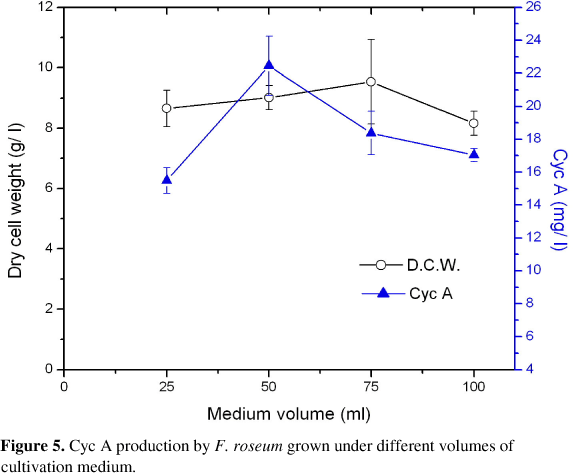
Effects of the Medium Volume: It is evident from Figure 5 that the dry cell weights increased with the increase in culture volume till maximum values (9.54 g/ l) when a 250 ml-flask containing 75 ml fermentation medium was used. The highest Cyc A level (22.46 mg/ l) was obtained by using 250 ml-flask containing production medium equal to 50 ml, afterwhich; the further increase in the size of the fermentation medium to 75 ml or 100 ml /250 ml flask is accompanied with a decrease in the Cyc A concentration.
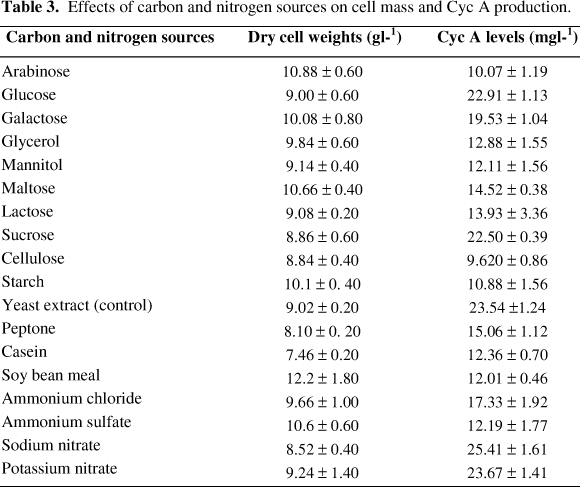
Effects of Carbon Sources: The effects of various carbon sources on the production of Cyc A were investigated (Table 3). Glucose was found to be the most effective substrate among the carbon sources used, and the highest Cyc A concentration obtained was 22.90 mg/ l. Sucrose produced also a satisfactory amounts of Cyc A (22.5 mg/ l) followed by galactose which gave a concentration of Cyc A, approximately twice that obtained with cellulose. Although the experimental fungus produced a low level of Cyc A when arabinose was used as a carbon substrate, but it gave a maximal cell mass (10.88 g/ l) on the same substrate. Maltose was also a good substrate for growth of the fungus and produced 98 % of the dry cell weight produced by arabinose.
Effects of Glucose Concentrations: Different concentrations of glucose (2.0 - 6.0 %, w/v) were added to medium containing 0.5 % (w/v) yeast extract. The maximum concentration of Cyc A (23.38 mg/ l) was recorded at 3.0 % of glucose, which was followed by a sharp decrease (16.26 and 16.22 mg/ l) at higher glucose concentrations (5.0 and 6.0 %, respectively). Dry cell weights increased as glucose concentrations increased till a maximum value (13.44 g/ l) at 5.0 % glucose concentration. Dry cell weights obtained at 6.0 % glucose represented, approximately 95.5 % of that produced at 5.0 %. However, dry cell weights recorded at 2.0 % was approximately, 62.5 % of that produced at 5.0 % (Figure 6).
