Abstract
This study aimed to evaluate the tolerance to salinity and temperature, the genetic diversity and the symbiotic efficiency of rhizobia isolates obtained from wild genotypes of common bean cultivated in soil samples from the States of Goiás, Minas Gerais and Paraná. The isolates were subjected to different NaCl concentrations (0%, 1%, 2%, 4% and 6%) at different temperatures (28 °C, 33 °C, 38 °C, 43 °C and 48 °C). Genotypic characterization was performed based on BOX-PCR, REP-PCR markers and 16S rRNA sequencing. An evaluation of symbiotic efficiency was carried out under greenhouse conditions in autoclaved Leonard jars. Among 98 isolates about 45% of them and Rhizobium freirei PRF81 showed a high tolerance to temperature, while 24 isolates and Rhizobium tropici CIAT899 were able to use all of the carbon sources studied. Clustering analysis based on the ability to use carbon sources and on the tolerance to salinity and temperature grouped 49 isolates, R. tropici CIAT899 and R. tropici H12 with a similarity level of 76%. Based on genotypic characterization, 65% of the isolates showed an approximately 66% similarity with R. tropici CIAT899 and R. tropici H12. About 20% of the isolates showed symbiotic efficiency similar to or better than the best Rhizobium reference strain (R. tropici CIAT899). Phylogenetic analysis of the 16S rRNA revealed that two efficient isolates (ALSG5A1 and JPrG6A8) belong to the group of strains used as commercial inoculant for common bean in Brazil and must be assayed in field experiments.
Keywords:
Phaseolus vulgaris; Rhizobium; Symbiotic efficiency; Carbon source use; Tolerance to salinity and temperature
Introduction
The common bean (Phaseolus vulgaris L.) is a leguminous plant of worldwide social and economic importance, providing most of the daily requirements of protein and carbohydrates for the poorest populations of South and Central America, Africa and India.11 Brougthon WJ, Hernández G, Blair M, Beebe S, Gepts P, Vardeyleyden J. Bean (Phaseolus ssp.): model food legume. Plant Soil. 2003;252:55-128. With respect to international agriculture, Brazil is the world's third largest producer of common bean, accounting for 12.7% of worldwide production.22 FAO – Food and Agricultural Organization of the United Nations; 2015. Available at: http://faostat3.fao.org/download/Q/QC/E. Accessed 19.05.15.
http://faostat3.fao.org/download/Q/QC/E...
In Brazil, the common bean is cultivated on a total area of 3.1 million hectares with a total grain production of approximately 2.8 million tons,33 Acompanhamento de safra brasileira: grãos, décimo levantamento. Brasília: CONAB – Companhia Nacional de Abastecimento; 2013. Available at: http://www.conab.gov.br/OlalaCMS/uploads/arquivos/13_07_09_09_04_53_boletim_graos_junho_2013.pdf Accessed 09.02.14.
http://www.conab.gov.br/OlalaCMS/uploads...
for which high amounts of nitrogen (N) are required.
Despite its abundance in the atmosphere, N is scarce in tropical soils due to the fast mineralization of organic matter in tropical conditions. Although the decomposition of organic matter is an important source of N for crops, the adequate supply of N to crops depends largely on the use of nitrogen fertilizers.44 Soares BL [D.Sc. Thesis] Avaliação técnica econômica do feijoeiro-comum inoculado com rizóbio em diferentes ambientes. Lavras, Brasil: Universidade Federal de Lavras; 2012, 150 pp. However, biological nitrogen fixation (BNF) is considered a more sustainable approach for supplying N to the production system.
BNF is a key process for the conversion of nitrogen gas (N2) into ammonia (NH3) performed by bacteria belonging to the group of rhizobia. The reduction reaction of N2 to NH3 is carried out by N-fixing bacteria or diazotrophic microorganisms containing the enzymatic complex in which nitrogenase takes part.55 Novais RF, Alvarez VVH, Barros NF, Fontes RLF, Cantarutti RB, Neves JCL. Fertilidade do Solo. Viçosa: Sociedade Brasileira de Ciência do Solo; 2007, 1017 pp. Among N-fixing bacteria of the rhizobia group, a variety of Rhizobium and Ensifer species is able to colonize and establish a symbiotic partnership with common bean.66 Dall’Agnol RF, Ribeiro RA, Ormeño-Orrillo E, et al. Rhizobium freirei sp. nov., a symbiont of Phaseolus vulgaris that is very effective at fixing nitrogen. Int J Syst Evol Microbiol. 2013;63:4167-4173.,77 Mhamdi R, Ardley J, Tian R, et al. High-quality permanent draft genome sequence of Ensifer meliloti strain 4H41, an effective salt- and drought-tolerant microsymbiont of Phaseolus vulgaris. Stand Genomic Sci. 2015;10:1-7.
To improve BNF efficiency, more efficient rhizobia strains are needed. Many isolating works have been performed using soil from different sites; however, as trap plant usually is used a commercial variety of common bean. The strategy used in our work was to collect soil in different sites and to use wild genotypes of common bean as trap plant looking for a better exploration of the rhizobial community, since wild genotypes show a broader genetic base. This work aimed to characterize and determine the symbiotic efficiency of rhizobia isolates obtained from the root nodules of wild genotypes of common bean.
Materials and methods
Bacterial strains and rhizobia isolates
The isolates evaluated in this work were obtained by Sampaio FB88 Sampaio FB [M.Sc. Dissertation] Isolados de rizóbios capturados de genótipos silvestres de feijoeiro: obtenção, morfologia e uso de fontes de carbono. Goiânia, Brasil: Universidade Federal de Goiás; 2013, 85 pp. and are available at the Collection of Microorganisms and Multifunctional Fungi of Embrapa Rice and Beans. Strains of Rhizobium tropici (CIAT899 and H12), Rhizobium freirei (PRF81) were used as reference strains in all analyses and, Rhizobium etli bv. phaseoli (CFN42) used in the BOX- and REP-PCR analyses.
Carbon source use (CSU) and tolerance to salinity and temperature (TST) assays
CSU was assayed for 98 isolates and for the R. tropici reference strains. Bacteria were kept for growth on modified YMA (Yeast Mannitol Agar) culture medium, without mannitol, added with individual carbon sources sucrose, glucose, malic acid, maleic acid, nicotinic acid, inositol, sorbitol, arabinose, fructose and glycerol. After incubation at 28 °C, bacterial growth was verified from 48 to 96 h at each 24 h.
The same isolates and reference strains were assessed for TST on YMA culture medium on a factorial (5 × 5) arrangement (concentrations of NaCl - 0%, 1%, 2%, 4%, 6% and temperature - 28 °C, 33 °C, 38 °C, 43 °C, 48 °C) incubated for a period of 48 h.
