Abstract
Copper mine drainages are restricted environments that have been overlooked as sources of new biocatalysts for bioremediation and organic syntheses. Therefore, this study aimed to determine the enzymatic activities (esterase, epoxide hydrolase and monooxygenase) of 56 heterotrophic bacteria isolated from a neutral copper mine drainage (Sossego Mine, Canaã dos Carajás, Brazil). Hydrolase and monooxygenase activities were detected in 75% and 20% of the evaluated bacteria, respectively. Bacterial strains with good oxidative performance were also evaluated for biotransformation of organic sulfides. Fourteen strains with good enzymatic activity were identified by 16S rRNA gene sequencing, revealing the presence of three genera: Bacillus, Pseudomonas and Stenotrophomonas. The bacterial strains B. megaterium (SO5-4 and SO6-2) and Pseudomonas sp. (SO5-9) efficiently oxidized three different organic sulfides to their corresponding sulfoxides. In conclusion, this study revealed that neutral copper mine drainages are a promising source of biocatalysts for ester hydrolysis and sulfide oxidation/bioremediation. Furthermore, this is a novel biotechnological overview of the heterotrophic bacteria from a copper mine drainage, and this report may support further microbiological monitoring of this type of mine environment.
Keywords:
Copper mine drainage; High-throughput screening; Hydrolases; Monooxygenases
Introduction
Brazil is the world's 15th largest producer of copper concentrate.11 ICSG – The International Copper Study Group. The World Copper Factbook 2013. Lisbon: ICSG; 2013. The mining company Vale S.A. is responsible for the majority of copper production, having reached 275,000 mt of copper concentrate in the first nine months of 2013,22 Blamey A. Brazil's Vale targets 289,000 mt nickel, 405,000 mtcopper in 2014. Platts; 2013. Available at: http://www.platts.com/latest-news/metals/london/brazils-vale-targets-289000-mt-nickel-405000-26507146. Accessed 28.01.14.
http://www.platts.com/latest-news/metals...
and operates at the Sossego Mine (Canaã dos Carajás, PA, since 2004) and at the Salobo Mine (Marabá, PA, since 2012). The process used for copper ore concentration involves crushing and subsequent semi-autogenous grinding (SAG), followed by comminution in a ball mill. The material is classified according to the particle size and then passes through a flotation process. The generated waste (a slurry) is deposited in a dam approximately 5200 m in length.
The aqueous wastes generated from mining activities are known as mine drainages and are described as restricted environments that often have a high metal content and low organic matter concentration. In addition, water drainage from mine wastes and abandoned mines are often acidic as a result of the extended exposure of sulfidic minerals to water and oxygen. However, the pH of copper mine drainage may drastically differ according to the waste chemical composition, the time of exposure and the microbial community.33 Johnson DB. Chemical and microbiological characteristics of mineral spoils and drainage waters at abandoned coal and metal mines. Water Air Soil Pollut. 2003;3:47-66.
Most microbial studies focus on the mineral-oxidizing prokaryotes from acid mine drainages (AMD), which usually consist of acidophilic bacteria and archaea. These microorganisms are able to oxidize ferrous iron and/or reduced forms of sulfur and accelerate the oxidative dissolution of sulfidic minerals, which may be applied to metal ore processing and concentrates in a biotechnological process known as biomining and bioleaching.44 Falteisek L, Čepička I. Microbiology of diverse acidic and non-acidic microhabitats within a sulfidic ore mine. Extremophiles. 2012;16:911-922.
5 Watling HR, Watkin ELJ, Ralph DE. The resilience and versatility of acidophiles that contribute to the bio-assisted extraction of metals from mineral sulphides. Environ Technol. 2010;31:915-933.-66 Hallberg KB. New perspectives in acid mine drainage microbiology. Hydrometallurgy. 2010;104:448-453.
However, the microbial community from copper mine drainages (acidic or neutral) has been overlooked as a source of new biocatalysts for bioremediation and the synthesis of organic compounds. To the best of our knowledge, there are no reports about the use of heterotrophic microorganisms isolated from these restricted environments as biocatalysts applied for bioremediation of organic pollutants or for synthetic organic chemistry.
From a biotechnological point of view, esterases, epoxide hydrolases and monooxygenases are some of the best-studied and most-applied enzymes. These biocatalysts are commonly used for the enantioselective production of value-added compounds, which are important to pharmaceutical and fine chemical industries. In this context, our research group has been working on the detection of enzymatic activity using fluorogenic probes and organic substrates of interest. Our focus has been on new biocatalysts from the Amazon and Atlantic rainforest,77 Mantovani SM, Oliveira LG, Marsaioli AJ. Whole cell quick-E for epoxide hydrolase screening using fluorescent probes. J Mol Catal B: Enzym. 2008;52-53:173-177. petroleum oil and formation water,88 Cruz GF, Angolini CFF, Oliveira LG, et al. Searching for monooxygenases and hydrolases in bacteria from an extreme environment. Appl Microbiol Biotechnol. 2010;87:319-329. human skin,99 Silva CP. Enzymatic Potential of the Human Skin Microbiota and its Effects on Fragrance Ingredients. University of Campinas;2012 [Ph.D. thesis]. and from Brazilian Culture Collections.1010 Gonçalves RAC, Porto ALM, Pinheiro L, Cagnon JR, Manfio GP, Marsaioli AJ. Multibioreaction methodology for Baeyer-Villiger monooxygenase monitoring. Food Technol Biotechnol. 2004;42:355-361.,1111 Sicard R, Chen LS, Marsaioli AJ, Reymond J-L. A fluorescence-based assay for Baeyer-Villiger monooxygenases, hydroxylases and lactonases. Adv Synth Catal. 2005;245:1041-1050.