Effects of Nitrogen Sources: Various organic and inorganic nitrogen sources were added (as equivalent to 5.0 g/ l of yeast extract) separately to the modified medium containing 3.0 % (w/v) glucose. The highest Cyc A level (25.41 mg/ l) was obtained on sodium nitrate compared with that produced on the tested nitrogen sources (Table 3). It is shown that the growth of F. roseum is not associated with Cyc A production because when soybean meal was used as a nitrogen source, the cell mass was maximum (12.2 g /l), however Cyc A production was minimum (12.01 mg/ l). Sodium nitrate as a substrate for growth produced 92 % of the dry cell weights produced by potassium nitrate, while the concentration of Cyc A produced in the presence of potassium nitrate was 93 % of that produced in the presence of sodium nitrate, although both of them are nitrate nitrogen sources. With respect to the effect of organic nitrogenous sources on F. roseum growth, yeast extract produced approximately 74 % of the cell mass produced by soybean meal, while the concentration of Cyc A produced by yeast extract was approximately twice of that obtained with soybean meal. Yeast extract as an organic nitrogenous substrate for growth and Cyc A production by F. roseum was more favorable than peptone and casein. With regard to the effect of ammonium nitrogen sources on growth and Cyc A production by F. roseum, ammonium sulphate gave a higher cell mass than that obtained with ammonium chloride, while the contrary took place in case of Cyc A production because the fungus produced higher concentration of Cyc A in presence of ammonium chloride than that produced in the presence of ammonium sulfate.
Effects of Sodium Nitrate Concentrations: Various concentrations of sodium nitrate (0.05, 0.1, 0.2, 0.3 and 0.4%, w/v) were added to media containing 3.0 % (w/v) glucose. When 0.2% sodium nitrate was used, maximum values of dry cell weight (~ 8.64 g/ l) and Cyc A (~ 27.43 mg/ l) were obtained, afterwhich a decrease was observed in both parameters on increasing the sodium nitrate concentration (Figure 7). These results showed that the optimum carbon/ nitrogen ratio (C/ N) of the broth medium should be equivalent to 15: 1 for achieving the highest production of Cyc A by F. roseum.
DISCUSSION
Results of different experiments to determine suitable and chemically defined broth medium for the production of Cyc A, reported to be immunosuppressive compound that has been widely used to diminish transplant rejection, reveal that a medium of the following composition was suitable for this purpose: glucose, 30; NaNO3, 2; KH2PO4, 1; MgSO4.7H2O, 0.5; KCL, 0.5 (g/ l) and pH adjusted to 6.0. The size of the fermentation medium up to 50 ml/250ml flask inoculated with 5 fungal agar discs (6 mm, diameter) of 7 days old of F. roseum culture and the incubation at 30 ºC at 120 rpm for 7 days were the optimum conditions for the production of Cyc A. Some Fusarium species were reported to produce Cyc A, as F. solani (28, 29), F. oxysporum (28), however no reports have not yet been studied on the production of Cyc A by F. roseum.
The main spectral features are typical for Cyc A: seven N-methyls, four N-H and one O-H were observed in 1H NMR analysis. Similar results were obtained by Ramana Murthy et al. (26). The data in Table 1 agree with that reported by Buchta et al. (9) on [Ala2, val11] cyclosporin, is analogous derivative of Cyc A from Mycelium sterilae.
A number of broth media have been used for production of Cyc A. Isaac et al. (18) used medium consisted of sorbose (2 %), vitamin assay Casamino acids (1 %), KH2PO4 (1 %) and KCL (0.5 %) for maximum production of Cyc A from T. inflatum, however, Balaraman and Mathew (4) used medium containing: glucose (8 %), casein acid hydrolysate (3 %), malt extract (2 %), peptone (1 %) and DL-α amino butyric acid (0.5 %) for maximum production of Cyc A by Tolypocladium species. Formulation of the medium composed of glucose (5 %), bactopeptone (1 %), KH2PO4 (0.5 %), KCL (0.25 %) proved to be suitable for production of Cyc A (2, 28).
