Abstract
The influences of fish infusion decarboxylase broth (IDB) on biogenic amines (BA) formation by lactic acid bacteria (LAB) were investigated. BA productions by single LAB strains were tested in five different fish (anchovy, mackerel, white shark, sardine and gilthead seabream) IDB. The result of the study showed that significant differences in ammonia (AMN) and BA production were observed among the LAB strains in fish IDB (p < 0.05). The highest AMN and TMA production by LAB strains were observed for white shark IDB. The all tested bacteria had decarboxylation activity in fish IDB. The uppermost accumulated amines by LAB strains were tyramine (TYM), dopamine, serotonin and spermidine. The maximum histamine production was observed in sardine (101.69 mg/L) and mackerel (100.84 mg/L) IDB by Leuconostoc mesenteroides subsp. cremoris and Pediococcus acidophilus, respectively. Lactobacillus delbrueckii subsp. lactis and Pediococcus acidophilus had a high TYM producing capability (2943 mg/L and 1157 mg/L) in sardine IDB.
biogenic amines; lactic acid bacteria; starter cultures; fish infusion broth
FOOD
RESEARCH PAPER
The influences of fish infusion broth on the biogenic amines formation by lactic acid bacteria
Esmeray KüleyI; Fatih ÖzogulI; Esra BalikçiI; Mustafa DurmusI; Deniz AyasII
IDepartment of Seafood Processing Technology, Faculty of Fisheries, Cukurova University, Adana, Turkey
IIDepartment of Seafood Processing Technology, Faculty of Fisheries, Mersin University, Mersin, Turkey
Send correspondence toSend correspondence to: E. Kuley Department of Seafood Processing Technology, Faculty of Fisheries, Cukurova University 01330 Balcaly, Adana, Turkey E-mail: eboga@cu.edu.tr
ABSTRACT
The influences of fish infusion decarboxylase broth (IDB) on biogenic amines (BA) formation by lactic acid bacteria (LAB) were investigated. BA productions by single LAB strains were tested in five different fish (anchovy, mackerel, white shark, sardine and gilthead seabream) IDB. The result of the study showed that significant differences in ammonia (AMN) and BA production were observed among the LAB strains in fish IDB (p < 0.05). The highest AMN and TMA production by LAB strains were observed for white shark IDB. The all tested bacteria had decarboxylation activity in fish IDB. The uppermost accumulated amines by LAB strains were tyramine (TYM), dopamine, serotonin and spermidine. The maximum histamine production was observed in sardine (101.69 mg/L) and mackerel (100.84 mg/L) IDB by Leuconostoc mesenteroides subsp. cremoris and Pediococcus acidophilus, respectively. Lactobacillus delbrueckii subsp. lactis and Pediococcus acidophilus had a high TYM producing capability (2943 mg/L and 1157 mg/L) in sardine IDB.
Key words: biogenic amines, lactic acid bacteria, starter cultures, fish infusion broth.
Introduction
Biogenic amines (BA) production in seafood are one of the considerable public concern since histamine and possibly other biogenic amines such as cadaverine and putrescine are responsible for histamine fish poisoning (Taylor 1986; Jorgensen et al., 2000) and serve as chemical indicators of fish spoilage (Alberto et al., 2002). The toxicity of histamine appears to be enhanced by cadaverine (CAD) and putrescine (PUT) since they inhibit the histaminedetoxifying enzymes, which are diamine oxidase and histamine N-methyltransferase (Stratton et al., 1991; Yongsawatdigul et al., 2004). Biogenic amines such as PUT, CAD, spermidine (SPD), spermine (SPN), histamine (HIM), tyramine (TYM) and tryptamine (TRP) are widely distributed in proteinaceous foods (Krizek et al., 2004). Most studies have been focused on BA formation in fish and meat products (Ruiz-Cappillas and Jimenez-Colmenero, 2004) due to their proteinaceous nature and propensity to form BA from free amino acids (Magwamba 2010).
Accumulation of BA in foods requires (I) the availability of precursors (i.e. amino acids), (II) the presence of microorganisms with amino acid decarboxylases enzyme, which either derived from environmental contamination or from an added starter culture and (III) favourable conditions that allow bacterial growth, decarboxylase synthesis and decarboxylase activity (Bodmer et al., 1999; Karovicova and Kohajdova, 2005; Stadnik and Dolatowski, 2010). Although amino acid decarboxylases are not widely distributed among bacteria, species of many genera such as Bacillus, Citrobacter, Clostridium, Klebsiella, Escherichia, Proteus, Pseudomonas, Shigella, Photobacterium and the lactic acid bacteria (Lactobacillus, Pediococcus, and Streptococcus) are capable of decarboxylating one or more amino acids (Halasz et al., 1994; Silla-Santos 1996; Karovicova and Kohajdova, 2005; Özogul and Özogul, 2005).
