Abstract
Filamentous fungi are considered to be the most important group of microorganisms for the production of plant cell wall degrading enzymes (CWDE), in solid state fermentations. In this study, two fungal strains Aspergillus niger MS23 and Aspergillus terreus MS105 were screened for plant CWDE such as amylase, pectinase, xylanase and cellulases (β-glucosidase, endoglucanase and filterpaperase) using a novel substrate, Banana Peels (BP) for SSF process. This is the first study, to the best of our knowledge, to use BP as SSF substrate for plant CWDE production by co-culture of fungal strains. The titers of pectinase were significantly improved in co-culture compared to mono-culture. Furthermore, the enzyme preparations obtained from monoculture and co-culture were used to study the hydrolysis of BP along with some crude and purified substrates. It was observed that the enzymatic hydrolysis of different crude and purified substrates accomplished after 26 h of incubation, where pectin was maximally hydrolyzed by the enzyme preparations of mono and co-culture. Along with purified substrates, crude materials were also proved to be efficiently degraded by the cocktail of the CWDE. These results demonstrated that banana peels may be a potential substrate in solid-state fermentation for the production of plant cell wall degrading enzymes to be used for improving various biotechnological and industrial processes.
solid state fermentation; plant cell wall degrading enzymes (CWDE); banana peels; Aspergillus strains; enzymatic hydrolysis
RESEARCH PAPER
Production of plant cell wall degrading enzymes by monoculture and co-culture of Aspergillus niger and Aspergillus terreus under SSF of banana peels
Shazia Rehman; Hina Aslam; Aqeel Ahmad; Shakeel Ahmed Khan; Muhammad Sohail
Department of Microbiology, University of Karachi, Karachi, Pakistan
Correspondence Correspondence: M. Sohail Department of Microbiology University of Karachi 75270 Karachi, Pakistan E-mail: msohail@uok.edu.pk
ABSTRACT
Filamentous fungi are considered to be the most important group of microorganisms for the production of plant cell wall degrading enzymes (CWDE), in solid state fermentations. In this study, two fungal strains Aspergillus niger MS23 and Aspergillus terreus MS105 were screened for plant CWDE such as amylase, pectinase, xylanase and cellulases (β-glucosidase, endoglucanase and filterpaperase) using a novel substrate, Banana Peels (BP) for SSF process. This is the first study, to the best of our knowledge, to use BP as SSF substrate for plant CWDE production by co-culture of fungal strains. The titers of pectinase were significantly improved in co-culture compared to mono-culture. Furthermore, the enzyme preparations obtained from monoculture and co-culture were used to study the hydrolysis of BP along with some crude and purified substrates. It was observed that the enzymatic hydrolysis of different crude and purified substrates accomplished after 26 h of incubation, where pectin was maximally hydrolyzed by the enzyme preparations of mono and co-culture. Along with purified substrates, crude materials were also proved to be efficiently degraded by the cocktail of the CWDE. These results demonstrated that banana peels may be a potential substrate in solid-state fermentation for the production of plant cell wall degrading enzymes to be used for improving various biotechnological and industrial processes.
Key words: solid state fermentation, plant cell wall degrading enzymes (CWDE), banana peels, Aspergillus strains, enzymatic hydrolysis.
Introduction
The chemical composition of plant cell wall comprises (approximately) 25% lignin and 75% carbohydrates or sugars (Smith, 1971). The carbohydrate fraction consists of cellulose, hemicelluloses (xylan etc.) and pectin, whereas the non-fermentable polyphenolic fraction is lignin, which acts as glue, holding together the cellulose fibers. The biomass provides a renewable chemical feedstock, which is degraded using a variety of physicochemical methods. These treatment processes possess many disadvantages; therefore, biological conversion using microbial enzymes is a topic of intense research around the globe.