The present work aimed to investigate hydrolases and monooxygenases in heterotrophic bacteria isolated from a neutral copper mine drainage, applying fluorescence-based high-throughput screening (HTS) assays and multibioreactions to monitor organic sulfide oxidations. Therefore, this study represents a new biotechnological application of poorly investigated, heterotrophic microbiota.
Materials and methods
General methods
Reagents were purchased from Sigma-Aldrich (Steinheim, Germany). Solvents were distilled from technical solvents. Fluorogenic probes and products for HTS assays (2-16) were synthesized by our group.1212 Bicalho B, Chen LS, Grognux J, Reymond JL, Marsaioli AJ. Studies on whole cell fluorescence-based screening for epoxide hydrolases and Baeyer-Villiger monooxygenases. J Braz Chem Soc. 2004;15:911-916. A free sample of 2-methyl-4-propyl-1,3-oxathiane (19) from Givaudan (Jaguaré, SP, Brazil) was provided by Natura perfumery and cosmetic industry (Cajamar, SP, Brazil). Ethyl phenyl sulfide (18) and oxidation products of substrates 18, 19 and 20 (sulfoxides 18a, 19a and 20a, respectively) were synthesized as described by Porto et al. (2002).1313 Porto ALM, Cassiola F, Dias SLP, et al. Aspergillus terreus CCT 3320 immobilized on chrysotile or cellulose/TiO2 for sulfide oxidation. J Mol Catal B: Enzym. 2002;19-20:327-334. All chemical reactions were monitored by silica gel TLC (aluminum foil, 60 F254 Merck), and visualization was obtained using UV or by spraying with p-anisaldehyde/sulfuric acid followed by heating at approximately 120 °C. Flash column chromatography was performed using Merck (Whitehouse Station, NJ, USA) silica gel 60 (0.04-0.063 mm, 230-400 mesh). NMR spectra were recorded on a Varian Inova 500 (Palo Alto, CA, USA) for 1H (499.88 MHz) and 13C (125.69 MHz) measurements. Chemical shifts (δ) are given in ppm, and coupling constants (J) are given in Hertz.
Enzymatic reactions were monitored by GC-MS using an Agilent 7890 gas chromatograph (Santa Clara, CA, USA) coupled with a Hewlett Packard 5975C-MSD (70 eV) spectrometer equipped with a fused silica capillary column (HP-5MS, 30 m × 0.25 mm i.d. × 0.25 µm film thickness). GC-MS analyses were conducted using a 1 mL min-1 He flow, split mode (20:1) and the following temperature program: initial temperature 50 °C, increasing at 10 °C min-1 to 200 °C and at 20 °C min-1 to 300 °C, remaining constant for 5 min. Enantiomer discrimination was performed on an Agilent 6850 gas chromatograph (Santa Clara, CA, USA) coupled with a flame ionization detector (GC-FID) using a fused silica capillary column Lipodex-E (Macherey-Nagel Inc., Bethlehem, PA, USA) with chiral phase Octakis-(2,6-di-O-pentyl-3-O-butyryl)-γ-cyclodextrin (28 m × 0.25 mm i.d. × 0.25 µm film thickness). The GC-FID analyses were conducted using a 1 mL min-1 H2 flow, split mode (10:1) and the following temperature program: initial temperature 50 °C, increasing at 10 °C min-1 to 100 °C and at 5 °C min-1 to 180 °C, remaining constant for 5 min.
Sample collection and isolation of heterotrophic bacteria
Samples from the Sossego copper mine drainage, Canaã dos Carajás, State of Pará, Brazil, were collected in August 2009. The samples were named SO5, SO6 and SO7; collected in sterile disposable flasks; and stored at 4 °C. The Sossego mining operation began in 2004. Thus, these are 5-year-old copper mine drainage samples.
The bacteria were isolated according to Eaton and Franson (2005).1414 Eaton AD, Franson MH. Standard Methods for the Examination ofWater and Wastewater. 21st ed. Washington, DC: AmericanPublic Health Association; 2005. Each sample (1 g) was homogenized with sterile water (9 mL) and serially diluted. One milliliter of each dilution was transferred to a Petri dish and homogenized with 12 mL of Plate Count Agar (PCA). The plates were incubated for 48 h at 35 °C. Colonies exhibiting a different morphology and color were re-isolated in trypticase soy agar (TSA). The bacteria were stored at -70 °C in LB medium containing glycerol.1515 Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989.
Fluorescence-based high-throughput screening assays
Bacteria were inoculated into LB solid medium1515 Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989. and incubated for 16 h at 37 °C. After growth, cell suspensions (0.2 mg mL-1) were prepared in borate buffer (20 mmol L-1 pH 7.4).
Assays were performed in flat-bottom polypropylene 96-well microtiter plates, which were incubated at 28 °C and 200 rpm. Fluorescence intensities were measured using a plate reader spectrophotometer (FlashScan 530 Analytic Jena) at 460 nm (excitation wavelength: 390 nm).