BA production by bio-preservative features of lactic acid bacteria (LAB) have been reported (Connil et al., 2002). Some strains of LAB synthesize histamine because of their ability to possess the histidine decarboxylase gene (Landete et al., 2005; Lucas et al., 2005). The presence of BA in fermented foods is due to the decarboxylase activity of the LAB used as starter culture, and the action of some spoilage bacteria (Marcobal et al., 2006). High BA contents have been reported in some fish products such as fish sauce, fish paste, fish salads, cold-smoked fish (Leuschner and Hammes, 1999; Petaja et al., 2000; Yongsawatdigul et al., 2004; Jorgensen et al., 2000; Gonzalez-Rodriguez et al., 2002; Thapa et al., 2006; Udomsil et al., 2010; Zaman et al., 2010; Zhong-Yi et al., 2010). The production of BA by LAB to be selected as starter cultures is not a desirable feature for food industry (Buchenhuskes 1993). Thus some safety aspects of the LAB isolates of food products must be investigated before considering their use for the bio-preservation (Matamoros et al., 2009). Before the use of the LAB in the fish product, the quantitative analysis of BA is very essential to confirm the bacterial production. A great amount of research has been focused on BA production in fish and fish products. However no study has been found on BA production by single LAB strains which is used in fish infusion broth. Therefore, the aim of the study was to investigate the function of some commercially important LAB strains on biogenic amine production in different fish infusion decarboxylase broth (IDB).
Materials and Methods
Bacterial strains
The used LAB species were Lactococcus lactis subsp. cremoris (MG 1363), Lactococcus lactis subsp. lactis (IL 1403), Lactobacillus plantarum (FI8595) and Streptococcus thermophilus (NCFB2392). They were obtained from Sutcu Imam University, Kahramanmaras, Turkey in BGML stock culture. Leuconostoc mesenteroides subsp. cremoris (DSMZ 20346), Lactobacillus acidophilus (ATCC 11975), Pediococcus acidophilus (ATCC 25741) and Lactobacillus delbrueckii subsp. lactis (ATCC 10697) were purchased from Institute of Refik Saydam Hifzisihha (Ankara, Turkey).
Fish species
In the present study, fish decarboxylase infusion broth was prepared using five different fish species which were gilthead seabream (Sparus aurata), sardine (Sardinella aurita) anchovy (Engraulis encrasicolus), white shark (Carcharodon carchairas), and mackerel (Scomber scombrus).
Biogenic amine analysis
Culture media and bacterial extraction
Fish infusion broth was prepared according to method of Okuzumi et al. (1982) with minor modifications. Two hundred fifty grams of fish flesh was homogenised with 2 volumes of water (w/v), steamed at 100 ºC for 1 hour and filtered. The filtrate was enriched with 1% glucose and 0.5% NaCl. In order to decarboxylate amino acid by bacteria, 3 mg pyridoxal HCl addition was made in each infusion broth before autoclaving process.
MRS and M17 broth were used for propagation of LAB cultures. Lactic acid bacterial strains were incubated at 37 ºC for 24 hour which after 0.5 mL of these bacterial cultures was removed and put into the fish IDB to decarboxylate amino acid.
For extraction of LAB cultures, 5 mL of the fish IDB containing LAB strains were removed to separate bottles and then 2 mL trichloroacetic acid was added. They were centrifuged at 3000xg for 10 min and then filtered through a Whatman filter paper (Whatman GmbH, Dassel, Germany). After that, 4 mL of bacterial supernatant was taken for derivatisation from each of LAB bacterial strains.
Chemical reagents
All BA standards were purchased from Sigma-Aldrich (Munich, Germany). The mobile phase consisted of acetonitrile and HPLC grade water for amine analyses.