Currently, microbial enzymes that attack plant cell wall are widely exploited in the biotechnology sector. Mainly filamentous fungi are reportedly involved in the efficient degradation of plant cell wall as they utilize the complex plant cell wall polysaccharides as a sole source of carbon by producing a wide range of hydrolases such as cellulases, hemicellulases and amylases. Due to the development of large-scale fermentation technologies, the production of microbial enzymes accounts for a significant proportion of the biotechnology industry's total output. Among the processes used for enzyme production, solid state fermentation (SSF) is favored because of a number of advantages. The process of SSF may be defined as the fermentation process of cultivation of microorganisms on a non-soluble material which is used as a source of nutrients as well as a physical support under controlled conditions without free-flowing aqueous phase (Pandey, 1992).
A variety of traditional (agricultural products) and non-traditional (agricultural and food processing wastes) substrates can be employed in SSF. Among such wastes, Banana peel (BP) is an under-utilized substrate. Banana is one of the major and highly consumed fruit, originated from the tropical region of South Asia (Anhwange et al., 2009). Because of the high use, banana waste i.e., peels are generated in bulk and constitute up to 30-40% of the total fruit weight. Being perishable, its disposal is a matter of concern in order to alleviate environmental pollution. Since, this fruit residue contains carbohydrates, proteins, fibers in a significant amount and lignin content is in less quantity (Essien et al., 2005), it can be used as a supplementary source for the production of industrial enzymes (Oberoi et al., 2011).
The carbohydrates (CHO) which are present in huge quantities in BP include celluloses, hemicelluloses, starch and a major proportion of pectic contents. The highly rich CHO-content (Essien et al., 2005) and some other basic minerals in BP provide the cheap source for the growth of microorganisms. Earlier, Trichoderma viridae has been reported to produce cellulase from BP under solid state fermentation (Sun et al., 2011). BP as a solid medium in SSF, not only provides advantage of being cost effective for the biosynthesis of a number of industrial enzymes but also promotes substantial reduction of the agro-industrial pollution to the environment (Ponnuswamy et al., 2011).
In the present work solid-state fermentation of BP is reported for the production of multienzyme preparation by mono- and co-culture of Aspergillus strains: A. niger and A. terreus. The cultures were previously reported for their ability to produce hydrolytic enzymes (Sohail et al., 2009).
Materials and Methods
Organism and culture maintenance
Strains of Aspergillus terreus (MS105) and A. niger (MS23) were obtained from the culture collection of the Department of Microbiology, University of Karachi. The cultures were maintained on SDA slants, stored at 4 °C and sub-cultured periodically.
Substrate pretreatment
The natural substrate, Banana peels (BP), was collected from a local fruit juice shop and was used after solar drying, ground and sieving through 100 mesh sizes.
Preparation of spore suspension (Inoculum)
Fungal cultures maintained on Sabouraud Dextrose agar (SDA) slants at 4 °C, were transferred to SDA plates and incubated at 37 °C for seven days. The spores on the agar surface were gently scraped and blended in 10 mL sterile saline, vortexed and used as spore suspension (inoculum). The spores were enumerated under microscope suspended in a hemocytometer. The count was finally adjusted to 5 x 107 spores/mL.
Solid state fermentation
Solid-state fermentation (SSF) was carried out according to the modified Kang et al. (2004) method. Briefly, 2 g BP were autoclaved in a 100 mL Erlenmeyer flask at 121 °C for 30 min. To moist the solid substarte, an appropriate quantity (3.25 mL) of mineral salt medium (MSM); as prescribed by Mandels and Weber (1969), was added to maintain the desired moisture level (i.e. 65%). Each flask was then inoculated with spore suspension to obtain a final concentration of 5 x 107 spores/gm substrate, mixed and incubated at 28 °C or 37 °C and 90% humidity for one week under static condition.
Extraction and storage of crude enzymes
After incubation, 40 mL of 50 mM sodium citrate buffer pH 4.8; containing 0.05% (w/v) tween-80 was added to each flask and kept under agitation at 150 rpm for 2 h. It was followed by the separation of fungal biomass and culture medium residues using four layers of Whatman#1 filter paper. Filtrate was centrifuged at 6000 rpm for 20 min to obtain clear cell-free culture supernatant (CFCS). CFCS was used as crude enzyme preparation and stored at 4 °C until used.