Genotypic characterization based on molecular markers
Based on CSU and TST, 55 isolates were selected for genotypic characterization. Genomic DNA was extracted according to Ausubel et al.99 Ausubel F, Brent R, Kingston R, et al. Short Protocols in Molecular Biology. 4th ed. New York: John Wiley and Sons, Inc.; 1999, 1512 pp. DNA quantity was estimated by spectrophotometry (NanoDrop®, Thermo scientific, Wilmington, USA), and DNA concentration was adjusted to 50 ng µL-1 for all samples. BOX-PCR was performed using the primer BOX A1R (5′-CTACGGCAAGGCGACG-3′), while REP-PCR was performed using the primers REP-1 (5′-IIIICGICGICATCIGGC-3′) and REP-2 (5′-ICGICTTATCIGGCCTAC-3′) according to Versalovic et al.1010 Versalovic J, Schneider M, De Bruijin FJ, Lupski JR. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol Cell Biol. 1994;5:25-40.
The BOX-PCR reaction was performed in a final volume of 15 µL, containing 2.9 µL of milli-Q water, 7.5 µL of 2× QIAGEN Multiplex PCR Master Mix (3 mM Mg2+), 1.6 µL of primer BOX A1R (50 pmol µL-1) and 3 µL of DNA template (50 ng µL-1). The REP-PCR reaction was performed in a final volume of 15 µL, containing 2.0 µL of milli-Q water, 7.5 µL of 2× QIAGEN Multiplex PCR Master Mix (3 mM Mg2+), 1.25 µL of each primer REP 1 and REP 2 (10 pmol µL-1) and 3 µL of DNA template (10 ng µL-1).
The amplification program was designed according to Kaschuk et al.1111 Kaschuk G, Hungria M, Andrade DS, Campo RJ. Genetic diversity of rhizobia associated with common bean (Phaseolus vulgaris L.) grown under the no-tillage and conventional systems in Southern Brazil. Appl Soil Ecol. 2006;32:210-220. PCR amplification consisted of an initial denaturing step (95 °C; 7 min); followed by 35 cycles of denaturation (94 °C; 1 min), annealing (55 °C; 1 min for BOX-PCR and 40 °C; 1 min for REP-PCR) and extension (65 °C; 8 min); followed by a final extension cycle (65 °C; 15 min). The PCR program was performed in a thermocycler Biocycler® (Applied Biosystems).
PCR products were subjected to electrophoresis on an agarose gel 1% (50 V; 7 h) in TAE buffer 0.75×1212 Sambrook J, Fritschi EF, Maniatis T. Molecular Cloning: A Laboratory Manual. New York: Cold Spring Harbor Laboratory Press; 1989. using 1 kb DNA Ladder® (Norgen) as a DNA band position marker. The agarose gel was stained with SYBR® green (Life Technologies) and visualized with a MultiDoc-it® system.
Symbiotic efficiency under greenhouse conditions
Based on genotypic characterization, 30 isolates were selected to evaluate their symbiotic efficiency. In addition to the isolates, the treatments were composed of two R. tropici strains (CIAT899 and H12), one R. freirei strain (PRF81), two nitrogen fertilized treatments (NT1 = 60 and NT2 = 120 kg ha-1 of N) and one control treatment (CT - without inoculation and without N).
Seeds of common bean cv. Pérola were sown in autoclaved Leonard jars in a random block design with three replicates. At five days after emergence (DAE), plantlets were inoculated with a cell suspension containing 1 × 109 cell mL-1 of each isolate and reference strain. Once a week, 200 mL of nutritive solution without N were added.1313 Franco AA, Döbereiner J. Especificidade hospedeira na simbiose com Rhizobium feijão e influência de diferentes nutrientes. Pesq Agropec Bras. 1967;2:467-474. To the nitrogen fertilized treatments NT1 and NT2, 1 and 2 mL, respectively, of a solution containing 106.68 mg mL-1 of urea were added.
Plants were harvested at 35 DAE. Roots were carefully washed, dried in a paper towel, and the nodules were detached and counted to determine the number of nodules (NN). The leaves were detached from shoots to determine the leaf area (LA) using a leaf area meter LI-COR model 3100. Shoots and nodules were dried (65 °C; 72 h) to determine the shoot dry mass (SDM) and nodule dry mass (NDM). Subsequently, to determine SDM, shoot of plants were milled to determine the total N (N-Total) using the Kjedahl method, as described by Silva and Queiroz.1414 Silva DJ, Queiroz AC. Análise de alimentos: métodos químicos e biológicos. 3.ed. Viçosa, MG: UFV; 2006, 235 pp.
16S rRNA sequencing analysis
Based on the symbiotic efficiency five isolates (ALSG5A1, JPrG1A1, JPrG6A8, JPrG8A7 and PCG4A2) were selected for 16S rRNA sequencing. The DNA of the isolates was obtained according to Laranjo et al.1515 Laranjo M, Machado J, Young JPW, Oliveira S. High diversity of chickpea Mesorhizobium species isolated in a Portuguese agricultural region. FEMS Microbiol Ecol. 2004;48(1):101-107. The 16S rRNA region was amplified by PCR reaction using the primers Y1 and Y3.1515 Laranjo M, Machado J, Young JPW, Oliveira S. High diversity of chickpea Mesorhizobium species isolated in a Portuguese agricultural region. FEMS Microbiol Ecol. 2004;48(1):101-107.,1616 Young JPW, Downer HL, Eardly BD. Phylogeny of the phototrophic Rhizobium strain BTAi1 by polymerase chain reaction-based sequencing of a 16S rRNA gene segment. J Bacteriol. 1991;173:2271-2277. The amplicons were purified and used on the sequencing reaction according to Laranjo et al.1515 Laranjo M, Machado J, Young JPW, Oliveira S. High diversity of chickpea Mesorhizobium species isolated in a Portuguese agricultural region. FEMS Microbiol Ecol. 2004;48(1):101-107. Sequences coding for the partial 16S rRNA genes of the isolates ALSG5A1, JPrG1A1, JPrG6A8, JPrG8A7 and PCG4A2 were obtained and when submitted to the GenBank database (www.ncbi.nlm.nih.gov) received the accession numbers KU598665, KU598663, KU598662, KU598664 and KU598661, respectively.
Statistical analyses
CSU, TST and genotypic characterization data were transformed into a binary matrix. The binary matrices were used for the construction of a similarity matrix using the Jaccard coefficient. The UPGMA (Unweighted pair-group method) was applied to transform the similarity matrix into a similarity dendrogram using NTSYSpc® software.1717 Rolf J. NTSYS-PC: Numerical Taxonomy and Multivariate Analysis System, Version 1.4. New York: Exeter; 1988.