Preparation of standard amine solution
HIM dihydrochloride (165.7 mg), TYM hydrochloride (126.7 mg), TRP hydrochloride (122.8 mg), PUT dihydrochloride (182.9 mg), 2-phenylethylamine (PHEN) hydrochloride (130.1 mg), CAD dihydrochloride (171.4 mg), SPD trihydrochloride (175.3 mg), SPN tetrahydrochloride (172.0 mg), 5-hydroxytryptamine (Serotonin, SER) (133.9 mg), 3-hydroxytyramine hydrochloride (Dopamine, DOP) (123.8 mg), agmatine (AGM) sulphate (175.4 mg), trimethylamine (TMA) hydrochloride (161.7 mg) and ammonium chloride (296.9 mg) were dissolved in 10 mL HPLC grade water. The final concentration of free base for each amine was 10 mg mL-1 solution.
Derivatisation of extract from bacterial broth culture
A stock solution was prepared by dissolving 2% benzoyl chloride in acetonitrile to enhance the reaction with amines. For derivatisation of standard amine solutions, 100 !L was taken (4 mL for extracted bacterial cultures) from each free base standard solution (10 mg mL-1).1mL of sodium hydroxide (2 M) was added, followed by 1 mL of 2% benzoyl chloride (dissolved in acetonitrile) and the solution mixed on a vortex mixer for 1 min. The reaction mixture was left at room temperature for 5 min and then centrifuged for 10 min. After that, the benzoylation was stopped by adding 2 mL of saturated sodium chloride solution and the solution extracted twice with 2 mL of diethyl ether. The upper organic layer was transferred into a clean tube after mixing. Afterwards, the organic layer was evaporated to dryness in a stream of nitrogen. The residue was dissolved in 1 mL of acetonitrile and 10 !L aliquots were injected into the HPLC.
Analytical method
BA analysis was done using the method of Özogul (2004) and measured in mg amines per litre broth. The confirmation of BA production was accomplished using a rapid HPLC method with a reversed phase column by using a gradient elution program. Same analytic method was used for ammonia and trimethylamine (TMA) separation.
HPLC apparatus and column
A Shimadzu Prominence HPLC apparatus (Shimadzu, Kyoto, Japan) equipped with a SPD-M20A diode array detector and two binary gradient pumps (Shimadzu LC-10AT), auto sampler (SIL 20AC), column oven (CTO-20AC), and a communication bus module (CBM-20A) with valve unit FCV-11AL was used. The column was a reverse-phase, ODS Hypersil 5, 250x4.6 mm (Phenomenex, Macclesfield, Cheshire, UK) for the BA analyses.
Statistical analysis
The mean value and standard deviation were calculated from the data obtained from the four samples for each treatment. One way ANOVA was used to determine the significance of differences at p < 0.05. All statistics were performed using SPSS 15.0 for Windows (SPSS Inc., Chicago, IL. USA).
Results
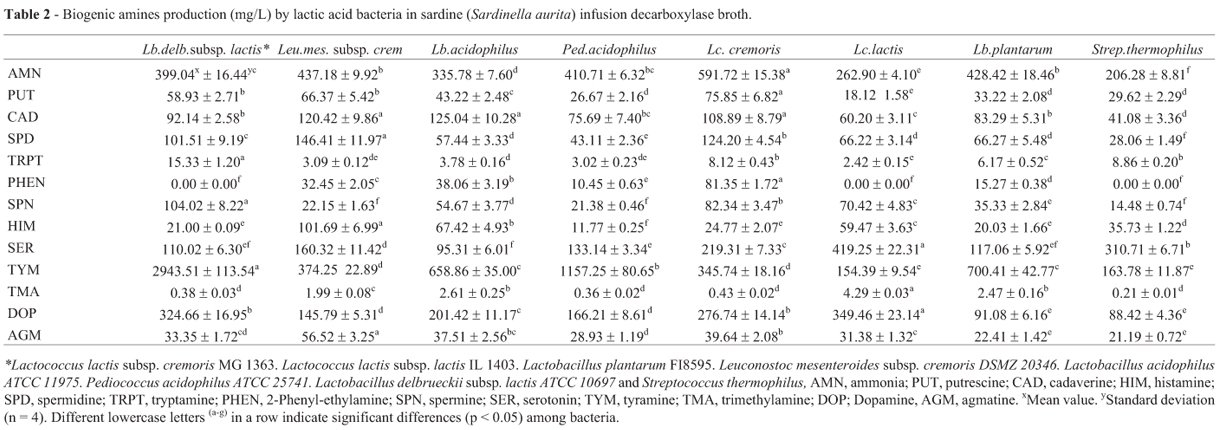
Ammonia (AMN) and BA production by LAB strains in five different fish IDB were given in Tables 1 to 5. Significant differences in AMN were found among the LAB strains and fish IDB (p < 0.05). The all used bacteria produced more than 200 mg/L of AMN. Lc. lactis subsp. cremoris produced the highest amount of AMN (591.72 mg/L) in sardine IDB,(p < 0.05). The lowest AMN production was observed by Strep. thermophilus in sardine infusion broth (Table 2), whereas LAB strains had the highest ability to produce AMN in white shark infusion broth (Table 5).
