ABSTRACT
L-asparaginase (EC 3.5.1.1) is an enzyme that catalysis mainly the asparagine hydrolysis in L-aspartic acid and ammonium. This enzyme is presented in different organisms, such as microorganisms, vegetal, and some animals, including certain rodent's serum, but not unveiled in humans. It can be used as important chemotherapeutic agent for the treatment of a variety of lymphoproliferative disorders and lymphomas (particularly acute lymphoblastic leukemia (ALL) and Hodgkin's lymphoma), and has been a pivotal agent in chemotherapy protocols from around 30 years. Also, other important application is in food industry, by using the properties of this enzyme to reduce acrylamide levels in commercial fried foods, maintaining their characteristics (color, flavor, texture, security, etc.) Actually, L-asparaginase catalyzes the hydrolysis of L-asparagine, not allowing the reaction of reducing sugars with this aminoacid for the generation of acrylamide. Currently, production of L-asparaginase is mainly based in biotechnological production by using some bacteria. However, industrial production also needs research work aiming to obtain better production yields, as well as novel process by applying different microorganisms to increase the range of applications of the produced enzyme. Within this context, this mini-review presents L-asparaginase applications, production by different microorganisms and some limitations, current investigations, as well as some challenges to be achieved for profitable industrial production.
Keywords:
L-asparaginase; Microbial production; Industrial production; Pharmaceutical application; Acrylamide
Introduction
L-asparaginase aminohydrolase (L-asparaginase, EC 3.5.1.1), has gained attention in recent years due to its important applications, as its use in pharmaceutical industry as an alternative for treatment of different cancers such as acute lymphoblastic leukemia, malignant diseases of the lymphoid system and Hodgkin's lymphomas.11 Appel IM, van Kessel-Bakvis C, Stigter R, Pieters R. Influence of two different regimens of concomitant treatment with asparaginase and dexamethasone on hemostasis in childhood acute lymphoblastic leukemia. Leukemia. 2007;21(11):2377-2380, http://dx.doi.org/10.1038/sj.leu.2404793.
http://dx.doi.org/10.1038/sj.leu.2404793...
Also, this enzyme is used in food industry to prevent the acrylamide formation when foods are processed in high temperatures.22 Mohan Kumar NS, Shimray CA, Indrani D, Manonmani HK. Reduction of acrylamide formation in sweet bread with L-asparaginase treatment. Food Bioprocess Technol. 2013;7(3):741-748, http://dx.doi.org/10.1007/s11947-013-1108-6
http://dx.doi.org/10.1007/s11947-013-110...
This use is important because acrylamide is a neurotoxin classified as potentially carcinogenic to humans.33 Medeiros Vinci R, Mestdagh F, De Meulenaer B. Acrylamide formation in fried potato products - present and future, a critical review on mitigation strategies. Food Chem. 2012;133(4):1138-1154, http://dx.doi.org/10.1016/j.foodchem.2011.08.001.
http://dx.doi.org/10.1016/j.foodchem.201...
Industrial L-asparaginase production presents some challenges, such as the search for new microorganisms able to produce it with less adverse effects. Nowadays, industrial production is carried out using bacteria such as Escherichia coli and Erwinia chrysanthemi.44 Keating MJ, Holmes R, Lerner S, Ho DH. L-asparaginase and PEG asparaginase - past, present, and future. Leuk Lymphoma. 1993;10(suppl):153-157, http://dx.doi.org/10.3109/10428199309149129.
http://dx.doi.org/10.3109/10428199309149...
However, the enzyme obtained from prokaryotic microorganisms usually presents some problems such as hypersensitivity and immune inactivation.55 Narta UK, Kanwar SS, Azmi W. Pharmacological and clinical evaluation of L-asparaginase in the treatment of leukemia. Crit Rev Oncol Hematol. 2007;61(3):208-221, http://dx.doi.org/10.1016/j.critrevonc.2006.07.009.
http://dx.doi.org/10.1016/j.critrevonc.2...
Within this context, eukaryotic microorganisms such as filamentous fungi66 Dange V, Peshwe S. Purification and biochemical characterization of L-asparaginase from Aspergillus niger and evaluation of its antineoplastic activity. Int J Sci Res. 2015;4(2):564-569. and yeasts77 Soler MF, Pedreira VA, Longo FF, et al. BB 25. Seleção de leveduras produtoras de L-asparaginase em meios sólido e líquido: uma comparação de diferentes metodologias de screening. J Basic Appl Pharm Sci. 2015;36(1). have been investigated for this enzymes production, due to better compatibility with the human system.
Currently, new studies have been carried out aiming to enhance production process and establish new ways for enzyme synthesis. Thus, some of these aspects are discussed, besides some generalities regarding L-asparaginase applications in pharmaceutical and food industries.
Reactions and mechanism
The hydrolysis process occurs in two steps through an intermediate: beta-acyl-enzyme (Fig. 1). In the first process step, the nucleophilic residue of the enzyme is activated by a strong base and attacks the amide carbon atom of L-asparagine (substrate), generating a product beta-acyl-enzyme intermediate. The second reaction step is an attack on the ester carbon made by a nucleophile activated by a water molecule.88 Verma N, Kumar K, Kaur G, Anand S. L-asparaginase: a promising chemotherapeutic agent. Crit Rev Biotechnol. 2007;27(1):45-62, http://dx.doi.org/10.1080/07388550601173926.
http://dx.doi.org/10.1080/07388550601173...
General mechanism of L-asparaginase reaction catalyzed. Dashed arrow shows nucleophilic attack.
This mechanism is comparable to serine-proteases classic mechanism, whose activities depends of an amino acid group, classified as catalytic triads. This catalytic triads is composed by one nucleophilic amino acid, serine (Ser), one base, histidine (His) and one amino acid with acid characteristic, aspartic acid (Asp), all connected by hydrogen bonds.88 Verma N, Kumar K, Kaur G, Anand S. L-asparaginase: a promising chemotherapeutic agent. Crit Rev Biotechnol. 2007;27(1):45-62, http://dx.doi.org/10.1080/07388550601173926.
http://dx.doi.org/10.1080/07388550601173...
L-asparaginase has also capacity to catalase other reactions. For example, L-asparaginase produced by Serratia marcescens is able to hydrolase 5% of L-glutamine when compared with L-asparaginase hydrolysis. The same effect occurs to L-asparaginase produced by Escherichia coli and Erwinia chrysanthemi. Other microorganisms, such as Pseudomonas sp. and Acinetobacter glutaminasificans, synthetize L-asparaginase with equal asparaginase and glutaminase activity. In some cases, L-asparaginase starts the L-glutamine hydrolysis only after complete conversion of L-asparaginase in aspartic acid. Actually, L-glutamine is a competitive inhibitor of L-asparagine hydrolysis.1212 Van den Berg H. Asparaginase revisited. Leuk Lymphoma. 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127.
http://dx.doi.org/10.3109/10428199309149...
L-glutamine and L-asparagine hydrolysis are similar due to the structural similarity from both amino acids. Therefore, the largest part of microbial L-asparagine presents cross glutaminase activity, with some exceptions such as L-asparaginase produced by Wolinella succinogenes, which do not present L-glutaminase activity.1212 Van den Berg H. Asparaginase revisited. Leuk Lymphoma. 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127.
http://dx.doi.org/10.3109/10428199309149...
Finally, L-asparaginase is also able to hydrolyze β-aspartyl peptide amide, however reaction yield is considerably low.1313 Noronkoski T, Stoineva IB, Ivanov IP, Petkov DD, Mononen I. Glycosylasparaginase-catalyzed synthesis and hydrolysis of -aspartyl peptides. J Biol Chem. 1998;273(41):26295-26297, http://dx.doi.org/10.1074/jbc.273.41.26295.
http://dx.doi.org/10.1074/jbc.273.41.262...
L-asparaginase applications
Pharmaceutical industry: antineoplastic action
The L-asparagine enzymatic hydrolysis in L-aspartate and ammonium was observed in a first time by Lang (1904),1414 Lang S. Uber desamidierung im Tierkorper. Beitr Chem Physiol Pathol. 1904;5:321-345. that detected L-asparaginase activity in bovine's tissues. Results of this researcher were confirmed by Furth and Friedmann (1910),1515 Furth O, Friedmann M. Uber die Verbreitung asparaginspaltender Organfermente. Biochemistry. 1910;26:435-440. that detected L-asparagine hydrolase in horse and pig organs, observing the same amount of L-asparaginase activity in both animals. Also, Clementi (1922)1616 Clementi A. La desamidation enzymatique de l’asparagine chez les differentes especes animals et la signification physiologique de sa presence dans l’organisme. Arch Intern Physiol. 1922;19:369-398. related that L-asparaginase in guinea pig serum, although antitumor activity of the enzyme was identified only some years later. In addition, Mashburn and Wriston (1964)1717 Mashburn LT, Wriston JC. Tumor inhibitory effect of L-asparaginase from Eschericha coli. Arch Biochem Biophys. 1964;105:450-452. demonstrated that L-asparaginase of E. coli had inhibitory capacity of tumors in rats. However, the large interest in enzyme started when Broome (1965)1818 Broome JD. Antilymphoma activity of L-asparaginase in vivo: clearance rates of enzyme preparations from guinea pig serum and yeast in relation to their effect on tumor growth. J Natl Cancer Inst. 1965;35(6):967-974. found that the regression lymphosarcoma transplants in rats treated with guinea pig serum was due to nutritional dependence on malignant cells of exogenous L-asparagine.