Data obtained from the greenhouse experiment were subjected to analysis of variance; when F was significant, the Scott Knott test of means was applied with a 5% probability using SISVAR statistical software.1818 Ferreira DF. Sisvar: a computer statistical analysis system. Ci Agrotecnol. 2011;35:1039-1042. Pearson correlation analyses were also performed for NDM and SDM, LA and N-Total using R statistical software.1919 Wessa P. Pearson Correlation (v1.0.6) in Free Statistics Software (v1.1.23-r7). Office for Research Development and Education; 2014. Available at: www.wessa.net/rwasp_correlation.wasp/. Accessed 10.01.14.
www.wessa.net/rwasp_correlation.wasp/...
For the 16S rRNA-based phylogeny, the sequences obtained were submitted to NCBI BLAST against a non-redundant nucleotide database for getting homologous sequences.2020 Altschul SF, Gish W, Miller M, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403-410. Sequences showing degree of similarity were aligned using the CLUSTAL W program.2121 Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;25:4876-4882. The evolutionary history was inferred using the Maximum-likelihood method,2222 Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993;10:512-526. with tree consensus being inferred from 500 replicates using bootstrap.2323 Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783-791. The evolutionary distances were computed using the Maximum Composite Likelihood method proposed by Tamura et al.2424 Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci. 2004;101:11030-11035. All positions containing gaps and missing data were eliminated. There were a total of 989 positions in the final dataset. Evolutionary analyses were conducted in MEGA6.2525 Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30:2725-2729.
Results
CSU and TST clustering analysis
CSU evaluation revealed the distribution of 98 isolates among 28 groups, while for TST the isolates were distributed into 27 groups. Among the reference strains, R. tropici CIAT899 grew in all carbon sources, showing the same reference of CSU observed to the 24 isolates (Table S1). Regarding TST, R. freirei PRF81 showed greater tolerance among the reference strains; however, 18 isolates were able to grow under more stringent conditions as compared to R. freirei PRF81 (Table S2).
Similarity analysis based on the CSU and TST data resulted in similarities among isolates varying from 60% to 100%, and five physiological groups (PG) were formed considering a similarity level of 75% (Fig. 1). PG1 comprises the reference strain R. freirei PRF81 and three isolates, representing 3% of the total isolates. The reference strains R. tropici CIAT899 and R. tropici H12 clustered with 49 isolates on PG4, representing 48.5% of the evaluated isolates. Moreover, 38, 7 and 1 isolates formed PG2, PG3 and PG5 clusters, respectively (Fig. 1).
Consensus dendrogram obtained by combining CSU and TST data of 98 isolates from wild genotypes of common bean. The dendrogram was generated using the UPGMA algorithm, and the similarity matrix was determined using the Jaccard coefficient.
Genotypic characterization based on BOX and REP-PCR markers
Based on the results of physiological clustering (Fig. 1), 55 isolates were selected for genotypic characterization with the REP- and BOX-PCR markers. Using this approach, isolates were distributed among 11 different genotype groups (GG) when considering a 65% similarity (Fig. 2).
Consensus dendrogram obtained by combining the REP and BOX-PCR data of 55 isolates from wild genotypes of common bean. The dendrogram was generated using the UPGMA algorithm, and the similarity matrix was determined using the Jaccard coefficient.
Fingerprinting analysis based on BOX- and REP-PCR markers showed that most of the isolates (65.45%) clustered with the R. tropici reference strains R. tropici CIAT899 and R. tropici H12, forming GG1 with an approximately 65% similarity. The isolates JPrG4A9, UbALG7A7 and PCG4A2 showed an approximately 69% similarity with the reference strain R. freirei PRF81 (GG3). Moreover, the isolate ALSG9A8 showed an approximately 72% similarity with the strain of R. etli bv. phaseoli CFN 42 (GG2). Even with a large fraction of isolates clustering with R. tropici reference strains, no instances of identical fingerprinting were observed among the bacteria studied, indicating high polymorphism among the isolates.
Symbiotic efficiency under greenhouse conditions
To evaluate symbiotic efficiency in greenhouse conditions, 30 isolates were used based on genotype clustering (Fig. 2). The inoculation significantly affected all of the evaluated parameters (Table 1).
Nodule number (NN - nº plant-1), nodule dry mass (NDM - mg plant-1), shoot dry mass (SDM - g plant-1), leaf area (LA - cm2 plant-1) and total nitrogen (N-Total - g kg plant-1) of common bean inoculated with different rhizobia isolates.
The NN showed significant difference among the isolates. The Significant greater values varied from 133 to 174 nodules plant-1 and, it were found for the isolates ALSG5A4, JPrG6A8, NVSG11A3, UnPaG11A9 and UnPaG8A12. Among the reference strains, the best results were observed for R. tropici CIAT899 and R. tropici H12, with 110 and 96 nodules plant-1, respectively. Those same isolates also showed high NDM values; however, there was no significant difference between the reference strains R. tropici CIAT899 and R. tropici H12. Moreover, 17 isolates showed greater NDM than the R. freirei PRF81 (Table 1).
Regarding SDM, greater values were observed for the treatment corresponding to 120 kg ha-1 of N (NT2). However, the isolates JPrG6A8, NVSG11A3, NVSG2A2, NVSG7A7, PCG2A5, UbALG3A5, UbALG4A6 and UnPaG8A12 showed values of SDM similar to those obtained by the reference strains R. tropici CIAT899 and R. tropici H12 and by the treatment corresponding to 60 kg ha-1 of N (NT1) (Table 1).
The effect of the treatments on LA was similar to those on SDM, except for the isolates JPrG6A8, UbALG3A5 and UbALG4A6, which had lower values of LA compared to the reference strains R. tropici CIAT899 and R. tropici H12. Additionally, the isolates NVSG2A6 and UnPaG11A9 showed values equal to those observed for the reference strains R. tropici CIAT899 and R. tropici H12 (Table 1).
Similar to SDM and LA, higher N-Total values were observed for NT1 and NT2 and between inoculated treatments. Among the inoculated treatments, the isolates NVSG2A2 and UnPaG8A12 had N-Total values that were statistically similar to that observed for the reference strain R. tropici CIAT899 (Table 1).