Hydrolase screening was performed using fluorogenic probes 2-6 (Scheme 1, 100 µmol L-1), NaIO4 (2 mmol L-1), BSA (2 mg mL-1) and bacterial cell suspension (0.1 mg mL-1) and was monitored up to 24 h. Monooxygenases were investigated using probes 7-10 (Scheme 1, 100 µmol L-1), BSA (2 mg mL-1) and bacterial cell suspension (0.1 mg mL-1) and were monitored up to 72 h. All enzymatic reactions were followed by positive controls, including hydrolyzed or oxidized fluorogenic products 11-16 instead of probes, and using negative controls developed in the absence of bacterial cells.
Fluorescence-based HTS assay for detection of esterases (EST, 2-4), epoxide hydrolases (EH, 5-6) and monooxygenases (BVMO, 7-10).
Substrate conversion (%) into product was calculated by comparing the fluorescence intensities of the reaction assay and positive control, considering the latter as 100%. Negative controls were used to monitor spontaneous probe hydrolysis or oxidation. Enzymatic activity was considered positive for conversions greater than 5%.
Multibioreaction assays
Bacteria were grown in LB liquid medium for 16 h at 37 °C and 200 rpm. Cells were recovered by centrifugation at 4500 × g for 15 min. Wet bacterial cells (0.25 g) were mixed with Sørensen buffer1616 Sørensen SPL. Enzymstudien: II. Mitteilung. Über die Messung und die Bedeutung der Wasserstoffionenkoncentration bei enzymatischen Prozessen. Biochem Z. 1909;21:131-304. (5 mL, 100 mmol L-1, pH 7.0) containing 1.5 µL of each substrate (18-21) in Erlenmeyer flasks (25 mL). The resulting solutions were incubated at 37 °C and 200 rpm. After 72 and 120 h, the reactions were interrupted by saturation with NaCl and extraction with ethyl acetate (2 × 2 mL). The organic phases were combined, dried over anhydrous MgSO4, derivatized with diazomethane, and an internal standard was added (benzophenone; 0.05 mg mL-1). All samples were analyzed by GC-MS and chiral GC-FID. Substrate stability and volatility under reaction conditions were monitored by reaction controls in the absence of bacterial cells.
Substrate conversions (%) were calculated by comparing the chromatographic peak areas of the reactants, products and internal standards.
Identification of bacteria
The bacterial strains displaying hydrolytic activity above 99% or monooxygenases capable of oxidizing organic sulfur compounds were identified by sequencing of 16S rRNA. Genomic DNA was isolated using the Wizard Genomic DNA Purification Kit (Promega), following the manufacturer's instructions. PCR amplification and sequencing of partial 16S rRNA genes were conducted as described by Miqueletto et al. (2011).1717 Miqueletto PB, Andreote FD, Dias ACF, Ferreira JC, Dos Santos Neto EV, Oliveira VM. Cultivation-independent methods applied to the microbial prospection of oil and gas in soil from a sedimentary basin in Brazil. AMB Express. 2011;1:1-16. The sequences obtained were compared with sequences available at GenBank (http://www.ncbi.nlm.nih.gov) and the Ribosomal Database Project (http://www.cme.msu.edu/RDP/html/index.html). Multiple alignments of 16S rRNA gene sequences were performed using ClustalW 1.6,1818 Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673-4680.
https://doi.org/10.1093/nar/22.22.4673...
and evolutionary analyses were conducted in the Molecular Evolutionary Genetics Analysis (MEGA) 6.06 package,1919 Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol. 2013;30:2725-2729. using the DNA substitution model Kimura 2-parameter.2020 Kimura M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111-120. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 0.5288)).2121 Yang Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: approximate methods. J Mol Evol. 1994;39:306-314. The phylogenetic reconstruction was performed using the Maximum Likelihood method with 1000 bootstrapped replicates.
Results
Enzymatic activity detected by HTS
The 56 heterotrophic bacteria isolated from the three samples of a neutral copper mine drainage were evaluated by fluorescence-based HTS screening using probes 2-10 (Scheme 1).
Assays performed in the presence of probes 2 and 3 (Table 1), which were used to detect esterases catalyzing the hydrolysis of short chain esters (acetate and propionate, respectively), indicated that 27 bacteria were able to hydrolyze probe 2, five of which converted more than 99% of the substrate (SO5-6, SO5-13, SO7-7, SO7-10 and SO7-15). Probe 3 was also hydrolyzed by 27 bacteria, and SO6-12 displayed outstanding performance, converting more than 99% of the substrate.
Esterase and monooxygenase activities of heterotrophic bacteria isolated from a neutral copper mine drainage detected by fluorescence-based HTS assays using probes 2-4a a See Scheme 1. and 7-10.a a See Scheme 1.
The results for short-chain ester biohydrolysis can be divided into four distinct groups: (a) 15 bacteria that exclusively catalyzed the hydrolysis of acetate esters (2), (b) 15 bacteria that exclusively hydrolyzed propionate esters (3), (c) 12 bacteria that were not able to discriminate between 2 and 3, and (d) 14 bacteria with no esterase activity, as determined using the tested fluorogenic probes.
The heterotrophic bacteria evaluated in this work clearly prefer short-chain esters as only two strains (SO5-7 and SO7-2) were able to hydrolyze the medium-chain ester 4 (Table 1).