There were also significant differences in BA production among the LAB strains (p < 0.05) for all fish IDB. The strains produced all amine in fish IDB (Tables 1 to 5) apart from spermine (SPN) and 2-phenylethylamine (PHEN). Putrescine (PUT) production by LAB strains was ranged from 1.85 (for Strep. thermophilus in white shark IDB) to 139.98 mg/L (for Ped. acidophilus in anchovy IDB).
Significant differences in histamine (HIM) production was observed among the LAB strains (p < 0.05). Lc. lactis subsp. cremoris, Leuc. mesenteroides subsp. cremoris and Lc. produced cadaverine (CAD) more than 100 mg/L in sardine IDB, whereas CAD production by LAB strains in the other mediums was less than 93 mg/L. In mackerel IDB, the lowest spermidine (SPD) production was found for Lactococcus lactis subsp. cremoris (11.54 mg/L). In anchovy IDB, SPD production by Lactobacillus spp. was 55 mg/L. Lb. delbrueckii subsp. lactis, Lc. lactis subsp. lactis and Strep. thermophilus in sardine IDB as well as Lc. lactis subsp. lactis in mackerel IDB had not an ability to produce PHEN. However, Lc. lactis subsp. cremoris was found as good PHEN producer in gilthead seabream IDB (209.52 mg/L). Lactococcus spp. had also good activity to produce PHEN in anchovy IDB (Table 1). In white shark IDB, significant PHEN production was observed for Lb. delbrueckii subsp. lactis and Leu. mesenteroides subsp. cremoris.
Most of LAB strains produced below 9 mg/L of TMA. However, Lb. plantarum and Strep. thermophilus accumulated significant amount of TMA (784.46 and 786.75 mg/L, respectively) in white shark IDB (p < 0.05) (Table 5). LAB strains i.e. Lb. delbrueckii subsp. lactis, Lc. lactis subsp. lactis and Lc. lactis subsp. cremoris also had a high TMA formation in anchovy IDB (Table 1). Lc. lactis subsp. lactis. was one of the LAB strains produced good amount of TMA in mackerel IDB. Tryptamine (TRP) was one of the lowest produced amine by LAB strains. Agmatine (AGM) production by LAB strains was between 14 and 101 mg/L.
Discussion
Lc. lactis subsp. lactis (4991.38 mg/L) and Ped. acidophilus (4348.38 mg/L) were characterized as the highest AMN producer in shark IDB among the tested LAB strains. In anchovy IDB, Ped. acidophilus produced the highest amount of AMN (670.95 mg/L), whereas Lc. lactis subsp. cremoris accumulated the lowest level of that (281.96 mg/L). Lc. lactis subsp. cremoris and Lb. plantarum in gilthead seabream IDB were the highest ability in AMN production (p < 0.05). In mackerel IDB, AMN production was ranged from 298 mg/L (Lb. delbrueckii subsp. lactis) to 507 mg/L (Lc. lactis subsp. lactis).
The highest accumulated amines by LAB strains were generally TYM, DOP, SER and SPD, although other amines produced at significant levels (p < 0.05). Bunkova et al. (2009) reported that Lc. lactis subsp. cremoris, Strep. thermophilus and Lb. delbrueckii subsp. bulgaricus produced TYM but did not produce other tested amines. Herring and tuna fish salads inoculated with Lactobacillus curvatus LTH 975 had PUT, CAD, HIM, TYM accumulation (Leuschner and Hammes, 1999). However, in coldsmoked fish fermented with LAB, the LAB used was unable to produce CAD, HIM or TYM (Leuschner and Hammes, 1999). Similar results were found by Thapa et al. (2006) who reported none of the strains of LAB including Lc. lactis subsp. cremoris, Lc. lactis subsp. lactis, Lb. plantarum, Leu. mesenteroides and Pediococcus pentosaceus isolated from traditionally processed fish products produced BA.