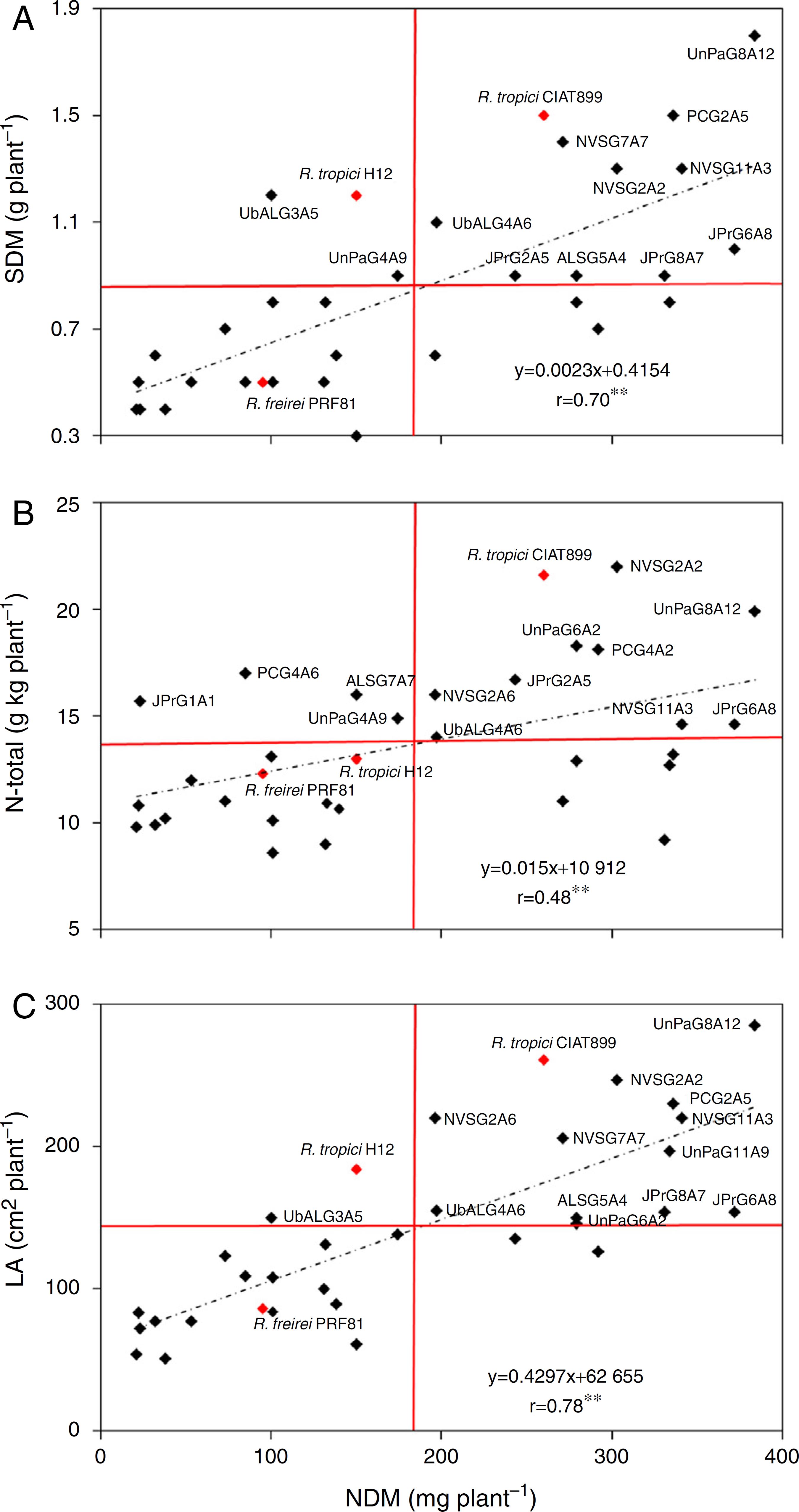
Correlation analysis of NDM with parameters of shoot growth (SDM, N-Total and LA) allowed for the identification of different classes of symbiotic efficiencies, such as few nodulation/few growth, few nodulation/high growth, high nodulation/few growth and high nodulation/high growth. The distribution of the isolates occurred in different quarters of the graphic defined in function of the general mean of the treatments (Fig. 3).
Pearson correlation analysis for (NDM) nodule dry mass and (SDM) shoot dry mass. (A) NDM and (N-Total) total nitrogen, (B) NDM and (LA) leaf area, and (C) common bean inoculated with different rhizobia isolates. Dashed lines represent the correlation tendency, and red lines represent the general mean for each parameter. ** p < 0.01.
Correlation analyses between NDM × SDM, NDM × N-Total and NDM × LA showed highly significant results, with correlation coefficients (r) of 0.7, 0.48 and 0.78, respectively. This analysis showed that for the three correlations performed, the isolates UnPaG8A12, NVSG11A3, NVSG2A2, JPrG6A8 and UbALG4A6 and, the reference strain R. tropici CIAT899, were above the general mean (Fig. 3). Remarkably the isolates UbALG3A5 and UnPaG4A9 (Fig. 3A), JPrG1A1, PCG4A6, ALSG7A7 and UnPaG4A9 (Fig. 3B) and UbALG3A5 (Fig. 3C) also showed high values of SDW, N-Total and LA, respectively, even with low values NDM.
Regarding the correlation between NDM and SDM, 10 isolates and the reference strain R. tropici CIAT899 appeared on the upper right quarter, indicating that these bacteria show a high capacity for both nodulation and accumulation of SDM. In contrast, the isolates UbALG3A5 and UnPaG4A9, and the reference strain R. tropici H12, accumulated a high SDM, even with few nodulations (Fig. 3A). The isolates and the reference strain R. freirei PRF81 appearing in the lower left quarter had a symbiotic performance below the general mean, and for those in the lower right quarter, the vegetal growth was weak, even with high nodulation. For the NDM × N-Total correlation, nine isolates and the reference strain R. tropici CIAT899 showed a high symbiotic efficiency, promoting high values for both NDM and N-Total. In addition, the isolates JPrG1A1, PCG4A6, ALSG7A7 and UnPaG4A9 were able to promote high N-Total accumulation with low NDM (Fig. 3B).
Regarding the NDM and LA correlation, twelve isolates and the reference strain R. tropici CIAT899 appeared on the upper right quarter, indicating that these bacteria show high capacity for both nodulation and LA growth, and the isolate UbALG3A5 and the reference strain R. tropici H12 promoted high LA with low NDM (Fig. 3C).