Considering its properties, L-asparaginase has been an important chemotherapeutic agent used for treatment of lymphoproliferative and lymphoma diseases. Particularly, it presents large importance in chemotherapeutic protocols for acute lymphoblastic leukemia (ALL) and Hodgkin's lymphomas.1919 Schrappe M, Reiter A, Ludwig WD, et al. Improved outcome in childhood acute lymphoblastic leukemia despite reduced use of anthracyclines and cranial radiotherapy: results of trial ALL-BFM 90. German-Austrian-Swiss ALL-BFM Study Group. Blood. 2000;95(11):3310-3322. http://www.ncbi.nlm.nih.gov/pubmed/10828010 Accessed 09.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/10828...
Cancer cells, mainly lymphatic cells, require high amount of asparagine for fast and malignant growth. In this way, cancer cells requires the amino acid from diet (blood serum) as well as amino acids produced by themselves. However, leukemic lymphoblasts and some others tumor cells do not have or present low quantity of L-asparagine synthetase used for L-asparagine syntheses. Thus, these malignant cells are dependent of asparagine from blood serum for their proliferation and survival.2020 Kiriyama Y, Kubota M, Takimoto T, et al. Biochemical characterization of U937 cells resistant to L-asparaginase: the role of asparagine synthetase. Leukemia. 1989;3(4):294-297. http://www.ncbi.nlm.nih.gov/pubmed/2564453 Accessed 17.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/25644...
,2121 Stams WAG, den Boer ML, Beverloo HB, et al. Sensitivity to L-asparaginase is not associated with expression levels of asparagine synthetase in t(12;21)+ pediatric ALL. Blood. 2003;101(7):2743-2747, http://dx.doi.org/10.1182/blood-2002-08-2446.
http://dx.doi.org/10.1182/blood-2002-08-...
L-asparaginase hydrolyzes asparagine from blood serum, leading tumor cells to death by lacking of an essential factor for protein synthetases (p53-dependent apoptosis). However, healthy cells are not affected, because they are able to produce asparagine using L-asparagine synthetase present in enough quantities (Fig. 2). Considering these concepts, Fig. 2 schematically shows the antineoplastic action of L-asparaginase.
Food industry: acrylamide formation
Acrylamide (C3H5NO) is also known as 2-propenamide, acrylic amide, ethylene carboxamide, propenamide, propanoic acid amide, monomer of acrylamide or acrylic acid amide, presenting 71.08 g/mol of molecular mass.2222 Friedman M. Chemistry, biochemistry, and safety of acrylamide. A review. J Agric Food Chem. 2003;51(16):4504-4526, http://dx.doi.org/10.1021/jf030204+.
http://dx.doi.org/10.1021/jf030204+...
Several studies show that L-asparagine is the main amino acid responsible for acrylamide production in fried and baked foods when reducing sugars are condensed with a carbonyl source. This phenomenon does not occur in boiled food.2323 Tareke E, Rydberg P, Karlsson P, Eriksson S, Törnqvist M. Acrylamide: a cooking carcinogen? Chem Res Toxicol. 2000;13(6):517-522. http://www.ncbi.nlm.nih.gov/pubmed/10858325 Accessed 18.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/10858...
Acrylamide formation has been quite studied in the last years. Zyzak et al. (2003)2424 Zyzak DV, Sanders RA, Stojanovic M, et al. Acrylamide formation mechanism in heated foods. J Agric Food Chem. 2003;51(16):4782-4787, http://dx.doi.org/10.1021/jf034180i.
http://dx.doi.org/10.1021/jf034180i...
detected that the amide chain present in the acrylamide structure is provided from L-asparagine. Reagents (L-asparagine or reducing sugars) reduction or removal is one of the evaluated strategies for decreasing acrylamide quantity in foods. For L-asparagine reduction, several options have been investigated, such as: selection of vegetal species with lower level of L-asparagine in their composition; deletion of important enzymes for L-asparagine biosynthesis control by suppression of specific genes; acid hydrolysis of L-asparagine leading the formation of aspartic acid and ammonia; and acetylation process of L-asparagine to form N-acetyl-L-asparagine, preventing the formation of acrylamide from intermediate N-glycosides.2222 Friedman M. Chemistry, biochemistry, and safety of acrylamide. A review. J Agric Food Chem. 2003;51(16):4504-4526, http://dx.doi.org/10.1021/jf030204+.
http://dx.doi.org/10.1021/jf030204+...
In the study of Zyzak et al. (2003),2424 Zyzak DV, Sanders RA, Stojanovic M, et al. Acrylamide formation mechanism in heated foods. J Agric Food Chem. 2003;51(16):4782-4787, http://dx.doi.org/10.1021/jf034180i.
http://dx.doi.org/10.1021/jf034180i...
authors confirmed that the use of L-asparaginase enzyme before frying or baking food process could reduce more than 99% acrylamide level in the processed final product. This is because the enzyme reduces more than 88% of the L-asparagine concentration from the initial feedstock. In last years, other works have dealt with this application of L-asparaginase, that can decrease the negative effects of acrylamide containing foods without impair their characteristics.33 Medeiros Vinci R, Mestdagh F, De Meulenaer B. Acrylamide formation in fried potato products - present and future, a critical review on mitigation strategies. Food Chem. 2012;133(4):1138-1154, http://dx.doi.org/10.1016/j.foodchem.2011.08.001.
http://dx.doi.org/10.1016/j.foodchem.201...
,2525 Zhang S, Xie Y, Zhang C, et al. Biochemical characterization of a novel L-asparaginase from Bacillus megaterium H-1 and its application in French fries. Food Res Int. 2015;77:527-533, http://dx.doi.org/10.1016/j.foodres.2015.08.031.
http://dx.doi.org/10.1016/j.foodres.2015...
26 Huang L, Liu Y, Sun Y, Yan Q, Jiang Z. Biochemical characterization of a novel L-asparaginase with low glutaminase activity from Rhizomucor miehei and its application in food safety and leukemia treatment. Appl Environ Microbiol. 2014;80(5):1561-1569, http://dx.doi.org/10.1128/AEM.03523-13.
http://dx.doi.org/10.1128/AEM.03523-13...
27 Anese M, Quarta B, Peloux L, Calligaris S. Effect of formulation on the capacity of L-asparaginase to minimize acrylamide formation in short dough biscuits. Food Res Int. 2011;44(9):2837-2842, http://dx.doi.org/10.1016/j.foodres.2011.06.025.
http://dx.doi.org/10.1016/j.foodres.2011...
-2828 Pedreschi F, Mariotti S, Granby K, Risum J. Acrylamide reduction in potato chips by using commercial asparaginase in combination with conventional blanching. LWT - Food Sci Technol. 2011;44(6):1473-1476, http://dx.doi.org/10.1016/j.lwt.2011.02.004.
http://dx.doi.org/10.1016/j.lwt.2011.02....
Production by different microorganisms
L-asparaginase is present in mammals, birds, plants, yeast, and a wide range of bacteria.1010 El-Bessoumy AA, Sarhan M, Mansour J. Production, isolation, and purification of L-asparaginase from Pseudomonas aeruginosa 50071 using solid-state fermentation. J Biochem Mol Biol. 2004;37(4):387-393, http://dx.doi.org/10.5483/BMBRep.2004.37.4.387.
http://dx.doi.org/10.5483/BMBRep.2004.37...
,2929 Wriston JC, Yellin TO. L-asparaginase: a review. Adv Enzymol Relat Areas Mol Biol. 1973;39:185-248. http://www.ncbi.nlm.nih.gov/pubmed/4583638 Accessed 23.06.16.
http://www.ncbi.nlm.nih.gov/pubmed/45836...
Although L-asparaginase production is observed in animals, plants,1212 Van den Berg H. Asparaginase revisited. Leuk Lymphoma. 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127.
http://dx.doi.org/10.3109/10428199309149...
,3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
the microorganisms are considered mainly source for L-asparagine synthesis.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
,3131 Deshpande N, Choubey P, Agashe M. Studies on optimization of growth parameters for L-asparaginase production by streptomyces ginsengisoli. Sci World J. 2014;2014:1-6, http://dx.doi.org/10.1155/2014/895167.
http://dx.doi.org/10.1155/2014/895167...
The production of this enzyme is mainly proceeded by submerged fermentation.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
Several researchers have studied the isolation of microbial strains that produce this important enzyme, such as Pseudomonas fluorescens,3232 Degroot N, Lichtenstein N. The action of Pseudomonas fluorescens extracts on asparagine and asparagine derivatives. Biochim Biophys Acta. 1960;40:99-110.Serratia marcescens,3333 Rowley B, Wriston JCJ. Partial purification and antilymphoma activity of Serratia marcescens L-asparaginase. Biochem Biophys Res Commun. 1967;28(2):160-165.Escherichia coli,3434 Kozak M, Jurga S. A comparison between the crystal and solution structures of Escherichia coli asparaginase II. Acta Biochim Pol. 2002;49(2):509-513. http://www.ncbi.nlm.nih.gov/pubmed/12362993 Accessed 17.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/12362...
Erwinia carotovora,3535 Wade HE, Elsworth R, Herbert D, Keppie J, Sargeant K. A new L-asparaginase with antitumour activity? Lancet. 1968;2(7571):776-777.Proteus vulgaris,3636 Tosa T, Sano R, Yamamoto K, Nakamura M, Ando K, Chibatha I. L-asparaginase from Proteus vulgaris. Appl Microbiol. 1972;22:387-392.Saccharomyces cerevisiae, Karnatakensis Streptomyces, Streptomyces venezuelae and several genres of fungi as Aspergillus, Penicillium and Fusarium.3737 Sarquis MI, de M, Oliveira EMM, Santos AS, da Costa GL. Production of L-asparaginase by filamentous fungi. Mem Inst Oswaldo Cruz. 2004;99(5):489-492, http://dx.doi.org/10.1590/S0074-02762004000500005.
http://dx.doi.org/10.1590/S0074-02762004...