Özogul and Özogul (2005) defined four categories in order to simplify the discussion of amine production by bacterial strains, which are: prolific amine former (> 1000 mg/L), good amine former (100-1000 mg/L), medium amine former (10-100 mg/L) and poor amine former (< 10 mg/L). According to this category, LAB strains seemed to poor PUT producer in white shark and mackerel IDB (Tables 4 and 5). However, Leuc. mesenteroides subsp. cremoris in white shark IDB, and Lc. lactis subsp. cremoris in sardine and gilthead seabream IDB accumulated high amount of PUT (> 75 mg/L). In mackerel IDB, PUT production was above the 5.7 mg/L and the highest production was found for Lb. plantarum (30.71 mg/L). Lb. plantarum N4 isolated from wine was able to produce putrescine from ornithine (Arena and Manca de Nadra, 2001). PUT was reported as main amine produced by Leu. mesenteroides and Lactobacillus zeae (Morrena-Arribas and Polo, 2008).
CAD was one of the most abundant amines in fish sauce with maximum reported value of 1429 ppm (Zaman et al., 2010). In the present study, Lc. lactis subsp. cremoris, Leuc. mesenteroides subsp. cremoris and Lc. acidophilus were good CAD producer in sardine IDB (> 100 mg/L) (Table 2). Similar production was also observed for Ped. acidophilus in anchovy IDB (Table 1). In the other mediums, CAD production by LAB strains was less than 93 mg/L. In gilthead seabream IDB, the highest CAD production was observed for Lc. lactis subsp. cremoris. In the recently study, Özogul (2011) reported that lower CAD production by Lc. lactis subsp. cremoris and Strep. thermophilus in histidine decarboxylase broth was found (< 17 mg/L). In the current study, the lowest CAD production by LAB strains was generally observed for white shark and mackerel IDB. There were also not significant differences between white shark and mackerel IDB in terms of CAD production by LAB strains apart from Ped. acidophilus and Lc. acidophilus.
SPD was one of the most accumulated amine by LAB strains. Leuc. mesenteroides subsp. cremoris, Ped. acidophilus and Strep. thermophilus produced more than 100 mg/L of SPD in sardine, mackerel and anchovy IDB, respectively. However, SPD production by Lc. acidophilus was 6-fold higher in white shark, compared with those bacteria. However, SPD production by LAB strains did not differ statistically except for Lb. acidophilus in white shark IDB (p > 0.05). The LAB strains produced medium amount of SPD in gilthead seabream IDB. Zaman et al. (2010) reported that SPD, TRP, PHEN and SPN were minor amines in fish sauce. Similarly, TRP was one of the lowest produced amine by LAB strains. The lowest TRP production was observed from Lc. lactis subsp. cremoris (mackerel IDB) and Strep. thermophilus (white shark IDB), whereas Leuc. mesenteroides subsp. cremoris and Lb. plantarum accumulated medium amount of TRP in white shark IDB.
Özogul (2011) reported that none of the LAB strains produced SPN in histidine decarboxylase broth. Similar results were obtained from this study associated with SPN production by Lc. lactis subsp. cremoris, Lc. lactis subsp. lactis, Leu. mesenteroides subsp. cremoris in white shark IDB. Lb. plantarum was one of the highest amounts of SPN producer. Similarly, SPN production by Lc. lactis subsp. cremoris in anchovy IDB was negligible level, whereas Strep. thermophilus produced high amount of SPN (116.71 mg/L). No significant differences in SPN were observed between Lc. lactis and Lb. plantarum in anchovy IDB (p > 0.05). There were also not significant differences (p > 0.05) in SPN production among the Ped. acidophilus, Lc. lactis subsp. lactis, Lb. plantarum and Strep. thermophilus in gilthead seabream IDB (~64 mg/L). In mackerel IDB, Lc. lactis subsp. cremoris produced low amount of SPN (8.62 mg/L), while SPN production by Lb. acidophilus was 51.30 mg/L (p < 0.05).