Enzyme assays
Activities of xylanase, pectinase, amylase, β-glucosidase, endoglucanase and filterpaperase in CFCS were determined by protocol described by Sohail et al. (2009). Briefly, crude enzyme preparation (25 µL) was incubated in the presence of 25 µL substrate; birhcwood xylan (0.5% w/v) for xylanase, citrus pectin (1% w/v) for pectinase, starch (0.5% w/v) for amylase, salicin (1% w/v) for β-glucosidase, carboxymethyl cellulose (1% w/v) for endoglucanase and Whatman # 1 filter paper strip (3 cm x 0.5 cm) for filterpaperase, in 50 mM Sodium citrate buffer, pH 4.8, at 50 °C for 30 min. Amount of reducing-sugar was determined by DNS method (Miller, 1969) against standard curves of xylose, galacturonic acid and glucose. One International Unit (IU) of enzyme was defined as the amount of enzyme that releases one micromole of reducing-sugar per minute under standard assay conditions.
Effect of supplementations on enzyme production
To analyze the effect of various inducers, BP was supplemented with xylan (0.5%), pectin (1%), starch (0.5%) and CMC (1%) for multi-enzyme production under the process of solid state fermentation.
Hydrolysis of crude and commercially available substrates
For hydrolysis, different crude substrates (i.e., banana peels, lime peels, potato peels, sugar-cane bagasse, corn leaves) and purified substrates (i.e., xylan, pectin, starch, avicel, CMC) were used. Crude substrates were treated as described for banana peels earlier. To the 100 mL Erlenmeyer flask, 0.5 g of substrate in 50 mM sodium citrate buffer pH 4.8 containing 0.02% sodium azide (antibacterial) and 5 IU of crude enzyme preparation were added, incubated at 50 °C for 26 h with agitation at 150 rpm in an orbital shaker. Samples were taken periodically and amount of reducing sugars were measured by DNS method (Miller, 1969) against standard curve of glucose.
Results and Discussions
The present study was carried out using banana peels (BP) as a novel solid support for microbial anchorage for higher levels of enzyme production in SSF. Previously, BP was reported as a substrate for the production of cellulase by Trichoderma (Sun et al., 2011), amylase by Penicillium sp. (Ponnuswamy et al., 2011), laccase by A. fumigatus (Vivekanand et al., 2011) under SSF processes. Since BP is enriched with carbohydrates, proteins, fats, dietary fibers and other basic nutrients (Essien et al., 2005) that can support fungal growth, therefore, the potential of Aspergillus strains was evaluated for the production of plant cell wall degrading enzymes using this substrate as growth medium. To our knowledge this is the first report describing the production of multi-enzyme preparation by mono- and co-culture under SSF of BP.
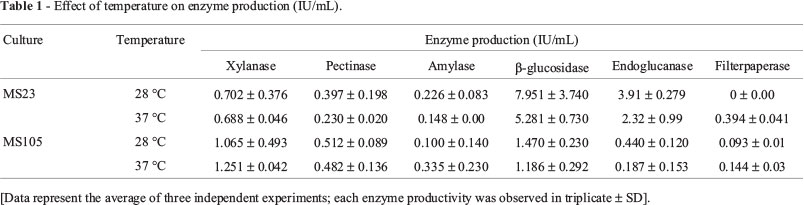
Temperature is one of the important factors which strongly affect the SSF process (Pandey et al., 2000).The cultivation temperature does not only affect the growth rate of organisms but it also has marked effects on the level of enzyme production. Therefore, this study was carried out at two different cultivation temperatures (i.e. 28 °C and 37 °C) in order to determine the optimum growth and enzyme production.