None of the strains were able to hydrolyze 5 and 6, which were specific for epoxide hydrolases, after a 24 h reaction period. Activity toward these probes was observed only after 72 h (data not shown); however, the conversions were below 10%, and these biocatalysts were not considered any further.
Monooxygenase screening using fluorogenic probes 7-10 indicated that among the 56 strains under evaluation, 11 were active (Table 1). Strains SO5-2, -18, SO6-12 and SO7-2 catalyzed the conversion of cyclic ketone 7 into 13. Strains SO5-1, -4, -5 and SO6-2 catalyzed the oxidation of 8 to lactone 14, and strains SO5-2, -5, -7, -9 and -10 oxidized ketone 9 to ester 15. None of the strains oxidized probe 10.
Biocatalytic sulfide oxidation by multibioreaction assays
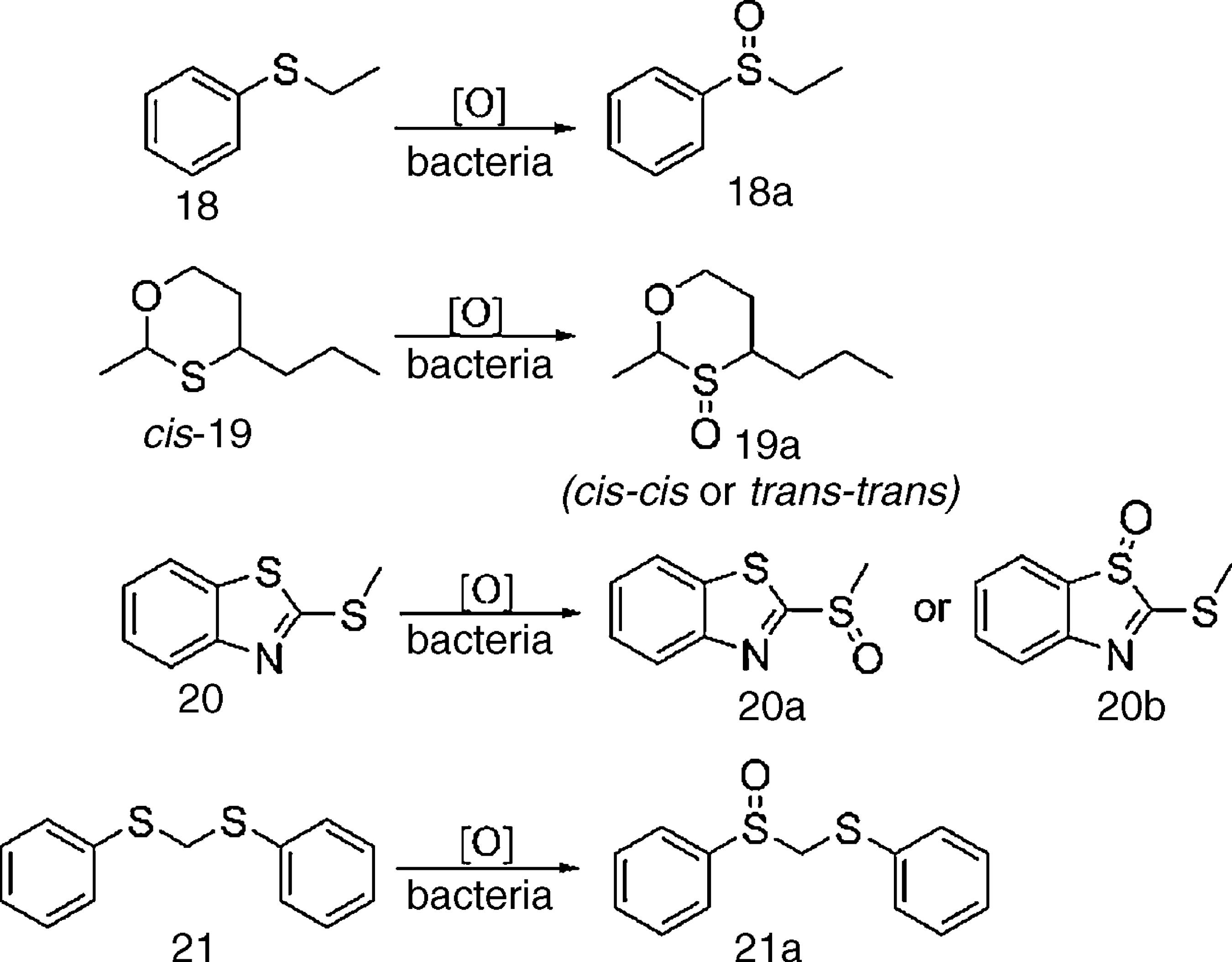
Multibioreaction experiments (Table 2) revealed that bacteria isolated from a neutral copper mine drainage were readily able to oxidize organic sulfur compounds to their respective sulfoxides (Scheme 2). All tested strains were able to catalyze the oxidation of at least one of the substrates. Eight strains oxidized 18, and three strains oxidized 19 and 20; however, none of the strains oxidized 21. The best results were observed for strains SO5-4, SO5-9 and SO6-2, which were able to oxidize three of the sulfides tested (18, 19 and 20).
Chromatographic conversion (%) of multibioreaction assays for sulfide (18-21) oxidation,a a See Scheme 2. catalyzed by bacteria from a neutral copper mine drainage.