HIM production varies depending on bacterial strains and IDB. Matamoros et al. (2009) reported that HIM and tyramine (TYM) production for Leuconostoc gelidum, Lactococcus piscium, Lactobacillus fuchuensis and Carnobacterium alterfunditum isolated from seafood products were below the 5 mg/L. HIM was the only major BA produced by various strains of lactic acid bacteria from fish sauce (Udomsil et al., 2010). In the recent study, it was shown that HIM production in histidine decarboxylase broth was negligible by Lc. lactis subsp. cremoris, whereas Lc. lactis subsp. lactis, Strep. thermophilus and Lb. plantarum did not produce HIM (Özogul, 2011). However, in the present study, the all strains had an ability to produce HIM in fish IDB, ranging from 2 to 102 mg/L (Tables 1-5) indicating that fish muscle was more favourable than specific medium on HIM production. The lowest HIM accumulation was found for Lc. lactis subsp. lactis (2.69 mg/L) and Ped. acidophilus (2.69 mg/L) in white shark and anchovy IDB, respectively. In anchovy IDB, the highest HIM production was found for Lb. delbrueckii subsp. lactis (47.10 mg/L), whereas Leu. mesenteroides subsp. cremoris, Lb. plantarum and Strep. thermophilus produced similar amount of HIM (p > 0.05). Lactobacillus buchneri LB14 and Lactobacillus buchneri ST2A produced 344 and 401 mg/L HIM in decarboxylase medium contained histidine, lysine, ornitine and tyrosine (Choudhury et al., 1990). Although most of LAB strains seemed to be medium HIM producer in fish IDB, Leu. mesenteroides subsp. cremoris and Ped. acidophilus was good HIM producer (~100 mg/L) in sardine and mackerel IDB, respectively.
No significant differences in HIM production was observed among the Lb. delbrueckii subsp. lactis, Lc. lactis subsp. cremoris and Lc. lactis subsp. lactis (~ 50 mg/L) in mackerel IDB. In gilthead seabream IDB, the lowest HIM production was found for Lb. delbrueckii subsp. lactis (5.62 mg/L) though there were no significant differences in HIM production between Lc. lactis subsp. cremoris (73.70 mg/L) and Lc. lactis subsp. lactis (74.37 mg/L) (p > 0.05). Tuna and herring salad inoculated with Lb. buchneri revealed 900 ppm and 670 ppm histamine, respectively (Leuschner and Hammes, 1999).
Fadda et al. (2001) found that Lb. plantarum, Lb. casei and Pediococcus acidilactici isolated from fermented sausages did not produce TYM in a medium supplemented with tyrosine at 20 mg/L. Udomsil et al. (2010) reported trace amounts of TYM production (1-5 mg/100 mL) by various LAB strains from fish sauce. In the current study, TYM was one of the main amines produced by LAB strains. Among the fish IDB, most of LAB strains showed high activity in producing TYM in sardine IDB (> 300 mg/L) (Table 2). The reported upper TYM limit of 100-800 mg kg-1 to be toxic doses in foods (Brink et al., 1990; Kim et al., 2009), which were generally exceed by most of LAB strains. Lb. delbrueckii subsp. lactis (2943.51 mg/L) and Ped. acidophilus (1157.25 mg/L) had the highest ability to produce TYM in sardine IDB. In white shark, most of the LAB strains produced less than 55 mg/L. Marino et al. (2008) reported that Strep. thermophilus was a TYM-producer strain. In the present study, Strep. thermophilus (241.57 mg/L), Ped. acidophilus (224.42 mg/L), Lb. plantarum (170.30 mg/L) and Lc. lactis subsp. cremoris (153.64 mg/L) produced the highest amount of TYM in anchovy IDB. There were not significant differences in TYM production between Leu. mesenteroides subsp. cremoris and Lb. acidophilus (~90 mg/L), and also for Lc. lactis subsp. lactis, Lb. plantarum and Strep. thermophilus (~180 mg/L) in gilthead seabream IDB.
The LAB strains were generally good SER producer. Among the fish IDB, the highest SER production was found by Lc. lactis subsp. lactis in mackerel IDB and Lb. delbrueckii subsp. lactis in anchovy IDB, whereas Lb. delbrueckii subsp. lactis was produced the lowest amount of SER in gilthead seabream IDB (p < 0.05). Leu. mesenteroides subsp. cremoris was the main LAB produced highest amount of SER in gilthead seabream IDB. LAB strains such as Lc. lactis subsp. lactis, Lc. lactis subsp. cremoris, Lb. plantarum and Strep. thermophilus showed lower activity of SER and DOP (1 and 5.5 mg/L, respectively) in histidine decarboxylase broth (Özogul, 2011). There were significant differences in DOP production among the LAB strains in fish IDB (p < 0.05). LAB strains produced significant amount of DOP, especially for sardine and white shark IDB. Although Lb. delbrueckii subsp. lactis was poor DOP producer in anchovy IDB, the highest DOP production by Lc. lactis subsp. cremoris was found in this medium (p < 0.05). In mackerel IDB, DOP production was ranged from 32.96 mg/L for Lc. lactis subsp. lactis to 224.74 mg/L for Ped. acidophilus.