The strain MS23 gave maximum productivity of almost all the investigated enzymes in this work at 28 °C which decreased upon increasing the temperature up to 37 °C (Table 1). Haltrich et al. (1996) in their study on fungal xylanases reported that A. niger grew well at 28 °C and xylanase production was significantly decreased at higher temperature (40 °C). Sun et al. (2011) also reported about higher production of cellulases under SSF of banana peels at 30 °C compared to at 40 °C. It might be due to that at temperatures higher than optimum, the growth of the fungus was inhibited and hence the enzyme production (Yaun and Rungyu, 1999; Rahman et al., 2003).
Whereas, the optimum temperature for the enzyme production by A. terreus was found to be 37 °C (Table 1) that was in line with Naseeb (2007). Gawande et al. (1999) also reported enhanced productivity of xylanase at 35 °C by A. terreus.
The heat generated from the metabolic activities of the fungi, growing on the specific solid substrate, also influence the SSF process, together with the environmental temperature. Mrudula and Anitharaj (2011) reported maximum pectinase production on orange peels under SSF by A. niger at the cultivation temperature 50 °C. Moretti et al. (2012) also recorded that the maximum activities were observed at 60 °C for xylanase and 70 °C for CMCase from A. fumigatus M.7.1 using sugar-cane bagasse and wheat bran (1:1 w/w) as SSF substrate. But in our study, enhanced production of pectinase by A. niger was obtained at 28 °C using banana peels. This can be attributed towards different substrate composition used for SSF processes and different strains used for plant cell wall degradation.
Various workers have used another strategy to increase the industrially important enzymes under SSF; they supplemented the solid substrate with one or more nutritional component that may act as an enzyme inducer. For instance, Wang et al. (2012) used α-cellulose as a model substrate on corn cob to induce the cellulase production by A. fumigatus ECU0811. Enhanced production of amylase and xylanase was observed by Singh et al. (2012) using Streptomyces sp. MSC702, when the production medium was mixed with wheat bran and rice bran. Dhillon et al. (2012) also studied the effect of various inducers (lactose, CuSO4, veratryl alcohol) on the rapid bioproduction of cellulase and hemicellulase using apple pomace as substrate. In the current study, BP was also supplemented with different carbon sources such as starch, birchwood xylan, citrus pectin and carboxymethyl cellulose (CMC) which resulted in the induction and repression of enzyme production in SSF. Almost all the carbon sources tested in this study positively enhanced the enzyme synthesis, proving BP a promising substrate for CWDE production.
The supplementation of xylan to BP enhanced the production of xylanase, pectinase and FPase by strain MS23 compared to the BP without supplementation (Table 2). Nonetheless, production of xylanase, amylase and β-glucosidase decreased upon similar supplementation by the strain MS105 (Table 3). Kang et al. (2004) has reported the nature of the substrate used in fermentation affects the induciblity of cellulases and hemicellulases. Gupta and Kar (2008) reported that 0.5% increment of xylan to corn cob and wheat bran induced xylanase production by Bacillus sp.
In this study, the addition of CMC to BP resulted in enhanced production of endoglucanase and FPase by MS105 (Table 3). This finding is in agreement with Sohail et al. (2011) where presence of 1% (w/v) CMC in the medium increased the titers of FPase and endoglucanase by Alternaria sp. Aslam et al. (2010) has also mentioned the increase in the levels of endoglucanase when Trichoderma sp. grew in the presence of CMC indicating that it is the substrate of choice for endoglucanase production.
It is generally agreed that the optimum production of pectinase can be achieved in the medium containing pectic materials as an inducer (Hang and Woodanms, 1994; Solis-Pereira et al., 1993; Naidu and Panda, 1998). Our study affirms this fact as the supplementation of citrus pectin to the banana peels resulted in substantial increase in pectinase production by both the strains tested (Tables 1 and 2). Furthermore, this substrate favored the production of other polysaccharide-degrading enzymes, as well, when supplemented.