Substrates (18-21) used for detection of organic sulfide biooxidation and their respective, expected products (18a-21a).
The enantioselectivities of the sulfide biooxidations catalyzed by SO5-4, SO5-9 and SO6-2 were assessed by chiral GC-FID, revealing low enantiomeric excesses (from 5 to 15%, data not shown). Nonetheless, oxidation of cis-19 was highly diastereoselective (over 99% de), providing only a racemic mixture of trans-trans-19a (2S,3S,4S and 2R,3R,4R, Scheme 3).
Diastereoselective biooxidation of sulfide rac-cis-19 catalyzed by bacteria SO5-4, SO5-9 and SO6-2.
Isolation and identification of heterotrophic bacteria from mine environments
All of the evaluated copper mine drainage samples had a pH between 7 and 8. Thus, specific low pH culture media for isolation of acidophilic bacteria were not used. Heterotrophic bacteria (56 strains) were isolated from the Sossego copper mine drainage samples (18 strains were isolated from sample SO5, 23 from sample SO6 and 15 from sample SO7) using serial dilutions and PCA.
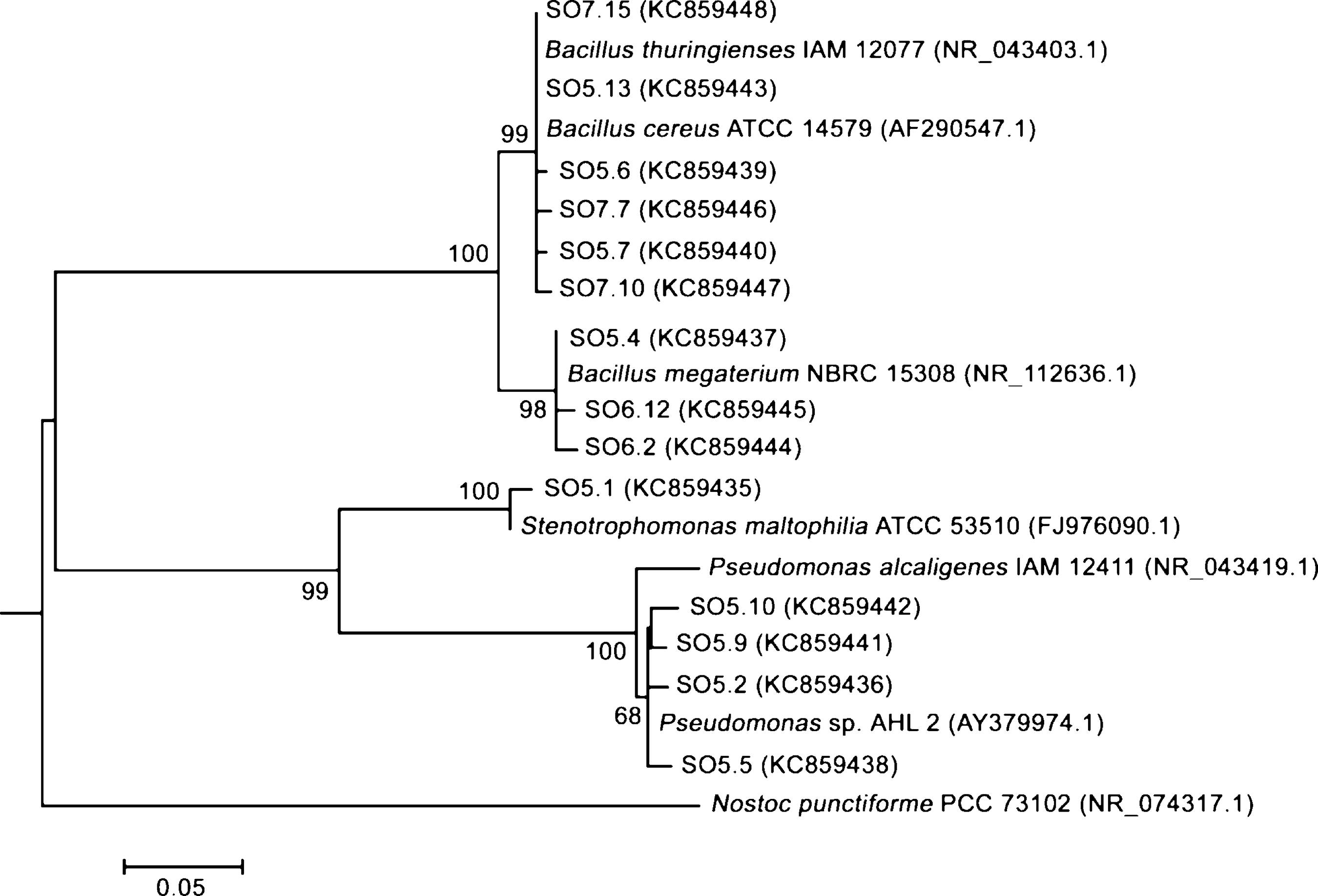
Fourteen strains with outstanding hydrolytic or oxidative activity (above 99% and 10%, respectively) were identified after partial sequencing of the 16S rRNA gene (Table 3, Fig. 1). Nine strains of Bacillus were identified as B. thuringiensis, B. megaterium, B. cereus and Bacillus sp. Other strains recovered were Stenotrophomonas maltophilia and Pseudomonas sp.