AGM is formed from arginine by the enzyme of arginine decarboxylase secreted from lactic acid and nitric acid-reducing bacteria during the fermentation process (Umezu et al., 1977). The lowest AGM production by LAB strains was in white shark IDB (< 42 mg/L) while the highest AGM accumulation was found for Strep. thermophilus (100.32 mg/L) and Ped. acidophilus (75.12 mg/L) in anchovy and mackerel IDB (p < 0.05), respectively. In sardine IDB, AGM production by LAB strains was below the 57 mg/L.
Conclusion
The study results showed that the tested LAB strains had an ability to produce high amount of BA in fish IDB, which were mainly TYM, HIM, DOP, SER and SPD production. Fish species also appeared to play an important role in BA production by LAB strains. Therefore both criteria have to be taken into consideration in order to prevent the bacterial BA production and food poisoning related to fish consumption. Consequently, the ability of LAB strains to produce biogenic amines in fish products can be considered as criteria for the selection of strains and fish species.
Submitted: January 23, 2012
Approved: July 23, 2012
All the content of the journal, except where otherwise noted, is licensed under a Creative Commons License CC BY-NC.
- Alberto MR, Arena ME, Manca de Nadra MC (2002) A comparative survey of two analytical methods for identification and quantification of biogenic amines. Food Control 13:125-129.
- Arena ME, Manca de Nadra MC (2001) Biogenic amine production by Lactobacillus J Appl Microbiol 90:158-162.
- Bodmer S, Imark C, Kneubühl M (1999) Biogenic amines in foods: Histamine and food processing. Inflamm Res 48:296-300.
- Brink BT, Damink C, Joosten HM, Huis In't Veld JH (1990) Occurence and formation of biologically active amines in foods. Int J Food Microbiol 11:73-84.
- Buchenhuskes HJ (1993) Selection criteria for lactic acid bacteria to be used as starter cultures in various food commodities. FEMS Microbiol Rev 12:253-272.
- Bunkova L, Bukka F, Hlobilova M, Vakatkova Z, Novakova D, Drab V (2009) Tyramine production of technological important strains of Lactobacillus, Lactococcus and Streptococcus Eur Food Res Tech 229:533-538.
- Choudhury N, Hansen W, Engesser D, Hammes WP, Holzapfel WH (1990) Formation of histamine and tyramine by lactic acid bacteria in decarboxylase assay medium. Lett Appl Microbiol 11:278-281.
- Connil N, Plissoneau L, Onno B, Pilet MF, Prevost H, Dousset X (2002) Growth of Carnobacterium divergens V41 and production of biogenic amines and divercin V41 in sterile coldsmoked salmon extract at varying temperatures, NaCl levels, and glucose concentrations. J Food Protect 65:333-338.
- Fadda S, Vignolo G, Oliver G (2001) Tyramine degradation and tyramine/histamine production by lactic acid bacteria and Kocuria strains. Biotechnol Lett 23:2015-2019.
- Gonzalez-Rodriguez MN, Sanz J, Santos JÁ, Otero A, Garcia-Lopez ML (2002) Numbers and types of microorganisms in vacuum-packed cold-smoked freshwater fish at the retail level. Int J Food Microbiol 77:161-168.
- Halász A, Barath A, Simon-Sarkadi L, Holzapfel W (1994) Biogenic amines and their production by micro-organisms in food. Trends Food Sci Tech 5:42-49.
- Jorgensen LV, Huss HH, Dalgaard P (2000) The effect of biogenic amine production by single bacterial cultures and metabiosis on cold-smoked salmon. J Appl Microbiol 89:920-934.
- Karovicová J, Kohajdová Z (2005) Biogenic amines in food. Chem Pap 59:70-79.
- Kim MK, Mah JH, Hwang HJ (2009) Biogenic amine formation and bacterial contribution in fish, squid and shellfish. Food Chem 116:87-95.
- Krizek M, Pavlicek T, Vacha F (2004) Formation of selected biogenic amines in carp meat J Sci Food Agr 82:1088-109.
- Landete JM, Ferrer S, Pardo I (2005) Which lactic acid bacteria are responsible for histamine production in wine. J Appl Microbiol 99:580-586.
- Leuschner RG, Hammes WP (1999) Formation of biogenic amine in mayonnaise, herring and tuna fish salad by lactobacilli. Int J Food Sci Nutr 50:159-64.
- Lucas PM, Wolken WA, Claisse O, Lolkema JS, Lonvaud-Funel A (2005) Histamine-producing pathway encoded on an unstable plasmid in Lactobacillus hilgardii 0006. Appl Environ Microb 71:1417-1424.