On the other hand, starch supplementation to BP under SSF, decreased the yield of CWDE particularly pectinase by MS105. It could be attributed to the repression mechanism brought about by the degradation of starch into glucose by the activity of amylase in the medium. Similar results of catabolite repression of enzyme production by glucose, has been reported by Nandakumar et al. (1999) for A. niger CFTRI 1105 and Alva et al. (2007) for Aspergillus sp. JGI 12. In contrast to the finding for MS105, our studies on MS23 showed that starch increment significantly increased the enzyme production by this strain (Table 2), indicating a different genetic regulatory mechanism.
Co-culturing is another strategy to improve enzyme production by industrially important strains. Since the two strains investigated in this study showed variation in the titers and proportion of different enzymes in their preparations, so these were co-cultured under SSF of BP. As the higher production of enzymes from MS23 was noted at 28 °C and from MS105 at 37 °C, therefore, the strains were co-cultured at these two temperatures and the productivity was compared with monocultures.
Results of co-culture studies demonstrated that pectinase was over-produced at 28 °C compared to the mono culture. The highest production of this enzyme by Aspergillus strains was observed when BP was supplemented with xylan at 28 °C (Table 4). Wang et al. (2006) also reported the increased production of cellulase, hemicellulase, glucoamylase and pectinase when mixed culture of Aspergillus niger was used under SSF process using bran as the main substrate. Elevated production of pectinases in the mixed culture is quite economical and of great commercial importance for various food industry applications; to increase the juice yield and its clarity, preventing haziness of wine in brewing industries, extraction of oil, fermentation of coffee and tea and in the preparation of animal feed (Jayani et al., 2005; Gummadi et al., 2003; Aguilar and Huitron, 1986). This mixed culture strategy not so effective in the production of other hydrolytic enzymes when compared with the findings of single strains production (Table 4 and 5), showing lower synergism among the two molds. Similar results were obtained by Massadeh et al. (2001) that the addition of A. terreus to T. reesei did not increase the reducing sugar concentration compared to single culture using sugarcane bagasse in solid substrate fermentation, showed that mixed culture resulted in a decrease in substrate degradation.
The fungal CWDE are frequently applied for the saccharification and hydrolysis of crude lignocellulosic substrates. The simple sugars released by saccharification process can then be utilized in a number of industrial and commercial processes. In the present study, the enzyme preparations obtained by SSF of BP were studied for their potential to degrade BP and other crude substrates along with some purified substrates.
Our studies revealed that pectin was maximally hydrolyzed after 26 h and produced a higher quantity of reducing sugars (Table 6) in the enzymatic preparation of both the fungal strains (mono and co-culture), attributing the highest activity of pectinase in these loadings.
Crude substrates took more time compared to purified substrates for efficient enzymatic hydrolysis. Although MS23 produced large amount of cellulases, but its enzyme preparation could not degrade xylan and CMC (Figure 1A) to a greater extent. It may be due to the complex nature of their structures that prevent easy access of enzymes or the enzyme preparation did not contain the optimum ratio of cellulolytic and xylanolytic components to degrade these substrates.

It was also found that the enzyme produced by the strain MS23 was unable to act upon lime peels as no reducing activity was detected in the reaction mixture containing lime peels (Figure 1B). This inability of enzymes can be explained by taking the complex and heterogeneous structure of plant cell walls in consideration. The literature survey revealed that enzymes extracted from A. niger is much more sensitive towards the substrate structure in comparison to that extracted from T. ressei (Zuhair, 2008).
Contrary to above findings, the Aspergillus sp. MS105 hydrolyzed both types of substrate i.e. crude and purified, to a greater extent (Figures 2A and 2B). Although, crude substrates took longer time but produced more amount of reducing sugars than purified substrates (Table 6). It implies that the synergistic action of the plant cell wall degrading enzymes has been found to be most effective for hydrolysis of crude substrates but not for purified substrates.

Our co-culture data showed that the enzyme cocktail of MS23 and MS105 did not hydrolyze crude substrates efficiently in comparison to the enzyme preparation from the monocultures (Figures 3A and 3B). Amongst purified substrates, the hydrolysis of pectin yielded the maximum reducing sugars (0.074 g) while the hydrolysis of xylan contributed the least (0.007 g) (Table 6). As explained earlier, it may be due to higher pectinolytic and low xylanolytic activities in enzyme loadings.