Identification of heterotrophic bacterial strains isolated from a neutral copper mine drainage by sequencing of the 16S rRNA gene.
Phylogenetic analysis based on partial 16 rRNA gene sequences of the identified bacterial strains and related microorganisms. The evolutionary history was inferred using the Maximum Likelihood method based on the Kimura 2-parameter model. The numbers at nodes indicate bootstrap percent values from the Maximum Likelihood analysis, based on 1000 resampled data sets. Only significant bootstrap values greater than 65% are shown near each node. Bars represent sequence divergence, and the sequence accession numbers are indicated in parenthesis. Nostoc punctiforme was used as the outgroup.
Discussion
The 56 heterotrophic bacterial strains isolated from this neutral copper mine drainage were evaluated by fluorescence-based HTS enzymatic screening and displayed distinct enzymatic activity toward the selected probes. Esterases and monooxygenases were found in 75% and 20% of strains, respectively. Hydrolases and oxidoreductases are frequently employed in industrial biocatalyzed processes,2222 Kirk O, Borchert TV, Fuglsang CC. Industrial enzyme applications. Curr Opin Biotechnol. 2002;13:345-351. and the search for novel sources of these enzymes is essential for the development of new processes. The HTS results, therefore, indicate that the restricted environment of a copper mine drainage is a promising source of new biocatalysts.
The best oxidation results were obtained using strains isolated from sample SO5 (Table 1), which were further evaluated by multibioreaction assays assessing sulfide biooxidation.
Reduced organic sulfur compounds, such as sulfides, were chosen as targets because of their presence in industrial aqueous wastes. These organic sulfides are often bad smelling and environmentally damaging. They are classified as air and water pollutants, and their bioremediation is an important ecological issue.2323 Habibi MH, Vosooghian H. Photocatalytic degradation of some organic sulfides as environmental pollutants using titanium dioxide suspension. J Photochem Photobiol A: Chem. 2005;174:45-52.
Sulfur atom oxidation is the first bioremediation step, producing sulfoxides (monooxygenation) or sulfones (dioxygenation), which are less volatile and have a less unpleasant odor. Furthermore, sulfoxides are industrially relevant compounds that have potential as drugs and are usually applied as precursors of aromas and flavors and used in asymmetric organic syntheses.2424 Bentley R. Role of sulfur chirality in the chemical processes of biology. Chem Soc Rev. 2005;34:609-624.
Compound 18 is usually employed to detect microbial sulfide oxidation and, in this study, was oxidized by all screened bacteria. Sulfide 19 is a common, fruity odor constituent and was oxidized to sulfoxide 19a by bacteria SO5-4, SO5-9 and SO6-2, which resulted in a change from its original odor to a new aroma valued by fragrance industries.2525 Bentley R. The nose as a stereochemist. Enantiomers and odor. Chem Rev. 2006;106:4099-40112. Furthermore, oxidation of rac-cis-19 provided only the rac-trans-trans-19a (Scheme 3), with over 99% of diastereomeric excess toward rac-cis-cis- 19a, which is chemically expected as both enantiomers have all substituents at equatorial positions. Oxidation of sulfide 20 was also accomplished by SO5-4, SO5-9 and SO6-2, providing sulfoxide rac-20a instead of rac-20b (Scheme 2) because the lone pair of the external sulfur atom of 20 is not part of the aromatic benzothiazole system.
These results are promising and indicate that biooxidation/bioremediation of organic sulfides can be performed by bacteria isolated from the restricted environment of neutral copper mine drainages.
Most isolated bacteria with outstanding enzymatic activities were identified as belonging to the Bacillus genus, which is consistent with the widespread distribution of Bacillus spp., including in mining environments.2626 Selenska-Pobell S, Flemming K, Tzvetkova T, Raff J, Schnorpfeil M, Geibler A. Bacterial communities in uranium mining waste piles and their interaction with heavy metals. In: Merkel BJ, Planer-Friedrich B, Wolkersdorfer, eds. Uranium in the Aquatic Environment. Berlin: Springer-Verlag; 2002.
27 Freitas DB, Reis M, Lima-Bittencourt CI, et al. Genotypic and phenotypic diversity of Bacillus spp. isolated from steel plant waste. BMC Res Notes. 2008;1:92.-2828 Dhanjal S, Cameotra SS. Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb Cell Fact. 2010;9:52-62.Bacillus species have displayed broad catabolic abilities and are the major industrial hydrolase producers, responsible for approximately 50% of the total enzyme market production.2929 Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial production. Can J Microbiol. 2004;50:1-17.
The other genera recovered were Stenotrophomonas and Pseudomonas, identified, respectively, as S. maltophilia and Pseudomonas sp. Both Stenotrophomonas and Pseudomonas have been previously isolated from heavy metal-contaminated soils and other environmental mining samples.3030 Freitas DB, Lima-Bittencourt CI, Reis MP, et al. Molecular characterization of early colonizer bacteria from wastes in a steel plant. Lett Appl Microbiol. 2008;47:241-249.
31 Morel MA, Ubalde MC, Oliveira-Bravo S, Callejas C, Gill PR, Castro-Sowinski SC. Cellular and biochemical response to Cr (VI) in Stenotrophomonas sp. FEMS Microbiol Lett. 2009;291:162-168.