Concerning to bacteria, the best producers of L-asparaginase are members of the Enterobacteriaceae family.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
For example, in pharmaceutical industry, this enzyme is produced mainly from bacteria such as Escherichia coli and Erwinia carotova, (also known as Erwinia chrysanthemi), generally used for leukemia and lymphoma treatment.44 Keating MJ, Holmes R, Lerner S, Ho DH. L-asparaginase and PEG asparaginase - past, present, and future. Leuk Lymphoma. 1993;10(suppl):153-157, http://dx.doi.org/10.3109/10428199309149129.
http://dx.doi.org/10.3109/10428199309149...
However, most of these treatments can result in immunological sensitization (hypersensitivity) and immune inactivation in patients that receive bacterial enzymes.55 Narta UK, Kanwar SS, Azmi W. Pharmacological and clinical evaluation of L-asparaginase in the treatment of leukemia. Crit Rev Oncol Hematol. 2007;61(3):208-221, http://dx.doi.org/10.1016/j.critrevonc.2006.07.009.
http://dx.doi.org/10.1016/j.critrevonc.2...
Another issue is that glutaminase activity generated by these enzymes can cause secondary effects such as allergic reaction, nausea, pancreatitis, diabetes and coagulation abnormalities.88 Verma N, Kumar K, Kaur G, Anand S. L-asparaginase: a promising chemotherapeutic agent. Crit Rev Biotechnol. 2007;27(1):45-62, http://dx.doi.org/10.1080/07388550601173926.
http://dx.doi.org/10.1080/07388550601173...
,3838 Duval M. Comparison of Escherichia coli-asparaginase with Erwinia asparaginase in the treatment of childhood lymphoid malignancies: results of a randomized European Organisation for Research and Treatment of Cancer - Children's Leukemia Group phase 3 trial. Blood. 2002;99(8):2734-2739, http://dx.doi.org/10.1182/blood.V99.8.2734.
http://dx.doi.org/10.1182/blood.V99.8.27...
Also, most of asparaginases has low stability and catalytic activity, presenting only active in a narrow pH range.3939 Zuo S, Zhang T, Jiang B, Mu W. Recent research progress on microbial L-asparaginases. Appl Microbiol Biotechnol. 2014;99(3):1069-1079, http://dx.doi.org/10.1007/s00253-014-6271-9.
http://dx.doi.org/10.1007/s00253-014-627...
Currently, L-asparaginases from E. coli and Erwinia chrysanthemi (synonymous of Erwinia carotovora) are the only preparation available for medical use.1212 Van den Berg H. Asparaginase revisited. Leuk Lymphoma. 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127.
http://dx.doi.org/10.3109/10428199309149...
L-asparaginase from E. coli produces two types of enzyme, L-asparaginase I (EC1), found in the cytoplasm and L-asparaginase II (EC2), with periplasmic origin.4040 Casale T Del, Sollitti P, Chesney RH. Cytoplasmic L-asparaginase: isolation of a defective strain and mapping of ansA. J Bacteriol. 1983;154(1):513-515. However, only the second one has anti-cancer activity.4141 Whitecar JP, Bodey GP, Harris JE, Freireich EJ. L-asparaginase. N Engl J Med. 1970;282(13):732-734, http://dx.doi.org/10.1056/NEJM197003262821307.
http://dx.doi.org/10.1056/NEJM1970032628...
Some studies describes EC1 as a constitutive enzyme and EC2 as secreted only as a response to exposure to low concentrations of nitrogen.88 Verma N, Kumar K, Kaur G, Anand S. L-asparaginase: a promising chemotherapeutic agent. Crit Rev Biotechnol. 2007;27(1):45-62, http://dx.doi.org/10.1080/07388550601173926.
http://dx.doi.org/10.1080/07388550601173...
EC2 has an estimated molecular weight of 141 kDa and its kM is about 12.5 µM, meaning a high affinity for substrate.4242 Kumar D, Sobha K. L-asparaginase from Microbes: a comprehensive review. Adv Biores. 2012;3(December):137-157. http://soeagra.com/abr/abrdec 2012/22.pdf.
http://soeagra.com/abr/abrdec 2012/22.pd...
Its half-life is around 1.24 ± 0.17 days and its optimum pH and temperature are 7-8 and 37 °C, respectively.1212 Van den Berg H. Asparaginase revisited. Leuk Lymphoma. 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127.
http://dx.doi.org/10.3109/10428199309149...
,4242 Kumar D, Sobha K. L-asparaginase from Microbes: a comprehensive review. Adv Biores. 2012;3(December):137-157. http://soeagra.com/abr/abrdec 2012/22.pdf.
http://soeagra.com/abr/abrdec 2012/22.pd...
As an alternative for treatment of patients allergic to L-asparaginase from E. coli, L-asparaginase from E. chrysanthemi (ErA) is used. It has half-life of 0.6 ± 0.13 days, a kM of 18 µM, molecular mass about 140 and 150 kDa, optimal pH 8 and 50 °C as optimal temperature.4343 Müller HJ, Boos J. Use of L-asparaginase in childhood ALL. Crit Rev Oncol Hematol. 1998;28(2):97-113. http://www.ncbi.nlm.nih.gov/pubmed/9768345 Accessed 18.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/97683...
,4444 Kumar S, Dasu VV, Pakshirajan K. Localization and production of novel L-asparaginase from Pectobacterium carotovorum MTCC 1428. Process Biochem. 2010;45(2):223-229, http://dx.doi.org/10.1016/j.procbio.2009.09.011.
http://dx.doi.org/10.1016/j.procbio.2009...
The difference between its kM and that one from E. coli's L-aspraginase is because glutaminase activity of ErA is higher.4242 Kumar D, Sobha K. L-asparaginase from Microbes: a comprehensive review. Adv Biores. 2012;3(December):137-157. http://soeagra.com/abr/abrdec 2012/22.pdf.
http://soeagra.com/abr/abrdec 2012/22.pd...
,4545 Aghaiypour K, Wlodawer A, Lubkowski J. Structural basis for the activity and substrate specificity of Erwinia chrysanthemi L-asparaginase. Biochemistry. 2001;40(19):5655-5664. http://www.ncbi.nlm.nih.gov/pubmed/11341830 Accessed 17.02.16.
http://www.ncbi.nlm.nih.gov/pubmed/11341...
In recent years, different studies were developed aiming to find this enzyme with improved characteristics compared to L-asparaginase from E. coli, with economically viable production as well as causing minimal collateral effects. Searching from different L-asparagine sources, specifically eukaryotic microorganisms, can lead to enzymes with less adverse effects and different features, which are advantageous for its application.4646 Sreenivasulu V, Jayaveera K, Rao PM. Solid-state fermentation for the production of L-asparaginase by Aspergillus Sp. Res J Pharmacogn Phytochem. 2009;1(1):21-25. http://www.indianjournals.com/ijor.aspx?target=ijor:rjpp&volume=1&issue=1&article=006 Accessed 15.02.16.
http://www.indianjournals.com/ijor.aspx?...
In the last years, eukaryotic fungi have been investigated as L-asparaginase source.4747 Mishra A. Production of L-asparaginase, an anticancer agent, from Aspergillus niger using agricultural waste in solid state fermentation. Appl Biochem Biotechnol. 2006;135(1):33-42, http://dx.doi.org/10.1385/ABAB:135:1:33.
http://dx.doi.org/10.1385/ABAB:135:1:33...
For L-asparaginase production by fungi, the genera Aspergillus, Penicillium and Fusarium have been studied.66 Dange V, Peshwe S. Purification and biochemical characterization of L-asparaginase from Aspergillus niger and evaluation of its antineoplastic activity. Int J Sci Res. 2015;4(2):564-569. Currently, fungal recombinant L-asparaginase from Aspergillus oryzae and Aspergillus niger has already been used in food industry for reduction of acrylamide formation in some foods.2828 Pedreschi F, Mariotti S, Granby K, Risum J. Acrylamide reduction in potato chips by using commercial asparaginase in combination with conventional blanching. LWT - Food Sci Technol. 2011;44(6):1473-1476, http://dx.doi.org/10.1016/j.lwt.2011.02.004.
http://dx.doi.org/10.1016/j.lwt.2011.02....
Moreover, authors have reported positive results by using endophytic fungi of the genus Colletotrichum, Eupenicillium, Talaromyces.4848 Theantana T, Hyde KD, Lumyong S. Asparaginase production by endophytic fungi from Thai medicinal plants: cytoxicity properties sources of endophytic fungi isolation of endophytic fungi identification of endophytic fungi Identification was based on colony and hyphal. Int J Integr Biol. 2009;7(1):1-8.,4949 Theantana T, Hyde KD, Lumyong S, Mai C. Aparaginase production by endophytic fungi isolated from some Thai medicinal plants. KMITL Sci Tech J. 2007;7:13-18. Also, positive asparaginolytic activity were also shown by researchers that used fungi isolated from marine environments, endophytes seaweed, of genera Alternaria, Chaetomium, Cladosporium, Colletotrichum, Curvularia, Nigrospora, Paecilomyces, Phaeotrichoconis, Phoma and Pithomyces.5050 Thirunavukkarasu N, Suryanarayanan TS, Murali TS, Ravishankar JP, Gummadi SN. L-asparaginase from marine derived fungal endophytes of seaweeds. Mycosphere. 2011;2(2):147-155, http://dx.doi.org/10.1111/j.1365-313X.2009.03887.x.zation.
http://dx.doi.org/10.1111/j.1365-313X.20...