- Magwamba C (2010) Detection and quantification of biogenic amines in fermented food products aold in Botswana. J Food Protect 73:1703-8.
- Marcobal A, de las Rivas B, Muñoz R (2006) Methods for the detection of bacteria producing biogenic amines on foods: A Survey. J Verbrauch Lebensm 1:187-196.
- Marino M, Maifreni M, Bartolomeoli I, Rondinini G (2008) Evaluation of amino acid-decarboxylative microbiota throughout the ripening of an Italian PDO cheese produced using different manufacturing practices. J Appl Microbiol 105:540-549.
- Matamoros S, Pilet MF, Gigout F, Prevost H, Leroi F (2009) Selection and evaluation of seafood-borne psychrotrophic lactic acid bacteria as inhibitors of pathogenic and spoilage bacteria. Food Microbiol 26:638-644.
- Moreno-Arribas MV, Polo MC (2008) Occurrence of lactic acid bacteria and biogenic amines in biologically aged wines. Food Microbiol 25:875-881.
- Okuzumi M, Okuda S, Awano M (1981) Occurence of psychrophilic and halophilic histamne forming bacteria (N-Group Bacteria) on/in red meat fish. Bull Jpn Soc Sci Fish 48:799-804.
- Ono K, Yamamoto K (1999) Contamination of meat with Campylobacter jejuni in Saitama, Japan. Int J Food Microbiol 47:211-219.
- Özogul F (2011) Effects of specific lactic acid bacteria species on biogenic amine production by food-borne pathogen. Int J Food Sci Technol 46:478-484.
- Özogul F, Özogul Y (2005) Formation of biogenic amines by Gram-negative rods isolated from fresh, spoiled, VP-packed and MAP-packed herring (Clupea harengus). Eur Food Res Technol 221:575-581.
- Özogul F (2004) Production of biogenic amines by Morganella morganii, Klebsiella pneumoniae and Hafnia alvei using a rapid HPLC method. Eur Food Res Technol 219:465-469.
- Petaja E, Eerola S, Petaja P (2000) Biogenic amines in coldsmoked fish fermented with lactic acid bacteria. Eur Food Res Technol 210:280-285.
- Ruiz-Cappillas C, Jimenez-Colmenero F (2004) Biogenic amines in meat and meat products. Crit Rev Food Sci Nutr 44:489-499.
- Silla-Santos MH (1996) Biogenic amines: their importance in foods. Int J Food Microbiol 29:213-231.
- Stadnik J, Dolatowski ZJ (2010) Biogenic amines in meat and fermented meat products. Acta Sci Pol Technol Aliment 3:251-263.
- Stratton JE, Hutkins RW, Taylor SL (1991) Biogenic amines in cheese and other fermented foods. J Food Protect 54:460-470.
- Suzzi G, Gardini F (2003) Biogenic amines in dry fermented sausages. Int J Food Microbiol 88:41-54.
- Taylor SL (1986) Histamine food poisoning: toxicology and clinical aspects. Crit Rev Toxicol 17:91-128.
- Thapa, N, Pal J, Tamang JP (2006) Phenotypic identification and technological properties of lactic acid bacteria isolated from traditionally processed fish products of the Eastern Himalayas. Int J Food Microbiol 107:33-38.
- Udomsil N, Rodtong S, Tanasupawat S, Yongsawatdigul J (2010) Proteinase-producing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds. Int J Food Microbiol 141:186-194.
- Umezu M, Shibata A, Maeda M (1977) Production of amines by nitrate reducing-bacteria and lactobacilli for sake-brewing. Hakkokogaku 55:68-74.
- Yongsawatdigul J, Choi YJ, Udomporn S (2004) Biogenic amines formation in fish sauce prepared from fresh and temperature abused Indian anchovy (Stolephorus indicus). J Food Sci 69:312-319.
- Zaman MZ, Bakar FA, Selamat J, Bakar J (2010) Occurrence of biogenic amines and amines degrading bacteria in fish sauce. Czech J Food Sci 28:440-449.
- Zhong-Yi L, Zhong-Hai L, Miao-Ling Z, Xiao-Ping D (2010) Effect of fermentation with mixed starter cultures on biogenic amines in bighead carp surimi. Int J Food Sci Technol 45:930-936.
Send correspondence to:
Publication Dates
-
Publication in this collection
09 Oct 2013 -
Date of issue
2013
History
-
Received
23 Jan 2012 -
Accepted
23 July 2012