32 Nangia Y, Wangoo N, Goyal N, Shekhawat G, Suri CR. A novel bacterial isolate Stenotrophomonas maltophilia as living factory for synthesis of gold nanoparticles. Microb Cell Fact. 2009;8:39-46.-3333 Choudhary S, Sar P. Uranium biomineralization by a metal resistant Pseudomonas aeruginosa strain isolated from contaminated mine waste. J Hazard Mater. 2011;186:336-343. Bacterial species from the Stenotrophomonas and Pseudomonas genera are well known for their metabolic diversity, multidrug resistance and ability to degrade a wide range of compounds, including pollutants.3434 Samanta SK, Singh OV, Jain RK. Polycyclic 420 aromatic hydrocarbons: environmental pollution and bioremediation. Trends Biotechnol. 2002;20:243-248.
35 Puchałka J, Oberhardt MA, Godinho M, et al. Genome-scale reconstruction and analysis of the Pseudomonas putida KT2440 metabolic network facilitates applications in biotechnology. PLoS Comput Biol. 2008;4:e1000210.-3636 Ryan RP, Monchy S, Cardinale M, et al. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol. 2009;7:514-525.
In this work, the Bacillus strains (SO5-6, SO5-13, SO6-12, SO7-7, SO7-10 and SO7-15) displayed good hydrolase activity (Tables 1 and 3). These strains, as well as the strains identified as Pseudomonas, presented a moderate level of activity for monooxygenases (Tables 1 and 3).
The best biocatalytic performance for sulfide oxidation was accomplished by Pseudomonas sp. (SO5-9), followed by two Bacillus megaterium strains (SO5-4 and SO6-2) (Tables 2 and 3). Mahmood et al. (2009)3737 Mahmood Q, Zheng P, Hu B, et al. Isolation and characterization of Pseudomonas stutzeri QZ1 from an anoxic sulfide-oxidizing bioreactor. Anaerobe. 2009;15:108-115. isolated a strain of Pseudomonas stutzeri that was characterized as a chemolithoautotrophic sulfide-oxidizing and nitrite-reducing strain from the sludge of an anoxic sulfide-oxidizing reactor. Lee et al. (2012)3838 Lee CY, Ho KL, Lee DJ, Su A, Chang JS. Electricity harvest from nitrate/sulfide containing wastewaters using microbial fuel cell with autotrophic denitrifier, Pseudomonas sp. C27. Int J Hydrogen Energy. 2012;37:15827-15832. identified a Pseudomonas sp. strain isolated from a sludge reactor that could successfully convert sulfide to elementary sulfur and thus demonstrated that sulfide oxidation could be applied to electricity generation from sulfide-containing industrial effluent. Therefore, our results indicate the mine strains' potential application as biocatalysts in organic syntheses and bioremediation, corroborating previous data that demonstrated the use of Pseudomonas and Bacillus in industry.
In conclusion, this study provides a novel overview of the heterotrophic bacteria from a copper mine drainage, which might be useful for future investigations of industrial processes, enzyme expression and bioremediation of contaminated environments. Furthermore, the obtained results will support further monitoring of the microbiological processes occurring in this type of restricted environment.
Acknowledgements
We thank Pablo dos Santos Pina for the samples from the Sossego mine. We also thank Companhia Ambiental do Estado de São Paulo (CETESB) for the isolation of the bacterial strains, especially Ana Paula G. Christ and Nancy de Castro Stoppe.
References
-
1ICSG – The International Copper Study Group. The World Copper Factbook 2013 Lisbon: ICSG; 2013.
-
2Blamey A. Brazil's Vale targets 289,000 mt nickel, 405,000 mtcopper in 2014. Platts; 2013. Available at: http://www.platts.com/latest-news/metals/london/brazils-vale-targets-289000-mt-nickel-405000-26507146 Accessed 28.01.14.
» http://www.platts.com/latest-news/metals/london/brazils-vale-targets-289000-mt-nickel-405000-26507146 -
3Johnson DB. Chemical and microbiological characteristics of mineral spoils and drainage waters at abandoned coal and metal mines. Water Air Soil Pollut 2003;3:47-66.
-
4Falteisek L, Čepička I. Microbiology of diverse acidic and non-acidic microhabitats within a sulfidic ore mine. Extremophiles 2012;16:911-922.
-
5Watling HR, Watkin ELJ, Ralph DE. The resilience and versatility of acidophiles that contribute to the bio-assisted extraction of metals from mineral sulphides. Environ Technol 2010;31:915-933.
-
6Hallberg KB. New perspectives in acid mine drainage microbiology. Hydrometallurgy 2010;104:448-453.
-
7Mantovani SM, Oliveira LG, Marsaioli AJ. Whole cell quick-E for epoxide hydrolase screening using fluorescent probes. J Mol Catal B: Enzym 2008;52-53:173-177.
-
8Cruz GF, Angolini CFF, Oliveira LG, et al. Searching for monooxygenases and hydrolases in bacteria from an extreme environment. Appl Microbiol Biotechnol 2010;87:319-329.
-
9Silva CP. Enzymatic Potential of the Human Skin Microbiota and its Effects on Fragrance Ingredients University of Campinas;2012 [Ph.D. thesis].
-
10Gonçalves RAC, Porto ALM, Pinheiro L, Cagnon JR, Manfio GP, Marsaioli AJ. Multibioreaction methodology for Baeyer-Villiger monooxygenase monitoring. Food Technol Biotechnol 2004;42:355-361.