Conclusion
Production of plant cell wall degrading enzymes by the Aspergillus strains MS23 and MS105 under SSF of BP, suggests an extensive amount of CWDE in monocultures and higher degree of pectinases in co-culture. To strengthen the biotechnological industries, this could be an effective approach by ultimately purifying the increased enzymes from this cocktail. Overall, these results support the use of BP as an excellent low-cost, promising substrate for the induction of multi-enzymes production under SSFprocesses.
Acknowledgments
The authors gratefully thank Dean Faculty of Science, University of Karachi, for financial assistance in pursuing this research program.
Submitted: July 4
Approved: April 17, 2014
All the content of the journal, except where otherwise noted, is licensed under a Creative Commons License CC BY-NC.
- Aguilar G, Huitron C (1986) Application of fed-batch cultures in the production of extracellular pectinase by Aspergillus specie. Enzyme Microb Technol 8:541-545.
- Alva S, Anupama J, Salva J, Chiu YY, Vyshali P, Shruti M, Yogeetha BS, Purvi J, Bhavya D, Ruchi K, Kumudini BS, Varalakshmi KN (2007) Production and characterization of fungal amylase enzyme isolated from Aspergillus sp JGI 12 in solid state culture. Afr J Biotechnol 6:576-581.
- Anhwange BA, Ugye TJ, Nyiaatagher TD (2009) Chemical composition of Musa sapientum (Banana) peels. EJEAF Che 8:437-442.
- Aslam N, Sheikh MA, Ashraf M, Jalil A (2010) Expression pattern of Trichoderma cellulases under different carbon sources. Pak. J Bot 42:2895-2902.
- Dhillon GS, Kaur S, Brar SK, Verma N (2012) Potential of apple pomace as a solid substrate for fungal cellulase and hemicellulase bioproduction through solid state fermentation. Ind Crops & Products 38:6-13.
- Essien JP, Akpan EJ, Essien EP (2005) Studies on mould growth and biomass production using waste banana peel. Bioresour Technol 96:1451-1456.
- Gawande PV, Kamat MY (1999) Production of Aspergillus xylanase by lignocellulosic waste fermentation and its application. J Appl Microbiol 87:511-519.
- Gummadi SN, Panda T (2003) Purification and biochemical properties of microbial pectinases - A review. Process Biochem 38:987-996.
- Gupta U, Kar R (2008) Effect of inducers on growth & xylanase production by a thermophilic Bacillus isolate on agro-residues and characterization of kinetic parameters. Int J Biotechnol & Biochem 4:347-358
- Haltrich D, Nidetzky B, Kulbe KD, Steiner W, Zupancic S (1996) Production of fungal xylanases. Bioresour Technol 58:137-161.
- Hang YD, Woodanms EE (1994) Production of fungal polygalacturonase from apple pomace. Lebensm Wiss U Technol 27:194-196.
- Jayani RS, Saxena SR, Gupta (2005) Microbial pectinolytic enzymes: A review. Process Biochem 40:2931-2944.
- Kang SW, Park YS, Lee JS, Hong SI, Kim SW (2004) Production of cellulases and hemicellulases by Aspergillus niger KK2 from lignocellulosic biomass. Bioresour Technol 91:153-156.
- Mandels M, Weber J (1969) The production of cellulases and their applications. Adv Chem Ser 95:391-414.
- Massadeh MI, Yusoff WMW, Omar O, Kader J (2001) Synergism of cellulose enzymes in mixed culture solid substrate fermentation. Biotechnol Lett 23:1771-1774.
- Miller GC (1969) Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Anal Chem 31:426-428.
- Moretti MMS, Bocchini-Martins DA, Silva RD, Rodriguez A, Sette LD, Gomes E (2012) Selection of thermophilic and thermotolerant fungi for the production ofcellulases and xylanases under solid-state fermentation. Braz J Microbiol 2012:1062-1071.