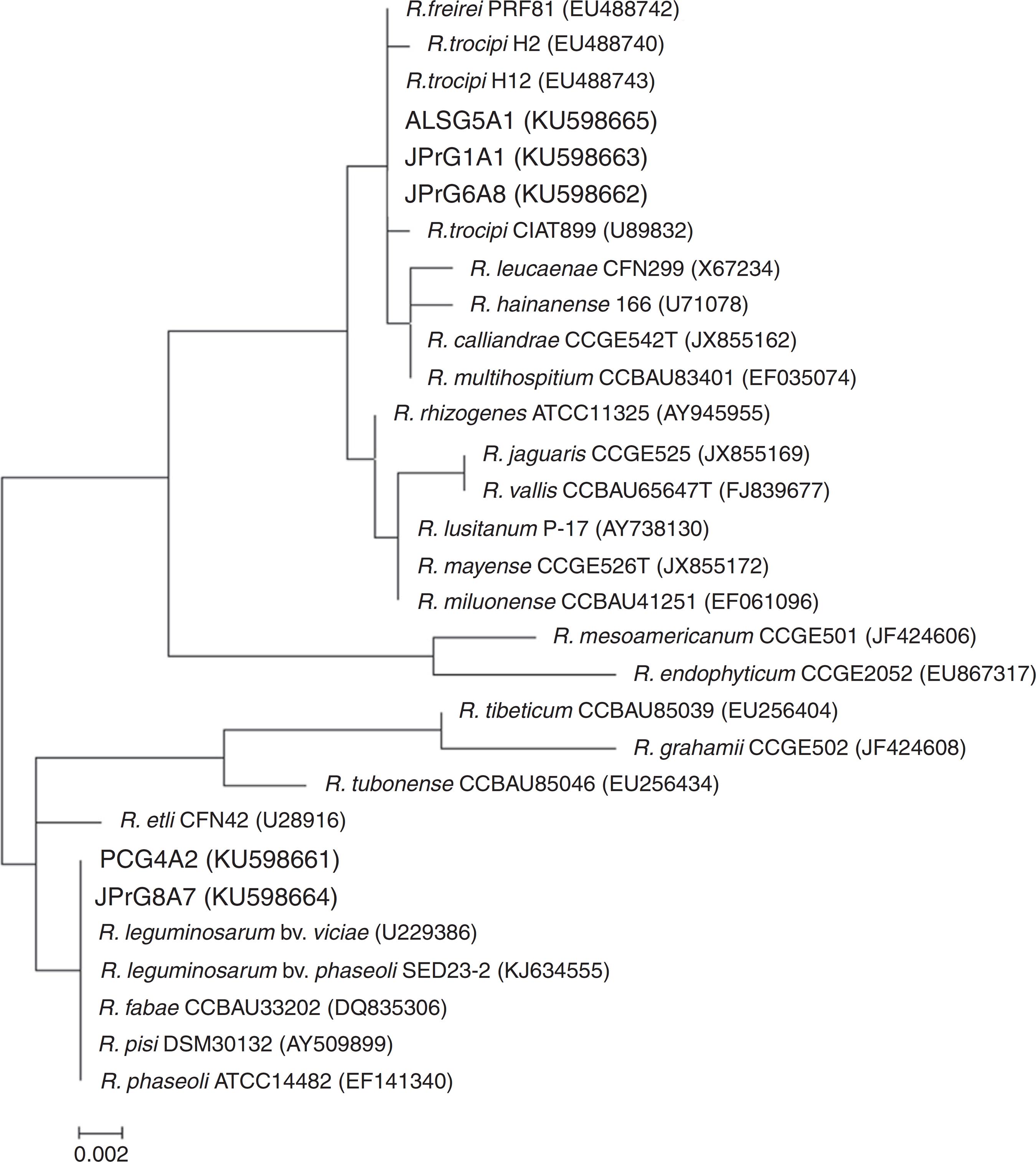
Sequencing analysis revealed the grouping of three isolates (ALSG5A1, JPRG1A1 and JPrG6A8) into the R. tropici and R. freirei cluster, while the other two isolates grouped with Rhizobium leguminosarum bv. viciae, R. leguminosarum bv. phaseoli, Rhizobium fabae, Rhizobium pisi and Rhizobium phaseoli strains (Fig. 4).
Maximum-likelihood phylogeny of the 16S rRNA gene showing the relationships between rhizobia isolates obtained from wild genotypes of common bean (in bold) and Rhizobium sp. reference strains. GenBank accession numbers are shown in parentheses. Bar, 2 nt substitutions per 1000 nt.
Discussion
The clustering analysis based on CSU and TST showed that the Rhizobium reference strains clustered with 52 isolates, forming the groups PG1 and PG4. Isolates grouped in these two PGs accounted for more than 50% of the isolates evaluated (Fig. 1).
The reference strain R. freirei PRF81 and the isolates NVSG7A7, ALSG10A8 and NVSG8A2 were grouped in the PG1 cluster and showed a high TST. However, these bacteria showed weak CSU (Table S2). High TST may be associated with a specific genetic control, conferring such ability to these bacteria. Gomes et al.2626 Gomes DF, Batista JSS, Schiavon AL, Andrade DS, Hungria M. Proteomic profiling of Rhizobium tropici PRF 81: identification of conserved and specific responses to heat stress. BMC Microbiol. 2012;12:1-12. showed that the PRF 81 (SEMIA 4080) strain is capable of differentially expressing some conserved heat-responsive proteins, such as DnaK and GroEL. They also reported the up-regulation of proteins involved in a variety of metabolic pathways, including translation factors and oxidative stress-responsive proteins, indicating that the responses of R. freirei strain PRF 81 to heat stress go beyond the induction of heat shock proteins.
Reference strains R. tropici CIAT899 and R. tropici H12 formed PG4 together with 48.5% of the evaluated isolates. These bacteria were not able to grow under temperatures of 43 and 48 °C; however, they grew until 2% NaCl content on the other tested temperatures. Moreover, these bacteria showed greater CSU than the bacteria of the PG1 (Table S1). These results corroborate those of Dall’Agnol et al.,66 Dall’Agnol RF, Ribeiro RA, Ormeño-Orrillo E, et al. Rhizobium freirei sp. nov., a symbiont of Phaseolus vulgaris that is very effective at fixing nitrogen. Int J Syst Evol Microbiol. 2013;63:4167-4173. who reported a greater capacity of R. tropici CIAT899 to use C source as compared to R. freirei PRF81.