Within this context, Table 1 present some works related to L-asparaginase production by bacteria and fungi.
Also, several studies have shown that Aspergillus genus is available to produce significant amounts of L-asparaginase. For example, Sarquis et al. (2004)3737 Sarquis MI, de M, Oliveira EMM, Santos AS, da Costa GL. Production of L-asparaginase by filamentous fungi. Mem Inst Oswaldo Cruz. 2004;99(5):489-492, http://dx.doi.org/10.1590/S0074-02762004000500005.
http://dx.doi.org/10.1590/S0074-02762004...
presented filamentous fungi like Aspergillus tamarii and Aspergillus terreus as producers of L-asparaginase by submerged fermentation, resulting in 38 U/L and 58.8 U/L, respectively. Authors concluded that enzyme production was regulated by the nitrogen source. Moreover, Balasubramanian et al. (2012),5151 Balasubramanian K, Ambikapathy V, Panneerselvam A. Production, isolation and purification of L-asparaginase from Aspergillus terreus using submerged fermentation. Int J Adv Pharm Res. 2012;3(2):778-783. in a screening study of L-asparaginase producers, reported that Aspergillus terreus was able to produce 9.3 U/mL of enzyme. In other study of culture conditions optimization (temperature 35 °C, initial pH 6.3, inoculum size 1% (v/v), agitation rate 140 rpm, and incubation time 58.5 h), Gurunathan and Sahadevan (2012)5252 Gurunathan B, Sahadevan R. Optimization of culture conditions and bench-scale production of L-asparaginase by submerged fermentation of Aspergillus terreus MTCC 1782. J Microbiol Biotechnol. 2012;22(7):923-929, http://dx.doi.org/10.4014/jmb.1112.12002.
http://dx.doi.org/10.4014/jmb.1112.12002...
reported L-asparaginase production of Aspergillus terreus by submerged fermentation, reaching production of 44.38 U/mL. In another optimization project, but by using Aspergillus niger, Anjum Zia et al. (2013)5353 Anjum Zia MAZ, Bashir R, Ahmed I, Iftikhar T. Production of L-asparaginase from Aspergillus Niger using agro wastes by-products in submerged fermentation process. J Teknol. 2013;62(2), http://dx.doi.org/10.11113/jt.v62.1879.
http://dx.doi.org/10.11113/jt.v62.1879...
verified a L-asparaginase activity of 2.83 U/ml under submerged fermentation. In that work, authors observed that glucose concentrations above 1% inhibited the enzyme production.
Another interesting technique for asparaginase production is the solid-state fermentation, that allows the use of agroindustrial residues as substrate or support.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
Within this context, recently, Dias et al. (2015)7676 Dias FFG, de Castro RJS, Ohara A, Nishide TG, Bagagli MP, Sato HH. Simplex centroid mixture design to improve L-asparaginase production in solid-state fermentation using agroindustrial wastes. Biocatal Agric Biotechnol. 2015, September, http://dx.doi.org/10.1016/j.bcab.2015.09.011.
http://dx.doi.org/10.1016/j.bcab.2015.09...
presented the use of different organic residues (wheat bran, soybean meal, cottonseed meal and orange peel), evaluating the production of L-asparaginase from Aspergillus niger. The maximum enzyme production (94.21 U/g) was obtained after 96 h of fermentation using mixture of wheat bran (1/3), soybean meal (1/3) and cottonseed meal (1/3).
In addition, yeasts have been becoming an interesting alternative for L-asparaginase production. Some investigations have reported, e.g., the use of the yeasts Pichia polymorpha and Candida utilis, for this enzyme production. L-asparaginase of P. polymorpha showed a kM value of 13.7 mM and optimum pH 6.7.7777 Foda MS, Zedan HH, Hashem Sa. Characterization of a novel L-asparaginase produced by Rhodotorula rubra. Rev Latinoam Microbiol. 1980;22(2):87-95. On the other hand, the enzyme produced by C. utilis has kM value of 77 µM.7878 Sakamoto T, Araki C, Beppu T, Arima K. Extracellular asparaginase from candida utilis, its properties as glycoprotein and antitumor activities extracellular asparaginase from candida utilis, its properties as glycoprotein and antitumor activities. Agric Biol Chem Agric BioI Chem. 1977;418(418):1365-1371, http://dx.doi.org/10.1080/00021369.1977.10862699.
http://dx.doi.org/10.1080/00021369.1977....
In a recent study, Soler et al. (2015)77 Soler MF, Pedreira VA, Longo FF, et al. BB 25. Seleção de leveduras produtoras de L-asparaginase em meios sólido e líquido: uma comparação de diferentes metodologias de screening. J Basic Appl Pharm Sci. 2015;36(1). tested 43 different strains of yeasts, verifying that only strains of Issatchenkia orientalis and Rhodotorula glutinis showed periplasmic L-asparaginase activity when growth in liquid CD-m. Also, Sajitha et al. (2015)7979 Sajitha S, Vidya J, Varsha K, Binod P. Cloning and expression of L-asparaginase from E. coli in eukaryotic expression system. Biochem Eng J. 2015;102:14-17, http://dx.doi.org/10.1016/j.bej.2015.02.027.
http://dx.doi.org/10.1016/j.bej.2015.02....
presented an investigation by using an expression study of gene ansB of E. coli, which encodes L-asparaginase enzyme, in yeast. This study was developed on a new protein expression system based on the yeast Pichia pastoris. The resulting enzyme was extracellular and showed activity of 2.5 U/mL at optimum temperature of 37 °C. By these results, authors concluded that this new system of expression could be effective for production of humanized enzyme by glycosylation patterns similar to mammals.7979 Sajitha S, Vidya J, Varsha K, Binod P. Cloning and expression of L-asparaginase from E. coli in eukaryotic expression system. Biochem Eng J. 2015;102:14-17, http://dx.doi.org/10.1016/j.bej.2015.02.027.
http://dx.doi.org/10.1016/j.bej.2015.02....
Industrial production of L-asparaginase
For industrial production of L-asparaginase, many factors need to be taken into account aiming to a process with higher yield and economic viability. For example, type and concentration of carbon and nitrogen sources, pH, aeration, temperature, fermentation time, and, mainly, the microbial agent, have great influence in the process.5252 Gurunathan B, Sahadevan R. Optimization of culture conditions and bench-scale production of L-asparaginase by submerged fermentation of Aspergillus terreus MTCC 1782. J Microbiol Biotechnol. 2012;22(7):923-929, http://dx.doi.org/10.4014/jmb.1112.12002.
http://dx.doi.org/10.4014/jmb.1112.12002...
,8080 Batool T, Makky EA, Jalal M, Yusoff MM. A comprehensive review on L-asparaginase and its applications. Appl Biochem Biotechnol. 2016;178(5):900-923, http://dx.doi.org/10.1007/s12010-015-1917-3.
http://dx.doi.org/10.1007/s12010-015-191...
As previously reported, several microorganisms are presented as L-asparaginase producers; however, bacteria E. coli and E. chrysanthemi are the current main microbial agents for industrial-scale production in pharmaceutical area, while the fungus Aspergillus oryzae is the most used in food industry.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
,8181 Hendriksen HV, Kornbrust BA, Østergaard PR, Stringer MA. Evaluating the potential for enzymatic acrylamide mitigation in a range of food products using an asparaginase from Aspergillus oryzae. J Agric Food Chem. 2009;57(10):4168-4176, http://dx.doi.org/10.1021/jf900174q.
http://dx.doi.org/10.1021/jf900174q...
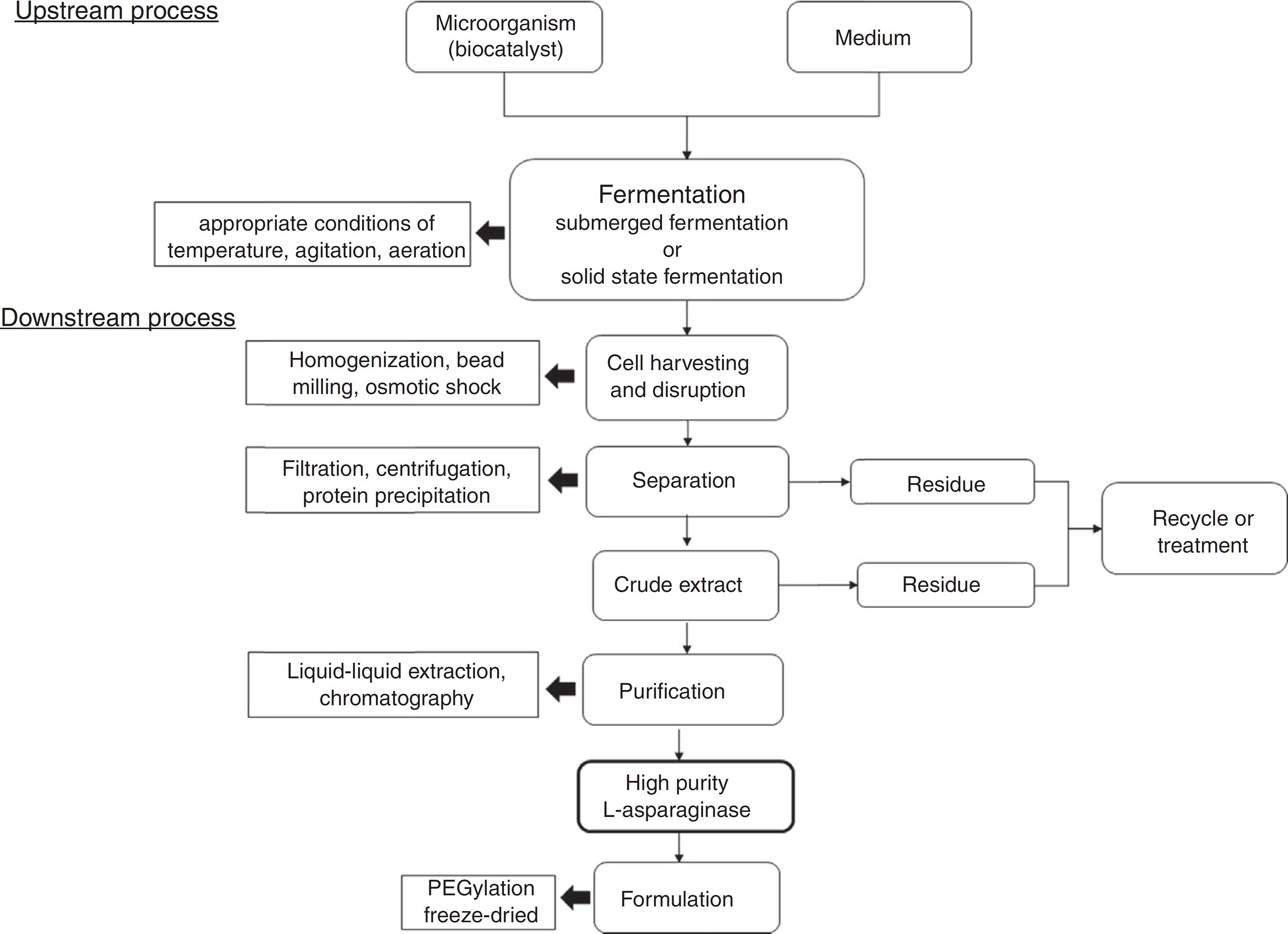
Fig. 3 shows an schematic representation for an industrial process for L-asparaginase production.