- Mrudula S, Anitharaj R (2011) Pectinase production in solid state fermentation by Aspergillus niger using orange peel as substrate. Global J Biotech and Biochem 6:64-71.
- Naidu GSN, Panda T (1998) Production of pectolytic enzymes - A review. Process Eng19:355-361.
- Nandakumar MP, Thakur MS, Raghavaro KSMS, Ghildyal NP (1999) Studies on catabolite repression in solid state fermentation for biosynthesis of fungal amylases. Lett Appl Microbiol 29:380-384.
- Naseeb S (2007) Studies on fungal xylanases. Karachi, Pakistan. M.Sc Dissertation, Department of Microbiology, University of Karachi.
- Oberoi HS, Vallani PV, Saida L, Bansal S, Hughes JD (2011) Ethanol production from banana peels using statistically optimized simultaneous saccharification and fermentation process. Waste Management 31:1576-1584.
- Pandey A (1992) Recent progress developments in solid state Fermentation. Process Biochem 27:109-117.
- Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process Biochem 35(10):1153-1169.
- Ponnuswamy V, Devi VSL, Vincent SGP (2011) Bioprocessing of banana peel for production of amylase by Penicilliun sp Asian J Exp Biol Sci 2:257-264.
- Rahman AK, Sugitani N, Hatsu M, Takamizawa K (2003) A role of xylanase, alpha-L-arabinofuranosidase, and xylosidase in xylan degradation. Canadian J Microbiol 49:58-64.
- Singh R, Kapoor V, Kumar V (2012) Utilization of agroindustrial wastes for the simultaneous production of amylase and xylanase by thermophilic actinomycetes. Braz J Microbiol 2012:1545-1552.
- Smith GS (1971) Plant cell wall structure and cell wall growth. Tuatara 19:43-50.
- Sohail M, Ahmad A, Khan SA (2011) Production of cellulases from Alternaria sp. MS28 and their partial characterization. Pak J Bot 43:3001-3006.
- Sohail M, Siddiqi R, Ahmad A, Khan SA (2009) Cellulase production from Aspergillus niger MS82: Effect of temperature and pH. New Biotechnol 25:437-441.
- Solis-Pereira S, Favela Torres E, Viniegra-Gonzales G, Gutierrz- Rojas M (1993) Effect of different carbons sources on the synthesis of pectinase by Aspergillus niger in submerged and solid state fermentation. Appl Microbiol Biotechnol 39:36-41.
- Sun HY, Li J, Zhao P, Peng M (2011) Banana peel: A novel substrate for cellulase production under solid-state fermentation. Afr J Biotechnol 10:17887-17890.
- Vivekanand V, Dwivedi P, Pareek N, Singh RP (2011) Banana peel: A Potential Substrate for Laccase Production by Aspergillus fumigatus VkJ2.4.5 in Solid-State Fermentation. Appl Biochem Biotechnol 165(1):204-20.
- Wang D, Sun J, Yu HL, Li CX, Bao J, Xu JH (2012) Maximum saccharification of cellulose complex by an enzyme cocktail supplemented with cellulose from newly isolated Aspergillus fumigates ECU 0811. Appl Biochem Biotechnol 66(1):176-186.
- Wang XJ, Bai JG, Liang YX (2006) Optimization of multienzyme production by two mixed strains in solid state fermentation. Appl Microbial Biotechnol 73:533-540.
- Yaun Q, Rungyu M (1999) Study on temperature oscillation in production of xylanase by Aspergillus niger Beijing Hugagong Daxue Xuebao 26:11-16.
- Zuhair SA (2008) The effect of crystallinity of cellulose on the rate of reducing sugars produced by heterogenous enzymatic hydrolysis. Biores Technol 99:4078-4085.
Publication Dates
-
Publication in this collection
13 Feb 2015 -
Date of issue
Dec 2014