In our work, R. tropici CIAT899 and R. tropici H12 were able to grow on culture medium containing glucose and glycerol. According to Castellane and Lemos,2727 Castellane TCL, Lemos EGM. Composição de exopolissacarídeos produzidos por estirpes de rizóbios cultivados em diferentes fontes de carbono. Pesq Agrop Bras. 2007;42:1503-1506. these C sources favor growth and exopolysaccharide (EPS) production in these strains. EPS production is related to the process of rhizobia adaptation to limiting environmental conditions,2828 Bomfeti CA, Florentino LA, Guimarães AP, Cardoso PG, Guerreiro MC, Moreira FMS. Exopolysaccharides produced by the symbiotic nitrogen-fixing bacteria of leguminosae. Rev Bras Ci Solo. 2011;35:657-671. allowing these bacteria to grow under conditions of high temperature and soil salinity. Thus, the evaluation of temperature and salinity tolerance is a key step in the selection process of rhizobia because such factors can inhibit their growth.66 Dall’Agnol RF, Ribeiro RA, Ormeño-Orrillo E, et al. Rhizobium freirei sp. nov., a symbiont of Phaseolus vulgaris that is very effective at fixing nitrogen. Int J Syst Evol Microbiol. 2013;63:4167-4173.,2929 Peter W, John RP, Kumar P, Thomas S. Tannin acyl hydrolase production by Citrobacter sp. isolated from tannin rich environmental, using Tamarindus indica seed powder. J Appl Sci Environ Manag. 2009;13:95-97.,3030 Elboutahiri N, Thami-Alami I, Udupa SM. Phenotypic and genetic diversity in Sinorhizobium meliloti and S. medicae from drought and salt affected regions of Morocco. BMC Microbiol. 2010;10:1-13.
On the fingerprinting analysis based on BOX- and REP-PCR markers the highest similarity (95.5%) was identified between the isolates NVSG11A3 and UnPaG7A3 (GG1). These results corroborate the work of Grange,3131 Grange L [M.Sc. Dissertation] Diversidade de rizóbio capaz de nodular o feijoeiro (Phaseolus vulgaris) isolado de solos da Região Nordeste e da Região Sul do Brasil. Londrina, Brasil: Universidade Estadual de Londrina; 2001, 87 pp. who reported high diversity among rhizobia isolates obtained from common bean nodules cultivated in Cerrado soils.
These markers are used to characterize and determine genotypic differences among bacterial strains to describe new species3232 Han TX, Han LL, Wu LJ, et al. Mesorhizobium gobiense sp. nov. and Mesorhizobium tarimense sp. nov., isolated from wild legumes growing in desert soils of Xinjiang, China. Int J Syst Evol Microbiol. 2008;58:2610-2618.
33 Lu YL, Chen WF, Wang ET, et al. Genetic diversity and biogeography of rhizobia associated with Caragana species in three ecological regions of China. Syst Appl Microbiol. 2009;32:351-361.-3434 Koedoeboecz L, Halbritter A, Mogyorossy T, Kesckes ML. Phenotypic and genotypic diversity of rhizobia in cropping areas under intensive and organic agriculture in Hungary. Eur J Soil Biol. 2009;45:394-399. and identifying the rhizobia strains used to compose the Brazilian commercial inoculants for common bean.3535 MAPA (Ministério da Agricultura, Pecuária e Abastecimento). Instrução Normativa Nº 30, de 12 de novembro de 2010; 2010. Available at: http://extranet.agricultura.gov.br/sislegis/. Accessed 13.06.11.
http://extranet.agricultura.gov.br/sisle...
Among the 30 isolates evaluated under greenhouse conditions, the isolates JPrG6A8, NVSG11A3, UnPaG8A12, UnPaG11A9 and ALSG5A4 produced higher values of NN than the reference strains and, higher NDM than the reference strain R. freirei PRF81. Although NN is not a determining factor for the efficiency of BNF,3636 Hungria M, Bohrer TRJ. Variability of nodulation and dinitrogen fixation capacity among soybean cultivars. Biol Fertil Soils. 2000;31:45-52. it may be indicative of the high nodulation efficiency of bacteria. Moreover, NN and NDM are measurements frequently used as indicators of nodulation.3737 Araújo FF, Carmona FG, Tiritan CS, Creste JE. Fixação biológica de N2 no feijoeiro submetido a dosagens de inoculante e tratamento químico na semente comparado à adubação nitrogenada. Act Sci Agron. 2007;29:535-540.
When NN and NDM were evaluated simultaneously, verified that 80% of the isolates had better or equal results compared to those of the R. tropici and R. freirei reference strains. Because NDM shows a better correlation with symbiotic performance,3636 Hungria M, Bohrer TRJ. Variability of nodulation and dinitrogen fixation capacity among soybean cultivars. Biol Fertil Soils. 2000;31:45-52. our results demonstrated that 17 isolates produced similar amounts of NDM compared to the reference strains R. tropici CIAT899 and R. tropici H12, indicating the high symbiotic efficiency of these isolates.
Results of nodulation had a direct effect on the accumulation of dry matter on the shoots of the plants. The treatments with higher NN and NDM also showed high values of SDM and LA. Among the inoculated treatments, greater values of SDM and LA were found for the reference strains R. tropici CIAT899 and R. tropici H12 and for the isolates NVSG11A3, NVSG2A2, NVSG7A7, PCG2A5 and UnPaG8A12. Measurements of SDM and LA provide important information about plant growth3838 Antunes JEL, Gomes RLF, Lopes ACA, Araújo ASF, Lyra MCCP, Figueiredo MVB. Eficiência simbiótica de isolados de rizóbio noduladores de feijão-fava (Phaseolus lunatus L.). Rev Bras Ci Solo. 2011;35:751-757. because these parameters are good indicators of plant nutritional status, which has a direct influence on crop production.3939 Xavier GR, Martins LMV, Rumjanek NG, Neves MCP. Tolerância de rizóbio de feijão-caupi à salinidade e à temperatura em condição in vitro. Rev Caatinga. 2007;20:1-9.,4040 Souza RA, Hungria M, Franchini JC, Maciel CD, Campo RJ, Zaia DAM. Conjunto mínimo de parâmetros para avaliação da microbiota do solo e da fixação biológica do nitrogênio pela soja. Pesq Agropec Bras. 2008;43:83-91. Moreover, this approach has been used for strain selection to compose bacterial inoculants.4040 Souza RA, Hungria M, Franchini JC, Maciel CD, Campo RJ, Zaia DAM. Conjunto mínimo de parâmetros para avaliação da microbiota do solo e da fixação biológica do nitrogênio pela soja. Pesq Agropec Bras. 2008;43:83-91.,4141 Gonzáles TO, Campanharo JC, Lemos EGM. Genetic characterization and nitrogen fixation capacity of Rhizobium strains on common bean. Pesq Agropec Bras. 2008;43:1177-1184.
The nitrogen content of plants (N-Total) was also affected by inoculation; however, only two isolates, NVSG2A2 and UnPaG8A12, showed N-Total content similar to that of the reference strain R. tropici CIAT899. Greater values of N-Total were observed to the nitrogen treatments. Similar results reported by Gonzáles et al.4141 Gonzáles TO, Campanharo JC, Lemos EGM. Genetic characterization and nitrogen fixation capacity of Rhizobium strains on common bean. Pesq Agropec Bras. 2008;43:1177-1184. in a field experiment showed that nitrogen treatment resulted in greater N-Total compared to inoculation.