Different types of culture medium have been explored for L-asparaginase production. However, carbon source and inductor (nitrogen source) are the more influencing components in the medium. For example, several studies have demonstrated that best inductors for reaching high yields are L-asparagina; 8282 Farag AM, Hassan SW, Beltagy EA, El-Shenawy MA. Optimization of production of anti-tumor L-asparaginase by free and immobilized marine Aspergillus terreus. Egypt J Aquat Res. 2015;41(4):295-302, http://dx.doi.org/10.1016/j.ejar.2015.10.002.
http://dx.doi.org/10.1016/j.ejar.2015.10...
,8383 Kiranmayi MU, Poda S, Vijayalakshmi M. Prodution and optimization of L-asparaginase by an actinobacterium isolated from Nizampatnam mangrove ecosystem. J Environ Biol. 2014;35:799-805.L-glutamine8383 Kiranmayi MU, Poda S, Vijayalakshmi M. Prodution and optimization of L-asparaginase by an actinobacterium isolated from Nizampatnam mangrove ecosystem. J Environ Biol. 2014;35:799-805. and L-proline,3737 Sarquis MI, de M, Oliveira EMM, Santos AS, da Costa GL. Production of L-asparaginase by filamentous fungi. Mem Inst Oswaldo Cruz. 2004;99(5):489-492, http://dx.doi.org/10.1590/S0074-02762004000500005.
http://dx.doi.org/10.1590/S0074-02762004...
,8484 Baskar G, Renganathan S. Production of L-asparaginase from natural substrates by Aspergillus terreus MTCC 1782: effect of substrate, supplementary nitrogen source and L-asparagine. Int J Chem React Eng. 2009;7:.
85 Baskar G, Renganathan S. Evaluation and screening of nitrogen source for L-asparaginase production by Aspergillus terreus MTCC 1782 using latin square design. J Math Stat. 2009;1(2):55-58.-8686 Tippani R, Sivadevuni G. Nutritional factors effecting the production of L-asparaginase by the Fusarium sp. Afr J Biotechnol. 2012;11(15):3692-3696, http://dx.doi.org/10.5897/AJB10.2355.
http://dx.doi.org/10.5897/AJB10.2355...
and the most common carbon source is glucose, in addition to alternative sources such as starch8787 Akilandeswari K, Kavitha K, Vijayalakshmi M. Production of bioactive enzyme L-asparaginase from fungal isolates of water sample through submerged fermentation. Int J Pharm Pharm Sci. 2012;4(suppl 4):363-366. and maltose.8282 Farag AM, Hassan SW, Beltagy EA, El-Shenawy MA. Optimization of production of anti-tumor L-asparaginase by free and immobilized marine Aspergillus terreus. Egypt J Aquat Res. 2015;41(4):295-302, http://dx.doi.org/10.1016/j.ejar.2015.10.002.
http://dx.doi.org/10.1016/j.ejar.2015.10...
,8888 Varalakshmi V, Raju K. Optimization of L-asparaginase production by Aspergillus terreus mtcc 1782 using bajra seed flour under solid state fermentation. Int J Res Eng Technol. 2013;02(09):121-129. http://ijret.org/Volumes/V02/I09/IJRET_110209020.pdf.
http://ijret.org/Volumes/V02/I09/IJRET_1...
L-asparaginase extraction and purification are other pivotal steps for the production of this enzyme. For example, for pharmaceutical application, high level of purification is needed. Other important concern is that most of microorganisms produce intracellular L-asparaginases, with few exceptions.
Different methods for downstream process are reported such as centrifugation, filtration, liquid-liquid extraction, chromatography and protein precipitation. Regarding industrial production, protein precipitation is an advantageous technique due to features such as ease scale up, with simple equipment requirements, low costs and possibility to use large number of precipitants. Additionally, the precipitant agent can be recycled in the final process, reducing the environmental impact associated to its disposal. Actually, precipitation is one of the first steps in the downstream process and it is usually combined with traditional techniques to enhance biomolecules purification and process yield.3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
Also, other highlighted step used for high degree of enzyme purity is chromatography, such as ionic exchange, affinity chromatography, size exclusion, and gel filtration.7171 Mohan Kumar NS, Manonmani HK. Purification, characterization and kinetic properties of extracellular L-asparaginase produced by Cladosporium sp. World J Microbiol Biotechnol. 2013;29(4):577-587, http://dx.doi.org/10.1007/s11274-012-1213-0.
http://dx.doi.org/10.1007/s11274-012-121...
,8989 Khushoo A, Pal Y, Singh BN, Mukherjee KJ. Extracellular expression and single step purification of recombinant Escherichia coli L-asparaginase II. Protein Expr Purif. 2004;38(1):29-36, http://dx.doi.org/10.1016/j.pep.2004.07.009.
http://dx.doi.org/10.1016/j.pep.2004.07....
For example, Lopes et al. (2015)3030 Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol. 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705.
http://dx.doi.org/10.3109/07388551.2015....
reported that the most used purification steps are gel filtration and ion exchange chromatography, which often are preceded by precipitation with (NH4)2SO4. According to authors, considering 50-80% of the total production costs of proteins are provided by extraction and purification steps, optimized downstream can result in significant economic viability.
For pharmaceutical applications, a step of conjugation with polyethylene glycol (PEG), or PEGylation, has been used to improve the compound biostability and bioavailability, influencing in pharmacokinetics and pharmacodynamics properties of the enzyme and reducing the immunological response against this biomolecule.9090 Mehvar R. Modulation of the pharmacokinetics and pharmacodynamics of proteins by polyethylene glycol conjugation. J Pharm Pharm Sci. 2000;3(1):125-136. http://www.ncbi.nlm.nih.gov/pubmed/10954682 Accessed 18.08.16.
http://www.ncbi.nlm.nih.gov/pubmed/10954...
However, this step of PEGylation can also result in loss of biological activity of the conjugate compared with the native enzyme.9191 Veronese FM, Pasut G. PEGylation, successful approach to drug delivery. Drug Discov Today. 2005;10(21):1451-1458, http://dx.doi.org/10.1016/S1359-6446(05)03575-0.
http://dx.doi.org/10.1016/S1359-6446(05)...
On the other hand, this step is not required in food industry application.
Freeze-drying is other important step to improve the long and short-term storage of the enzyme L-asparaginase formulation. It can prevent most water-related reactions by sublimating water from the frozen product under vacuum, also allowing sterile drying without heating or chemical sterilization. However, problems related to cold denaturation, freeze denaturation and osmotic pressure increase due to dehydration and cryoconcentration.9292 Singh S, Kolhe P, Wang W, Nema S. Large-scale freezing of biologics - a practitioner's review, part one: fundamental aspects. Bioprocess Int. 2009;(7):32-44.
Conclusion and future recommendations
L-asparaginase is an interesting enzyme with important applications in pharmaceutical and food industry. However, its use in these industrial sectors requires some specific properties, as security for use by humans. As chemotherapeutic agent, an efficient action is required, in addition to reduced adverse effects, such as hypersensitivity and immune inactivation. In food, this enzyme helps to decrease the concentrations of acrylamide (carcinogenic compound for humans) formed in the process, maintaining their nutritional and sensory properties. Thus, research work seeking for new L-asparaginases, mainly produced by eukaryotic microorganisms, instead of bacterial enzymes currently used, has potential to obtain new enzymes with desirable properties. These discoveries have to be followed by an intensive work aiming to increase the process productivity to enable and extend the use of this enzyme, mainly in food industries. Taking this into account, tools of molecular biology are useful, although even a more traditional work of biochemical engineering have not been extensively related in literature, indicating needs of further works such as different process configuration evaluation, as well as use of bioreactors options.