Most reports have been made of the production of Cyc A in the culture filtrates (1, 12, 14, 18, 21, 29). However we found two papers reported the extraction of Cyc A from the culture filtrates and mycelia of both Tolypocladium species (4) and Aspergillus terreus (28). Our results showed that high concentration of the synthesized Cyc A by F. roseum was produced in the fungal culture filtrate and a lower level was produced in the fungal mycelia.
In the present study, it was observed that the agitation rate at 200 rpm showed an increase in fungal pellets more than those recorded in case of the agitation rate at 120 rpm. This result agrees with that stated by Sallam et al. (28). The maximum Cyc A yield was obtained at 120 rpm. The same agitation rate was used by Moussaïf et al. (23) for maximum production of cyclosporin by Acremonium luzulae. The production of Cyc A is mainly carried out by submerged fermentation (3, 25). Zocher et al. (38) used 115 rpm for Cyc A biosynthesis from T. inflatum. Kobel and Traber (19) used 180 rpm for the maximum production of Cyc A, while Evers et al. (15) used 200 rpm for maximum cyclosporin production.
Our results showed that Cyc A was produced by F. roseum in maximum yields at pH 6 of the broth medium and after incubation at 30 ºC for 7 days. Abdel-Fattah et al. (1) adjusted the cultivation medium to pH 5.9 for maximum production of Cyc A from T. inflatum after incubation at 27 ºC. Additionally, Kobel and Traber (19), Chun and Agathos (10) and Manuela et al. (22) showed that the best yield of Cyc A was obtained at pH 5.7. Some reports used the incubation temperature at 27 ºC for production of Cyc A (18, 28, 38). The maximum Cyc A production has been reported to vary with respect to the fermentation time course, where Sakamoto et al. (27) found that the best time span for Cyc A production was 10 days, Isaac et al. (18) found that 12 days of growth is a good period for Cyc A production from T. inflatum UAMH 2472. On the other hand, Traber et al. (34) found that 14 days is the best period for maximal production of Cyc A from T. inflatum NRRL 8044. Our data revealed that maximum production of Cyc A was obtained after 7 and 9 days, respectively; then decreased after 9 days and gave the same relative levels at the fermentation period (10-14 days) then decreased after 16 days of incubation.
Cyc A is not generally produced during the vegetative mycelial growth stage, but is generated only in the later pellet production stage (19). Our study showed that Cyc A production is affected by the number of inoculated fungal agar discs and medium size. A 250 ml flask containing production medium equal to 50 ml afforded the highest relative yield of Cyc A. In accordance with this result Abdel-Fattah et al. (1) and Lee et al. (21) used the same volume of the production medium. However, Zocher et al. (38) and Agathos et al. (2) found that 250 ml flask containing 100 ml production medium gave the highest Cyc A yield. The optimum size of the inoculum for the highest Cyc A production was varied in the previous studies, where some reports showed that 3 % of spore inoculum generated the highest level of Cyc A productivity (21). However Chun and Agathos (11) and Abdel-Fattah et al. (1) used 5 % inoculum size for Cyc A production, others reports showed that the best yields of Cyc A were obtained at 10 % inoculum size (2, 19). In our study we found that the maximum concentration of Cyc A was obtained on inoculating 50 ml production medium with 5 fungal agar discs (6 mm, diameter) of 7-days old F. roseum culture.
With regard to the effect of different carbon substrates on Cyc A productivity by F. roseum, glucose (3 %, w/v) was found to be the most favourable source for production of the highest concentration of Cyc A. These results were found to be parallel to those of many reports which showed that glucose as a carbon source (2-3 %) is the most suitable source for the highest production of Cyc A (3, 4, 11, 26, 31), however, Isaac et al. (18) found that sorbose (2 %) followed by glucose (2 %) were the most favorable carbon sources for maximum production of Cyc A by T. niveum UAMH 2472.