The use of correlation of NDM with parameters of shoot growth (SDM, N-Total and LA) allows to identify more efficient isolates, which are located at the upper left and upper right quarters (Fig. 3). Interestingly, our results revealed a high symbiotic efficiency of the isolates JPrG6A8, NVSG11A3, NVSG2A2, UbALG4A6 and UnPaG8A12, showing NDM, SDM, N-Total and LA values similar to the best reference strain, R. tropici CIAT899. Moreover, the isolate UnPaG4A9 also had promising results, with high values of SDM and N-Total even under low NDM. These are very interesting features for rhizobial isolates, since it can allow to provide better producing results under field conditions.2626 Gomes DF, Batista JSS, Schiavon AL, Andrade DS, Hungria M. Proteomic profiling of Rhizobium tropici PRF 81: identification of conserved and specific responses to heat stress. BMC Microbiol. 2012;12:1-12.,3131 Grange L [M.Sc. Dissertation] Diversidade de rizóbio capaz de nodular o feijoeiro (Phaseolus vulgaris) isolado de solos da Região Nordeste e da Região Sul do Brasil. Londrina, Brasil: Universidade Estadual de Londrina; 2001, 87 pp.,3434 Koedoeboecz L, Halbritter A, Mogyorossy T, Kesckes ML. Phenotypic and genotypic diversity of rhizobia in cropping areas under intensive and organic agriculture in Hungary. Eur J Soil Biol. 2009;45:394-399.,3939 Xavier GR, Martins LMV, Rumjanek NG, Neves MCP. Tolerância de rizóbio de feijão-caupi à salinidade e à temperatura em condição in vitro. Rev Caatinga. 2007;20:1-9.
Sequencing analysis of the 16S rRNA gene revealed that the isolates ALSG5A1, JPrG1A1 and JPrG6A8 are very closely related with the reference strains of R. tropici CIAT899, R. tropici H12 and R. freirei PRF81, which are used in Brazil as commercial inoculant for common bean4242 Menna P, Hungria M, Barcellos FG, Bangel EV, Hess PN, Martínez-Romero E. Molecular phylogeny based on the 16S rRNA gene of elite rhizobial strains used in Brazilian commercial inoculants. Syst Appl Microbiol. 2006;29:315-332.; however, the isolate JPrG1A1 showed low efficiency in the greenhouse experiment (Table 1).
Considering that many studies have discussed the importance of investigating the efficiency of biological nitrogen fixation in the selection of isolates for the development of inoculants,4343 Carvalho FG, Selbach PA, Bizarro MJ. Eficiência e competitividade de variantes espontâneos isolados de estirpes de Bradyrhizobium spp. recomendadas para a cultura da soja (Glycine max). Rev Bras Ci Solo. 2005;29:883-891.,4444 Melloni R, Moreira FMS, Nobrega RSA, Siqueira JO. Eficiência e diversidade fenotípica de bactérias diazotróficas que nodulam caupi (Vigna unguiculata (L.) Walp) e feijoeiro (Phaseolus vulgaris L.) em solos de mineração de bauxita em reabilitação. Rev Bras Ci Solo. 2006;30:235-246. the isolates ALSG5A1 and JPrG6A8 must be tested for their agronomic efficiency under field conditions, aiming to state their potential use as inoculant for common bean.
Conclusions
Forty-five percent of the isolates evaluated and the reference strain R. freirei PRF81 show high tolerance to temperature, while 24% of the isolates and the reference strain R. tropici CIAT899 are able to use all of the carbon sources studied. Clustering analysis based on physiological parameters group 50% of the isolates, R. tropici CIAT899 and R. tropici H12 with a similarity level of 76%. REP- and BOX-PCR markers group about 65% of the isolates, R. tropici CIAT899 and R. tropici H12 with a similarity level of 66%. About 20% of the isolates show symbiotic efficiency similar to or better than the best Rhizobium reference strain (CIAT899). By the phylogenetic analysis of the 16S rRNA the isolates ALSG5A1 and JPrG6A8 belong to the group of strains used as commercial inoculant for common bean in Brazil.
Acknowledgments
The authors thank the National Council for Scientific and Technological Development (CNPq) and the Brazilian Agricultural Research Corporation (Embrapa) by the technical and financial support.
Appendix A Supplementary data
Supplementary data associated with this article can be found, in the online version, at doi:10.1016/j.bjm.2016.09.002.
References
-
1Brougthon WJ, Hernández G, Blair M, Beebe S, Gepts P, Vardeyleyden J. Bean (Phaseolus ssp.): model food legume. Plant Soil 2003;252:55-128.
-
2FAO – Food and Agricultural Organization of the United Nations; 2015. Available at: http://faostat3.fao.org/download/Q/QC/E Accessed 19.05.15.
» http://faostat3.fao.org/download/Q/QC/E -
3Acompanhamento de safra brasileira: grãos, décimo levantamento Brasília: CONAB – Companhia Nacional de Abastecimento; 2013. Available at: http://www.conab.gov.br/OlalaCMS/uploads/arquivos/13_07_09_09_04_53_boletim_graos_junho_2013.pdf Accessed 09.02.14.
» http://www.conab.gov.br/OlalaCMS/uploads/arquivos/13_07_09_09_04_53_boletim_graos_junho_2013.pdf -
4Soares BL [D.Sc. Thesis] Avaliação técnica econômica do feijoeiro-comum inoculado com rizóbio em diferentes ambientes Lavras, Brasil: Universidade Federal de Lavras; 2012, 150 pp.
-
5Novais RF, Alvarez VVH, Barros NF, Fontes RLF, Cantarutti RB, Neves JCL. Fertilidade do Solo Viçosa: Sociedade Brasileira de Ciência do Solo; 2007, 1017 pp.
-
6Dall’Agnol RF, Ribeiro RA, Ormeño-Orrillo E, et al. Rhizobium freirei sp. nov., a symbiont of Phaseolus vulgaris that is very effective at fixing nitrogen. Int J Syst Evol Microbiol 2013;63:4167-4173.
-
7Mhamdi R, Ardley J, Tian R, et al. High-quality permanent draft genome sequence of Ensifer meliloti strain 4H41, an effective salt- and drought-tolerant microsymbiont of Phaseolus vulgaris. Stand Genomic Sci 2015;10:1-7.
-
8Sampaio FB [M.Sc. Dissertation] Isolados de rizóbios capturados de genótipos silvestres de feijoeiro: obtenção, morfologia e uso de fontes de carbono. Goiânia, Brasil: Universidade Federal de Goiás; 2013, 85 pp.