Acknowledgments
The authors would like to thank FAPESP (Process 2014/27055-2), CNPq, CAPES and Programa Estudantes-Convênio de Pós-Graduação - PEC-PG, da CAPES/CNPq - Brazil for financial support, and the Coleção de Culturas Tropical Fundação André Tosello.
REFERENCES
-
1Appel IM, van Kessel-Bakvis C, Stigter R, Pieters R. Influence of two different regimens of concomitant treatment with asparaginase and dexamethasone on hemostasis in childhood acute lymphoblastic leukemia. Leukemia 2007;21(11):2377-2380, http://dx.doi.org/10.1038/sj.leu.2404793
» http://dx.doi.org/10.1038/sj.leu.2404793 -
2Mohan Kumar NS, Shimray CA, Indrani D, Manonmani HK. Reduction of acrylamide formation in sweet bread with L-asparaginase treatment. Food Bioprocess Technol 2013;7(3):741-748, http://dx.doi.org/10.1007/s11947-013-1108-6
» http://dx.doi.org/10.1007/s11947-013-1108-6 -
3Medeiros Vinci R, Mestdagh F, De Meulenaer B. Acrylamide formation in fried potato products - present and future, a critical review on mitigation strategies. Food Chem 2012;133(4):1138-1154, http://dx.doi.org/10.1016/j.foodchem.2011.08.001
» http://dx.doi.org/10.1016/j.foodchem.2011.08.001 -
4Keating MJ, Holmes R, Lerner S, Ho DH. L-asparaginase and PEG asparaginase - past, present, and future. Leuk Lymphoma 1993;10(suppl):153-157, http://dx.doi.org/10.3109/10428199309149129
» http://dx.doi.org/10.3109/10428199309149129 -
5Narta UK, Kanwar SS, Azmi W. Pharmacological and clinical evaluation of L-asparaginase in the treatment of leukemia. Crit Rev Oncol Hematol 2007;61(3):208-221, http://dx.doi.org/10.1016/j.critrevonc.2006.07.009
» http://dx.doi.org/10.1016/j.critrevonc.2006.07.009 -
6Dange V, Peshwe S. Purification and biochemical characterization of L-asparaginase from Aspergillus niger and evaluation of its antineoplastic activity. Int J Sci Res 2015;4(2):564-569.
-
7Soler MF, Pedreira VA, Longo FF, et al. BB 25. Seleção de leveduras produtoras de L-asparaginase em meios sólido e líquido: uma comparação de diferentes metodologias de screening. J Basic Appl Pharm Sci 2015;36(1).
-
8Verma N, Kumar K, Kaur G, Anand S. L-asparaginase: a promising chemotherapeutic agent. Crit Rev Biotechnol 2007;27(1):45-62, http://dx.doi.org/10.1080/07388550601173926
» http://dx.doi.org/10.1080/07388550601173926 -
9Hill JM. L-asparaginase therapy for leukemia and other malignant neoplasms. J Am Med Assoc 1967;202(9):882, http://dx.doi.org/10.1001/jama.1967.03130220070012
» http://dx.doi.org/10.1001/jama.1967.03130220070012 -
10El-Bessoumy AA, Sarhan M, Mansour J. Production, isolation, and purification of L-asparaginase from Pseudomonas aeruginosa 50071 using solid-state fermentation. J Biochem Mol Biol 2004;37(4):387-393, http://dx.doi.org/10.5483/BMBRep.2004.37.4.387
» http://dx.doi.org/10.5483/BMBRep.2004.37.4.387 -
11Shrivastava A, Khan AA, Khurshid M, Kalam MA, Jain SK, Singhal PK. Recent developments in L-asparaginase discovery and its potential as anticancer agent. Crit Rev Oncol Hematol 2016;100:1-10, http://dx.doi.org/10.1016/j.critrevonc.2015.01.002
» http://dx.doi.org/10.1016/j.critrevonc.2015.01.002 -
12Van den Berg H. Asparaginase revisited. Leuk Lymphoma 2011;52(2):168-178, http://dx.doi.org/10.3109/10428199309149127
» http://dx.doi.org/10.3109/10428199309149127 -
13Noronkoski T, Stoineva IB, Ivanov IP, Petkov DD, Mononen I. Glycosylasparaginase-catalyzed synthesis and hydrolysis of -aspartyl peptides. J Biol Chem 1998;273(41):26295-26297, http://dx.doi.org/10.1074/jbc.273.41.26295
» http://dx.doi.org/10.1074/jbc.273.41.26295 -
14Lang S. Uber desamidierung im Tierkorper. Beitr Chem Physiol Pathol 1904;5:321-345.
-
15Furth O, Friedmann M. Uber die Verbreitung asparaginspaltender Organfermente. Biochemistry 1910;26:435-440.
-
16Clementi A. La desamidation enzymatique de l’asparagine chez les differentes especes animals et la signification physiologique de sa presence dans l’organisme. Arch Intern Physiol 1922;19:369-398.
-
17Mashburn LT, Wriston JC. Tumor inhibitory effect of L-asparaginase from Eschericha coli Arch Biochem Biophys 1964;105:450-452.
-
18Broome JD. Antilymphoma activity of L-asparaginase in vivo: clearance rates of enzyme preparations from guinea pig serum and yeast in relation to their effect on tumor growth. J Natl Cancer Inst 1965;35(6):967-974.
-
19Schrappe M, Reiter A, Ludwig WD, et al. Improved outcome in childhood acute lymphoblastic leukemia despite reduced use of anthracyclines and cranial radiotherapy: results of trial ALL-BFM 90. German-Austrian-Swiss ALL-BFM Study Group. Blood 2000;95(11):3310-3322. http://www.ncbi.nlm.nih.gov/pubmed/10828010 Accessed 09.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/10828010 -
20Kiriyama Y, Kubota M, Takimoto T, et al. Biochemical characterization of U937 cells resistant to L-asparaginase: the role of asparagine synthetase. Leukemia 1989;3(4):294-297. http://www.ncbi.nlm.nih.gov/pubmed/2564453 Accessed 17.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/2564453 -
21Stams WAG, den Boer ML, Beverloo HB, et al. Sensitivity to L-asparaginase is not associated with expression levels of asparagine synthetase in t(12;21)+ pediatric ALL. Blood 2003;101(7):2743-2747, http://dx.doi.org/10.1182/blood-2002-08-2446
» http://dx.doi.org/10.1182/blood-2002-08-2446 -
22Friedman M. Chemistry, biochemistry, and safety of acrylamide. A review. J Agric Food Chem 2003;51(16):4504-4526, http://dx.doi.org/10.1021/jf030204+
» http://dx.doi.org/10.1021/jf030204+ -
23Tareke E, Rydberg P, Karlsson P, Eriksson S, Törnqvist M. Acrylamide: a cooking carcinogen? Chem Res Toxicol 2000;13(6):517-522. http://www.ncbi.nlm.nih.gov/pubmed/10858325 Accessed 18.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/10858325 -
24Zyzak DV, Sanders RA, Stojanovic M, et al. Acrylamide formation mechanism in heated foods. J Agric Food Chem 2003;51(16):4782-4787, http://dx.doi.org/10.1021/jf034180i
» http://dx.doi.org/10.1021/jf034180i -
25Zhang S, Xie Y, Zhang C, et al. Biochemical characterization of a novel L-asparaginase from Bacillus megaterium H-1 and its application in French fries. Food Res Int 2015;77:527-533, http://dx.doi.org/10.1016/j.foodres.2015.08.031
» http://dx.doi.org/10.1016/j.foodres.2015.08.031 -
26Huang L, Liu Y, Sun Y, Yan Q, Jiang Z. Biochemical characterization of a novel L-asparaginase with low glutaminase activity from Rhizomucor miehei and its application in food safety and leukemia treatment. Appl Environ Microbiol 2014;80(5):1561-1569, http://dx.doi.org/10.1128/AEM.03523-13
» http://dx.doi.org/10.1128/AEM.03523-13 -
27Anese M, Quarta B, Peloux L, Calligaris S. Effect of formulation on the capacity of L-asparaginase to minimize acrylamide formation in short dough biscuits. Food Res Int 2011;44(9):2837-2842, http://dx.doi.org/10.1016/j.foodres.2011.06.025
» http://dx.doi.org/10.1016/j.foodres.2011.06.025 -
28Pedreschi F, Mariotti S, Granby K, Risum J. Acrylamide reduction in potato chips by using commercial asparaginase in combination with conventional blanching. LWT - Food Sci Technol 2011;44(6):1473-1476, http://dx.doi.org/10.1016/j.lwt.2011.02.004
» http://dx.doi.org/10.1016/j.lwt.2011.02.004 -
29Wriston JC, Yellin TO. L-asparaginase: a review. Adv Enzymol Relat Areas Mol Biol 1973;39:185-248. http://www.ncbi.nlm.nih.gov/pubmed/4583638 Accessed 23.06.16.
» http://www.ncbi.nlm.nih.gov/pubmed/4583638 -
30Lopes AM, Oliveira-Nascimento L, de Ribeiro A, et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Crit Rev Biotechnol 2015;8551(December):1-18, http://dx.doi.org/10.3109/07388551.2015.1120705
» http://dx.doi.org/10.3109/07388551.2015.1120705 -
31Deshpande N, Choubey P, Agashe M. Studies on optimization of growth parameters for L-asparaginase production by streptomyces ginsengisoli. Sci World J 2014;2014:1-6, http://dx.doi.org/10.1155/2014/895167
» http://dx.doi.org/10.1155/2014/895167 -
32Degroot N, Lichtenstein N. The action of Pseudomonas fluorescens extracts on asparagine and asparagine derivatives. Biochim Biophys Acta 1960;40:99-110.