Most previous studies showed that peptone (1%, w/v) was the most optimum nitrogen source for production of the best yield of Cyc A from Tolypocladium species (3, 4, 11, 31, 32), however Isaac et al. (18) found that 1 % vitamin assay Casamino acids was the optimum nitrogen source for maximum productivity of Cyc A by T. niveum UAMH 2472, although casitone or peptone gave good yields. In our study we found that the highest Cyc A productivity (ranged from 25.41 to 27.43 mg/ l) was obtained from F. roseum by using sodium nitrate (0.2 %, w/v) and the Cyc A concentration produced in the presence of peptone was approximately, 59.3 % of that produced in the presence of sodium nitrate, as nitrogen sources. A previous study (29) reported that Cyc A concentration of 2.5 mg/ l was produced by F. solani in potato dextrose broth and lower yields were obtained in Richard broth.
In conclusion, the present study investigated the selection of cultivation medium that resulted in enhancing the production of Cyc A by F. roseum, a new Cyc A-producing isolate. In addition, some optimal nutritional and physicochemical conditions for the production of Cyc A were also described. It was observed that the highest productivity of Cyc A was maintained when the fermentation process was carried out using a medium formulation composed of (g/ l): glucose, 30; NaNO3, 2; KH2PO4, 1; MgSO4.7H2O, 0.5; KCL, 0.5, pH 6.0, in a size up to 50ml/250ml capacity flask, inoculated with five agar discs (6 mm, diameter) of 7 days old F. roseum culture, and incubated at 120 rpm for 7 days.
ACKNOWLEDGEMENTS
The authors thank Prof. Dr. Ezzat, S.M. (Botany Department, Faculty of Science, Zagazig University, Egypt) for valuable advice and constructive criticism during progress of this work and Helmy, A.R., assistant lecturer of organic chemistry (Chemistry Department, Faculty of Science, Zagazig University, Egypt) for his critical comments on the results of 1H NMR analysis.
Submitted: June 30, 2009; Returned to authors for corrections: March 10, 2010; Approved: June 21, 2010.
- 1. Abdel-Fattah, Y.R.; El Enshasy, H.; Anwar, M.; Omar, H.; Aboelmagd, E. (2007). Application of factorial experimental designs for optimization of cyclosporine A production by Tolypocladium inflatum in submerged culture. J. Microbiol. Biotechnol. 17 (12), 1930- 1936.
- 2. Agathos, S.N.; Marshal, J. W.; Marati, C.; Parekh, R.; Modhosingh, C. (1986). Physiological and genetic factors for process development of cyclosporin fermentation. J. Ind. Microbiol. 1, 39- 48.
- 3. Balakrishnan, K.; Pandey, A. (1996). Influence of amino acids on the biosynthesis of cyclosporin A by Tolypocladium inflatum Appl. Microbiol. Biotechnol. 45, 800- 803.
- 4. Balaraman, K.; Mathew, N. (2006). Optimization of media composition for the production of cyclosporin A by Tolypocladium species. Indian J. Med. Res. 123, 525- 530.
- 5. Billich, A.; Zocher, R. (1987). Enzymatic synthesis of cyclosporin A. J. Biol. Chem. 262, 17258- 17259.
- 6. Beauchesen, P.R.; Chung, N.S.; Wasan, K.M. (2007). Cyclosporine A: a review of current oral and intravenous delivery systems. Drug Dev. Ind. Pharm. 33, 211- 220.
- 7. Borel, J. F. (1986). Cyclosporin and its future. Prog. Allergy. 38, 9-18.
- 8. Booth, C. (1971): The genus Fusarium Commonwealth, Mycological Institute, Kew Surrey, England.