-
11Sicard R, Chen LS, Marsaioli AJ, Reymond J-L. A fluorescence-based assay for Baeyer-Villiger monooxygenases, hydroxylases and lactonases. Adv Synth Catal 2005;245:1041-1050.
-
12Bicalho B, Chen LS, Grognux J, Reymond JL, Marsaioli AJ. Studies on whole cell fluorescence-based screening for epoxide hydrolases and Baeyer-Villiger monooxygenases. J Braz Chem Soc 2004;15:911-916.
-
13Porto ALM, Cassiola F, Dias SLP, et al. Aspergillus terreus CCT 3320 immobilized on chrysotile or cellulose/TiO2 for sulfide oxidation. J Mol Catal B: Enzym 2002;19-20:327-334.
-
14Eaton AD, Franson MH. Standard Methods for the Examination ofWater and Wastewater 21st ed. Washington, DC: AmericanPublic Health Association; 2005.
-
15Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989.
-
16Sørensen SPL. Enzymstudien: II. Mitteilung. Über die Messung und die Bedeutung der Wasserstoffionenkoncentration bei enzymatischen Prozessen. Biochem Z 1909;21:131-304.
-
17Miqueletto PB, Andreote FD, Dias ACF, Ferreira JC, Dos Santos Neto EV, Oliveira VM. Cultivation-independent methods applied to the microbial prospection of oil and gas in soil from a sedimentary basin in Brazil. AMB Express 2011;1:1-16.
-
18Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 1994;22:4673-4680.
» https://doi.org/10.1093/nar/22.22.4673 -
19Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol 2013;30:2725-2729.
-
20Kimura M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 1980;16:111-120.
-
21Yang Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: approximate methods. J Mol Evol 1994;39:306-314.
-
22Kirk O, Borchert TV, Fuglsang CC. Industrial enzyme applications. Curr Opin Biotechnol 2002;13:345-351.
-
23Habibi MH, Vosooghian H. Photocatalytic degradation of some organic sulfides as environmental pollutants using titanium dioxide suspension. J Photochem Photobiol A: Chem 2005;174:45-52.
-
24Bentley R. Role of sulfur chirality in the chemical processes of biology. Chem Soc Rev 2005;34:609-624.
-
25Bentley R. The nose as a stereochemist. Enantiomers and odor. Chem Rev 2006;106:4099-40112.
-
26Selenska-Pobell S, Flemming K, Tzvetkova T, Raff J, Schnorpfeil M, Geibler A. Bacterial communities in uranium mining waste piles and their interaction with heavy metals. In: Merkel BJ, Planer-Friedrich B, Wolkersdorfer, eds. Uranium in the Aquatic Environment Berlin: Springer-Verlag; 2002.
-
27Freitas DB, Reis M, Lima-Bittencourt CI, et al. Genotypic and phenotypic diversity of Bacillus spp. isolated from steel plant waste. BMC Res Notes 2008;1:92.
-
28Dhanjal S, Cameotra SS. Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb Cell Fact 2010;9:52-62.
-
29Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial production. Can J Microbiol 2004;50:1-17.
-
30Freitas DB, Lima-Bittencourt CI, Reis MP, et al. Molecular characterization of early colonizer bacteria from wastes in a steel plant. Lett Appl Microbiol 2008;47:241-249.
-
31Morel MA, Ubalde MC, Oliveira-Bravo S, Callejas C, Gill PR, Castro-Sowinski SC. Cellular and biochemical response to Cr (VI) in Stenotrophomonas sp. FEMS Microbiol Lett 2009;291:162-168.
-
32Nangia Y, Wangoo N, Goyal N, Shekhawat G, Suri CR. A novel bacterial isolate Stenotrophomonas maltophilia as living factory for synthesis of gold nanoparticles. Microb Cell Fact 2009;8:39-46.
-
33Choudhary S, Sar P. Uranium biomineralization by a metal resistant Pseudomonas aeruginosa strain isolated from contaminated mine waste. J Hazard Mater 2011;186:336-343.
-
34Samanta SK, Singh OV, Jain RK. Polycyclic 420 aromatic hydrocarbons: environmental pollution and bioremediation. Trends Biotechnol 2002;20:243-248.
-
35Puchałka J, Oberhardt MA, Godinho M, et al. Genome-scale reconstruction and analysis of the Pseudomonas putida KT2440 metabolic network facilitates applications in biotechnology. PLoS Comput Biol 2008;4:e1000210.
-
36Ryan RP, Monchy S, Cardinale M, et al. The versatility and adaptation of bacteria from the genus Stenotrophomonas Nat Rev Microbiol 2009;7:514-525.
-
37Mahmood Q, Zheng P, Hu B, et al. Isolation and characterization of Pseudomonas stutzeri QZ1 from an anoxic sulfide-oxidizing bioreactor. Anaerobe 2009;15:108-115.
-
38Lee CY, Ho KL, Lee DJ, Su A, Chang JS. Electricity harvest from nitrate/sulfide containing wastewaters using microbial fuel cell with autotrophic denitrifier, Pseudomonas sp. C27. Int J Hydrogen Energy 2012;37:15827-15832.
Publication Dates
-
Publication in this collection
Oct-Dec 2016
History
-
Received
25 Mar 2015 -
Accepted
22 Feb 2016