-
33Rowley B, Wriston JCJ. Partial purification and antilymphoma activity of Serratia marcescens L-asparaginase. Biochem Biophys Res Commun 1967;28(2):160-165.
-
34Kozak M, Jurga S. A comparison between the crystal and solution structures of Escherichia coli asparaginase II. Acta Biochim Pol 2002;49(2):509-513. http://www.ncbi.nlm.nih.gov/pubmed/12362993 Accessed 17.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/12362993 -
35Wade HE, Elsworth R, Herbert D, Keppie J, Sargeant K. A new L-asparaginase with antitumour activity? Lancet 1968;2(7571):776-777.
-
36Tosa T, Sano R, Yamamoto K, Nakamura M, Ando K, Chibatha I. L-asparaginase from Proteus vulgaris Appl Microbiol 1972;22:387-392.
-
37Sarquis MI, de M, Oliveira EMM, Santos AS, da Costa GL. Production of L-asparaginase by filamentous fungi. Mem Inst Oswaldo Cruz 2004;99(5):489-492, http://dx.doi.org/10.1590/S0074-02762004000500005
» http://dx.doi.org/10.1590/S0074-02762004000500005 -
38Duval M. Comparison of Escherichia coli-asparaginase with Erwinia asparaginase in the treatment of childhood lymphoid malignancies: results of a randomized European Organisation for Research and Treatment of Cancer - Children's Leukemia Group phase 3 trial. Blood 2002;99(8):2734-2739, http://dx.doi.org/10.1182/blood.V99.8.2734
» http://dx.doi.org/10.1182/blood.V99.8.2734 -
39Zuo S, Zhang T, Jiang B, Mu W. Recent research progress on microbial L-asparaginases. Appl Microbiol Biotechnol 2014;99(3):1069-1079, http://dx.doi.org/10.1007/s00253-014-6271-9
» http://dx.doi.org/10.1007/s00253-014-6271-9 -
40Casale T Del, Sollitti P, Chesney RH. Cytoplasmic L-asparaginase: isolation of a defective strain and mapping of ansA. J Bacteriol 1983;154(1):513-515.
-
41Whitecar JP, Bodey GP, Harris JE, Freireich EJ. L-asparaginase. N Engl J Med 1970;282(13):732-734, http://dx.doi.org/10.1056/NEJM197003262821307
» http://dx.doi.org/10.1056/NEJM197003262821307 -
42Kumar D, Sobha K. L-asparaginase from Microbes: a comprehensive review. Adv Biores 2012;3(December):137-157. http://soeagra.com/abr/abrdec 2012/22.pdf
» http://soeagra.com/abr/abrdec 2012/22.pdf -
43Müller HJ, Boos J. Use of L-asparaginase in childhood ALL. Crit Rev Oncol Hematol 1998;28(2):97-113. http://www.ncbi.nlm.nih.gov/pubmed/9768345 Accessed 18.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/9768345 -
44Kumar S, Dasu VV, Pakshirajan K. Localization and production of novel L-asparaginase from Pectobacterium carotovorum MTCC 1428. Process Biochem 2010;45(2):223-229, http://dx.doi.org/10.1016/j.procbio.2009.09.011
» http://dx.doi.org/10.1016/j.procbio.2009.09.011 -
45Aghaiypour K, Wlodawer A, Lubkowski J. Structural basis for the activity and substrate specificity of Erwinia chrysanthemi L-asparaginase. Biochemistry 2001;40(19):5655-5664. http://www.ncbi.nlm.nih.gov/pubmed/11341830 Accessed 17.02.16.
» http://www.ncbi.nlm.nih.gov/pubmed/11341830 -
46Sreenivasulu V, Jayaveera K, Rao PM. Solid-state fermentation for the production of L-asparaginase by Aspergillus Sp. Res J Pharmacogn Phytochem 2009;1(1):21-25. http://www.indianjournals.com/ijor.aspx?target=ijor:rjpp&volume=1&issue=1&article=006 Accessed 15.02.16.
» http://www.indianjournals.com/ijor.aspx?target=ijor:rjpp&volume=1&issue=1&article=006 -
47Mishra A. Production of L-asparaginase, an anticancer agent, from Aspergillus niger using agricultural waste in solid state fermentation. Appl Biochem Biotechnol 2006;135(1):33-42, http://dx.doi.org/10.1385/ABAB:135:1:33
» http://dx.doi.org/10.1385/ABAB:135:1:33 -
48Theantana T, Hyde KD, Lumyong S. Asparaginase production by endophytic fungi from Thai medicinal plants: cytoxicity properties sources of endophytic fungi isolation of endophytic fungi identification of endophytic fungi Identification was based on colony and hyphal. Int J Integr Biol 2009;7(1):1-8.
-
49Theantana T, Hyde KD, Lumyong S, Mai C. Aparaginase production by endophytic fungi isolated from some Thai medicinal plants. KMITL Sci Tech J 2007;7:13-18.
-
50Thirunavukkarasu N, Suryanarayanan TS, Murali TS, Ravishankar JP, Gummadi SN. L-asparaginase from marine derived fungal endophytes of seaweeds. Mycosphere 2011;2(2):147-155, http://dx.doi.org/10.1111/j.1365-313X.2009.03887.x.zation
» http://dx.doi.org/10.1111/j.1365-313X.2009.03887.x.zation -
51Balasubramanian K, Ambikapathy V, Panneerselvam A. Production, isolation and purification of L-asparaginase from Aspergillus terreus using submerged fermentation. Int J Adv Pharm Res 2012;3(2):778-783.
-
52Gurunathan B, Sahadevan R. Optimization of culture conditions and bench-scale production of L-asparaginase by submerged fermentation of Aspergillus terreus MTCC 1782. J Microbiol Biotechnol 2012;22(7):923-929, http://dx.doi.org/10.4014/jmb.1112.12002
» http://dx.doi.org/10.4014/jmb.1112.12002 -
53Anjum Zia MAZ, Bashir R, Ahmed I, Iftikhar T. Production of L-asparaginase from Aspergillus Niger using agro wastes by-products in submerged fermentation process. J Teknol 2013;62(2), http://dx.doi.org/10.11113/jt.v62.1879
» http://dx.doi.org/10.11113/jt.v62.1879 -
54Pokrovskaya MV, Aleksandrova SS, Pokrovsky VS, et al. Cloning, expression and characterization of the recombinant Yersinia pseudotuberculosis L-asparaginase. Protein Expr Purif 2012;82(1):150-154, http://dx.doi.org/10.1016/j.pep.2011.12.005
» http://dx.doi.org/10.1016/j.pep.2011.12.005 -
55Singh Y, Gundampati RK, Jagannadham MV, Srivastava SK. Extracellular L-asparaginase from a protease-deficient bacillus aryabhattai ITBHU02: purification, biochemical characterization, and evaluation of antineoplastic activity in vitro. Appl Biochem Biotechnol 2013;171(7):1759-1774, http://dx.doi.org/10.1007/s12010-013-0455-0
» http://dx.doi.org/10.1007/s12010-013-0455-0 -
56Prema P, Devi MN, Alagumanikumaran N. Production of tumor inhibitory L-asparaginase by wild and mutant strains of Pseudomonas fluorescens Int J Adv Res J 2013;1(4):163-171.
-
57Kishore V, Nishita KP, Manonmani HK. Cloning, expression and characterization of L-asparaginase from Pseudomonas fluorescens for large scale production in E. coli BL21. 3 Biotech 2015;5(6):975-981, http://dx.doi.org/10.1007/s13205-015-0300-y
» http://dx.doi.org/10.1007/s13205-015-0300-y -
58Mahajan RV, Kumar V, Rajendran V, Saran S, Ghosh PC, Saxena RK. Purification and characterization of a novel and robust L-asparaginase having low-glutaminase activity from Bacillus licheniformis: in vitro evaluation of anti-cancerous properties, trackman PC. PLoS ONE 2014;9(6):e99037, http://dx.doi.org/10.1371/journal.pone.0099037
» http://dx.doi.org/10.1371/journal.pone.0099037 -
59Meena B, Anburajan L, Dheenan PS, et al. Novel glutaminase free L-asparaginase from Nocardiopsis alba NIOT-VKMA08: production, optimization, functional and molecular characterization. Bioprocess Biosyst Eng 2015;38(2):373-388, http://dx.doi.org/10.1007/s00449-014-1277-3
» http://dx.doi.org/10.1007/s00449-014-1277-3 -
60Meena B, Anburajan L, Vinithkumar NV, et al. Molecular expression of L-asparaginase gene from Nocardiopsis alba NIOT-VKMA08 in Escherichia coli: a prospective recombinant enzyme for leukaemia chemotherapy. Gene 2016, http://dx.doi.org/10.1016/j.gene.2016.05.003
» http://dx.doi.org/10.1016/j.gene.2016.05.003 -
61Dharmaraj S. Study of L-asparaginase production by Streptomyces noursei MTCC 10469, isolated from marine sponge Callyspongia diffusa. Iran J Biotechnol 2011;9(2):102-108.
-
62Usha R, Mala KK, Venil CK, Palaniswamy M. Screening of actinomycetes from mangrove ecosystem for L-asparaginase activity and optimization by response surface methodology. Polish J Microbiol/Pol Tow Mikrobiol=Polish Soc Microbiol 2011;60(3):213-221. http://www.ncbi.nlm.nih.gov/pubmed/22184928 Accessed 18.08.16.