- 9. Buchta, M.; Jegorov, A.; Cvak, L.; Havlíèek, V.; Budìínský§; Sedmeara, P. (1998). A cyclosporin from Mycelium sterilae Phytochem. 48 (7), 1195-1198.
- 10. Chun, G.-T.; Agathos, S.N. (1989). Immobilization of T. inflatum spores into porous celite beads for cyclosporin A production. J. Biotechnol. 9, 237- 254.
- 11. Chun, G.-T.; Agathos, S.N. (1991). Comparative studies of physiological and environmental effects on the production of cyclosporin A in suspended and immbolized cells of Tolypocladium inflatum, Biotechno. Bioeng. 37, 256- 265.
- 12. Dreyfuss, M.; Härri, E.; Hofmann, H.; Kobel, H.; Pache, W.; Tscherter, H. (1976). Cyclosporin A and C. New metabolites from Trichoderma polysporum (Link ex Pers.) Rifai. Eur. J. Appl. Microbiol. 3, 125-133.
- 13. Domsch, K.H.; Gams, W.; Anderson T.- H. (1980). Compendium of Soil Fungi, Vol.1, Academic Press, A subsidiary of Horcourt, Brace, Jovanovich Publishers, London.
- 14. El-Enshasy, H.; Abdel-Fattah,Y.; Atta, A.; Anwar, M.; Omar, H., Abou El Magd, S.; Abou Zahra, R. (2008). Kinetics of cell growth and cyclosporin A production by Tolypocladium inflatum when scaling up from shake flask to bioreactor. J.Microbiol. Biotechnol. 18 (1), 128-134.
- 15. Evers, M.; Migrani, S.; Carri, J.; Christophe, E. (1999). Preparation of novel cyclosporins derivatives and pharmaceutical composition. Ind. Appl. Microbiol. 49, 193.
- 16. Gams, W. (1997). Tolypocladium Eine Hyphomycetengattung mit geschwollenen Phialiden. Persoonia. 6, 185- 191.
- 17. Heitman, J.; Movva, N.R.; Hall, M.N. (1992). Proline isomerase at the crossroads of protein folding, signal transduction and immunosuppression. New Biol. 4, 448- 460.
- 18. Issac, C.E.; Jonse, A.; Pickard, M.A. (1990). Production of cyclosporins by Tolypocladium niveum strains. Antimicrob. Agents Chemother. 34 (1), 121- 127.
- 19. Kobel, H.; Traber, R. (1982). Directed biosynthesis of cyclosporins. Eur. J. Appl. Microbiol. Biotechnol. 14, 237- 240.
- 20. Lawen, A.; Zocher, R. (1990). Cyclosporin synthetase. The most complex peptide synthesizing multienzyme polypeptide so far described. J. Biol. Chem. 256, 11355- 11360.
- 21. Lee, M.-J.; Lee, H.N.; Han, K.; Kim, E.S. (2008). Spore inoculum optimization to maximize cyclosporin A production in Tolypocladium niveum J. Microbiol. Biotechnol. 18 (5), 913- 917.
- 22. Manuela, O.; Gerhard, S.; Klaus, T.; Zhaung, S.; Hanspecter, M.; Rene, T.; Elisabeth, S. (1996). Biosynthesis of the unusual aminoacid (4R)-4[(E)-2-butenyl]-4-methyl-L-theronin of cyclosporin A. Enzymatic analysis of the reaction sequence including identification of the methylation precursor in a polyketide pathway. Biochem. 35, 8401- 8412.
- 23. Moussaïf, M.; Jacques, P.; Schaarwächter, P.; Budzikiewicz, H.; Thonart, P. (1997). Cyclosporine C is the main antifungal compound produced by Acremonium luzulae Appl. Environ. Microbiol. 63 (5), 1739- 1743.