» http://www.ncbi.nlm.nih.gov/pubmed/22184928 -
63Jia M, Xu M, He B, Rao Z. Cloning, expression, and characterization of L-asparaginase from a newly isolated Bacillus subtilis B11-06. J Agric Food Chem 2013;61(39):9428-9434, http://dx.doi.org/10.1021/jf402636w
» http://dx.doi.org/10.1021/jf402636w -
64Hatanaka T, Usuki H, Arima J, et al. Extracellular production and characterization of two Streptomyces L-asparaginases. Appl Biochem Biotechnol 2011;163(7):836-844, http://dx.doi.org/10.1007/s12010-010-9087-9
» http://dx.doi.org/10.1007/s12010-010-9087-9 -
65Yaacob MA, Hasan WANW, Ali MSM, et al. Characterisation and molecular dynamic simulations of J15 asparaginase from Photobacterium sp. strain J15. Acta Biochim Pol 2014;61(4):745-752. http://www.ncbi.nlm.nih.gov/pubmed/25337608 Accessed 18.08.16.
» http://www.ncbi.nlm.nih.gov/pubmed/25337608 -
66Bansal S, Gnaneswari D, Mishra P, Kundu B. Structural stability and functional analysis of L-asparaginase from Pyrococcus furiosus. Biochemistry 2010;75(3):375-381, http://dx.doi.org/10.1134/S0006297910030144
» http://dx.doi.org/10.1134/S0006297910030144 -
67Sudhir AP, Agarwaal VV, Dave BR, Patel DH, Subramanian RB. Enhanced catalysis of L-asparaginase from Bacillus licheniformis by a rational redesign. Enzyme Microb Technol 2016;86:1-6, http://dx.doi.org/10.1016/j.enzmictec.2015.11.010
» http://dx.doi.org/10.1016/j.enzmictec.2015.11.010 -
68Krishnapura PR, Belur PD. Partial purification and characterization of L-asparaginase from an endophytic Talaromyces pinophilus isolated from the rhizomes of Curcuma amada. J Mol Catal B Enzym 2016;124:83-91, http://dx.doi.org/10.1016/j.molcatb.2015.12.007
» http://dx.doi.org/10.1016/j.molcatb.2015.12.007 -
69Lincoln L, Niyonzima FN, More SS. Purification and properties of a fungal L-asparaginase from trichoderma viride pers: sf grey. J Microbiol Biotechnol food Sci 2015, http://dx.doi.org/10.15414/JMBFS.2014.4.4.310-316
» http://dx.doi.org/10.15414/JMBFS.2014.4.4.310-316 -
70Dange VU, Peshwe SA. Production, purification andcharacterizationof fungal L-asparaginase. Bionano Front 2011;4:162-167.
-
71Mohan Kumar NS, Manonmani HK. Purification, characterization and kinetic properties of extracellular L-asparaginase produced by Cladosporium sp. World J Microbiol Biotechnol 2013;29(4):577-587, http://dx.doi.org/10.1007/s11274-012-1213-0
» http://dx.doi.org/10.1007/s11274-012-1213-0 -
72Shrivastava A, Khan AA, Shrivastav A, Jain SK, Singhal PK. Kinetic studies of L-asparaginase from Penicillium digitatum Prep Biochem Biotechnol 2012;42(6):574-581, http://dx.doi.org/10.1080/10826068.2012.672943
» http://dx.doi.org/10.1080/10826068.2012.672943 -
73Patro KR, Gupta N. Extraction, purification and characterization of L-asparaginase from Penicillium sp. by submerged fermentation. Int J Biotechnol Mol Biol Res 2012;3(3):30-34, http://dx.doi.org/10.5897/IJBMBR11.066
» http://dx.doi.org/10.5897/IJBMBR11.066 -
74Elshafei AM, Hassan MM, Abouzeid MA-E, Mahmoud DA, Elghonemy DH. Purification, characterization and antitumoractivity of L-asparaginase from Penicillium brevicompactum NRC 829. Br Microbiol Res J 2012;2(3):158-174.
-
75Thakur M, Lincoln L, Niyonzima FN, More SS. Isolation, purification and characterization of fungal extracellular L-asparaginase from Mucor hiemalis J Biocatal Biotransform 2013;2(2), http://dx.doi.org/10.4172/2324-9099.1000108
» http://dx.doi.org/10.4172/2324-9099.1000108 -
76Dias FFG, de Castro RJS, Ohara A, Nishide TG, Bagagli MP, Sato HH. Simplex centroid mixture design to improve L-asparaginase production in solid-state fermentation using agroindustrial wastes. Biocatal Agric Biotechnol. 2015, September, http://dx.doi.org/10.1016/j.bcab.2015.09.011
» http://dx.doi.org/10.1016/j.bcab.2015.09.011 -
77Foda MS, Zedan HH, Hashem Sa. Characterization of a novel L-asparaginase produced by Rhodotorula rubra. Rev Latinoam Microbiol 1980;22(2):87-95.
-
78Sakamoto T, Araki C, Beppu T, Arima K. Extracellular asparaginase from candida utilis, its properties as glycoprotein and antitumor activities extracellular asparaginase from candida utilis, its properties as glycoprotein and antitumor activities. Agric Biol Chem Agric BioI Chem 1977;418(418):1365-1371, http://dx.doi.org/10.1080/00021369.1977.10862699
» http://dx.doi.org/10.1080/00021369.1977.10862699 -
79Sajitha S, Vidya J, Varsha K, Binod P. Cloning and expression of L-asparaginase from E. coli in eukaryotic expression system. Biochem Eng J 2015;102:14-17, http://dx.doi.org/10.1016/j.bej.2015.02.027
» http://dx.doi.org/10.1016/j.bej.2015.02.027 -
80Batool T, Makky EA, Jalal M, Yusoff MM. A comprehensive review on L-asparaginase and its applications. Appl Biochem Biotechnol 2016;178(5):900-923, http://dx.doi.org/10.1007/s12010-015-1917-3
» http://dx.doi.org/10.1007/s12010-015-1917-3 -
81Hendriksen HV, Kornbrust BA, Østergaard PR, Stringer MA. Evaluating the potential for enzymatic acrylamide mitigation in a range of food products using an asparaginase from Aspergillus oryzae J Agric Food Chem 2009;57(10):4168-4176, http://dx.doi.org/10.1021/jf900174q
» http://dx.doi.org/10.1021/jf900174q -
82Farag AM, Hassan SW, Beltagy EA, El-Shenawy MA. Optimization of production of anti-tumor L-asparaginase by free and immobilized marine Aspergillus terreus Egypt J Aquat Res 2015;41(4):295-302, http://dx.doi.org/10.1016/j.ejar.2015.10.002
» http://dx.doi.org/10.1016/j.ejar.2015.10.002 -
83Kiranmayi MU, Poda S, Vijayalakshmi M. Prodution and optimization of L-asparaginase by an actinobacterium isolated from Nizampatnam mangrove ecosystem. J Environ Biol 2014;35:799-805.
-
84Baskar G, Renganathan S. Production of L-asparaginase from natural substrates by Aspergillus terreus MTCC 1782: effect of substrate, supplementary nitrogen source and L-asparagine. Int J Chem React Eng 2009;7:.
-
85Baskar G, Renganathan S. Evaluation and screening of nitrogen source for L-asparaginase production by Aspergillus terreus MTCC 1782 using latin square design. J Math Stat 2009;1(2):55-58.
-
86Tippani R, Sivadevuni G. Nutritional factors effecting the production of L-asparaginase by the Fusarium sp. Afr J Biotechnol 2012;11(15):3692-3696, http://dx.doi.org/10.5897/AJB10.2355
» http://dx.doi.org/10.5897/AJB10.2355 -
87Akilandeswari K, Kavitha K, Vijayalakshmi M. Production of bioactive enzyme L-asparaginase from fungal isolates of water sample through submerged fermentation. Int J Pharm Pharm Sci 2012;4(suppl 4):363-366.
-
88Varalakshmi V, Raju K. Optimization of L-asparaginase production by Aspergillus terreus mtcc 1782 using bajra seed flour under solid state fermentation. Int J Res Eng Technol 2013;02(09):121-129. http://ijret.org/Volumes/V02/I09/IJRET_110209020.pdf
» http://ijret.org/Volumes/V02/I09/IJRET_110209020.pdf -
89Khushoo A, Pal Y, Singh BN, Mukherjee KJ. Extracellular expression and single step purification of recombinant Escherichia coli L-asparaginase II. Protein Expr Purif 2004;38(1):29-36, http://dx.doi.org/10.1016/j.pep.2004.07.009
» http://dx.doi.org/10.1016/j.pep.2004.07.009 -
90Mehvar R. Modulation of the pharmacokinetics and pharmacodynamics of proteins by polyethylene glycol conjugation. J Pharm Pharm Sci 2000;3(1):125-136. http://www.ncbi.nlm.nih.gov/pubmed/10954682 Accessed 18.08.16.
» http://www.ncbi.nlm.nih.gov/pubmed/10954682 -
91Veronese FM, Pasut G. PEGylation, successful approach to drug delivery. Drug Discov Today 2005;10(21):1451-1458, http://dx.doi.org/10.1016/S1359-6446(05)03575-0
» http://dx.doi.org/10.1016/S1359-6446(05)03575-0 -
92Singh S, Kolhe P, Wang W, Nema S. Large-scale freezing of biologics - a practitioner's review, part one: fundamental aspects. Bioprocess Int 2009;(7):32-44.
Publication Dates
-
Publication in this collection
Dec 2016
History
-
Received
22 Aug 2016 -
Accepted
06 Sept 2016

 Source: Based on Hill (1967)
Source: Based on Hill (1967) Source: Based on Van den Berg (2011).
Source: Based on Van den Berg (2011).