- 24. Nakajima, H.; Hamasaki, T.; Nishimura, K.; Kimura, Y.; Udagawa, S.; Sato, S. (1988). Isolation of 2-acetylamino-3-hydroxy-4-methyl-oct-6-enoic-acid,a derivative of the "C9 amino acid" residue of cyclosporins, produced by the fungus Neocosmospora varinfecta E.F. Smith. Agric. Biol. Chem. 52, 1621- 1623.
- 25. Pitchard, D.I. (2005). Sourcing a chemical succession for cyclosporin from parasites and human pathogens. Drug Discovery Today. 10, 688-691.
- 26. Ramana Murthy, M.V.; Mohan, E.V.S.; SadhuKhan, A.K. (1999). Cyclosporin A production by Tolypocladium inflatum using solid state fermentation. Process Biochem. 34, 269- 280.
- 27. Sakamoto, K.; Tsujii, E.; Miyauchi, M.; Nakanishi, T.; Yamashita, M.; Shigematsu, N.; Tada, T.; Lzumi, S.; Okuhara, M. (1993). FR 901459, a novel immunosuppressant isolated from Stachybotrys chartarum No 19392. J. Antibiot. 46, 1788- 1798.
- 28. Sallam, L.A.R.; El-Refai, A.H.; Hamdy, A.A.; El-Minofi, H.A.; Abdel-Salam, I.S. (2003). Role of some fermentation parameters on cyclosporine A production by a new isolate of Aspergillus terreus J. Gen. Appl. Microbiol. 49, 321- 328.
- 29. Sawai, K.; Okuno, T.; Tereda, Y.; Harada,Y.; Wawamura, K.; Sasaki, H.; Takao, S. (1981). Isolation and properties of two antifungal substances from Fusarium solani Agric. Biol. Chem. 45, 1223- 1228.
- 30. Schmidt, B.; Reisner, D.; Lawen, A.; Kleinkauf, H. (1992). Cyclosporin synthetase is a 1.4 M Da multienzyme polypeptide. Re-evaluation of the molecular mass of various peptide synthetases. FEBS Lett. 307, 355- 360.
- 31. Sekar, C.; Rajasekar, V.W.; Balaraman, K. (1997). Production of cyclosporin by solid state fermentation Bioprocess. Eng. 17, 257- 259.
- 32. Sekar, C.; Balaraman, K. (1998): Optimization studies on the production of cyclosporin A by solid state fermentation. Bioprocess Eng. 18, 293- 296
- 33. Sumegi,V.; Haszon, I.; Bereczki, C.; Papp, F.; Turi, S. (2008). Long-term follow-up after cyclophosphamide and cyclosporine A therapy in steroid-dependant and resistant nefrotic syndrome. Pediatr. Nephrol. 23, 1085- 1092.
- 34. Traber, R.; Hofmann, H.; Kobel, H. (1989). Cyclosporins, new analogous by precursor directed biosynthesis. J. Antibiot. 40: 591- 597.
- 35. Von Arx, J.A. (1986). Tolypocladium, a synonym of Beauveria Mycotaxon. 25, 153- 158.
- 36. Zhou, X.; Hu, Y.; Tian, Y.; Hu, X. (2009). Effect of N-trimethyl chitosan enhancing the dissolution properties of the lipophilic drugs cyclosporin A. Carbohydrate Polymers. 76, 285- 290.
- 37. Zocher, R.; Madry, N.; Peeter, H.; Klienkauf, H. (1984). Biosynthesis of cyclosporin A. Phytochem. 23, 549- 551.
- 38. Zocher, R.;Nihira, T.; Paul, E.; Madry, N.; Peeters, H.; Kleinkauf, H.; Keller, U. (1986). Biosynthesis of cyclosporin A: Partial purification and properties of a multifunctional enzyme from Tolypocladium inflatum Biochem. 25; 550- 555.
Publication Dates
-
Publication in this collection
09 Aug 2010 -
Date of issue
Dec 2010
History
-
Accepted
21 June 2010 -
Reviewed
10 Mar 2010 -
Received
30 June 2009