-
9Ausubel F, Brent R, Kingston R, et al. Short Protocols in Molecular Biology 4th ed. New York: John Wiley and Sons, Inc.; 1999, 1512 pp.
-
10Versalovic J, Schneider M, De Bruijin FJ, Lupski JR. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol Cell Biol 1994;5:25-40.
-
11Kaschuk G, Hungria M, Andrade DS, Campo RJ. Genetic diversity of rhizobia associated with common bean (Phaseolus vulgaris L.) grown under the no-tillage and conventional systems in Southern Brazil. Appl Soil Ecol 2006;32:210-220.
-
12Sambrook J, Fritschi EF, Maniatis T. Molecular Cloning: A Laboratory Manual New York: Cold Spring Harbor Laboratory Press; 1989.
-
13Franco AA, Döbereiner J. Especificidade hospedeira na simbiose com Rhizobium feijão e influência de diferentes nutrientes. Pesq Agropec Bras 1967;2:467-474.
-
14Silva DJ, Queiroz AC. Análise de alimentos: métodos químicos e biológicos 3.ed. Viçosa, MG: UFV; 2006, 235 pp.
-
15Laranjo M, Machado J, Young JPW, Oliveira S. High diversity of chickpea Mesorhizobium species isolated in a Portuguese agricultural region. FEMS Microbiol Ecol 2004;48(1):101-107.
-
16Young JPW, Downer HL, Eardly BD. Phylogeny of the phototrophic Rhizobium strain BTAi1 by polymerase chain reaction-based sequencing of a 16S rRNA gene segment. J Bacteriol 1991;173:2271-2277.
-
17Rolf J. NTSYS-PC: Numerical Taxonomy and Multivariate Analysis System, Version 1.4 New York: Exeter; 1988.
-
18Ferreira DF. Sisvar: a computer statistical analysis system. Ci Agrotecnol 2011;35:1039-1042.
-
19Wessa P. Pearson Correlation (v1.0.6) in Free Statistics Software (v1.1.23-r7) Office for Research Development and Education; 2014. Available at: www.wessa.net/rwasp_correlation.wasp/ Accessed 10.01.14.
» www.wessa.net/rwasp_correlation.wasp/ -
20Altschul SF, Gish W, Miller M, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol 1990;215:403-410.
-
21Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997;25:4876-4882.
-
22Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 1993;10:512-526.
-
23Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985;39:783-791.
-
24Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci. 2004;101:11030-11035.
-
25Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30:2725-2729.
-
26Gomes DF, Batista JSS, Schiavon AL, Andrade DS, Hungria M. Proteomic profiling of Rhizobium tropici PRF 81: identification of conserved and specific responses to heat stress. BMC Microbiol. 2012;12:1-12.
-
27Castellane TCL, Lemos EGM. Composição de exopolissacarídeos produzidos por estirpes de rizóbios cultivados em diferentes fontes de carbono. Pesq Agrop Bras. 2007;42:1503-1506.
-
28Bomfeti CA, Florentino LA, Guimarães AP, Cardoso PG, Guerreiro MC, Moreira FMS. Exopolysaccharides produced by the symbiotic nitrogen-fixing bacteria of leguminosae. Rev Bras Ci Solo 2011;35:657-671.
-
29Peter W, John RP, Kumar P, Thomas S. Tannin acyl hydrolase production by Citrobacter sp. isolated from tannin rich environmental, using Tamarindus indica seed powder. J Appl Sci Environ Manag. 2009;13:95-97.
-
30Elboutahiri N, Thami-Alami I, Udupa SM. Phenotypic and genetic diversity in Sinorhizobium meliloti and S. medicae from drought and salt affected regions of Morocco. BMC Microbiol 2010;10:1-13.
-
31Grange L [M.Sc. Dissertation] Diversidade de rizóbio capaz de nodular o feijoeiro (Phaseolus vulgaris) isolado de solos da Região Nordeste e da Região Sul do Brasil Londrina, Brasil: Universidade Estadual de Londrina; 2001, 87 pp.
-
32Han TX, Han LL, Wu LJ, et al. Mesorhizobium gobiense sp. nov. and Mesorhizobium tarimense sp. nov., isolated from wild legumes growing in desert soils of Xinjiang, China. Int J Syst Evol Microbiol. 2008;58:2610-2618.
-
33Lu YL, Chen WF, Wang ET, et al. Genetic diversity and biogeography of rhizobia associated with Caragana species in three ecological regions of China. Syst Appl Microbiol 2009;32:351-361.
-
34Koedoeboecz L, Halbritter A, Mogyorossy T, Kesckes ML. Phenotypic and genotypic diversity of rhizobia in cropping areas under intensive and organic agriculture in Hungary. Eur J Soil Biol 2009;45:394-399.
-
35MAPA (Ministério da Agricultura, Pecuária e Abastecimento). Instrução Normativa Nº 30, de 12 de novembro de 2010; 2010. Available at: http://extranet.agricultura.gov.br/sislegis/ Accessed 13.06.11.
» http://extranet.agricultura.gov.br/sislegis/ -
36Hungria M, Bohrer TRJ. Variability of nodulation and dinitrogen fixation capacity among soybean cultivars. Biol Fertil Soils. 2000;31:45-52.
-
37Araújo FF, Carmona FG, Tiritan CS, Creste JE. Fixação biológica de N2 no feijoeiro submetido a dosagens de inoculante e tratamento químico na semente comparado à adubação nitrogenada. Act Sci Agron 2007;29:535-540.
-
38Antunes JEL, Gomes RLF, Lopes ACA, Araújo ASF, Lyra MCCP, Figueiredo MVB. Eficiência simbiótica de isolados de rizóbio noduladores de feijão-fava (Phaseolus lunatus L.). Rev Bras Ci Solo 2011;35:751-757.
-
39Xavier GR, Martins LMV, Rumjanek NG, Neves MCP. Tolerância de rizóbio de feijão-caupi à salinidade e à temperatura em condição in vitro. Rev Caatinga 2007;20:1-9.
-
40Souza RA, Hungria M, Franchini JC, Maciel CD, Campo RJ, Zaia DAM. Conjunto mínimo de parâmetros para avaliação da microbiota do solo e da fixação biológica do nitrogênio pela soja. Pesq Agropec Bras 2008;43:83-91.
-
41Gonzáles TO, Campanharo JC, Lemos EGM. Genetic characterization and nitrogen fixation capacity of Rhizobium strains on common bean. Pesq Agropec Bras. 2008;43:1177-1184.
-
42Menna P, Hungria M, Barcellos FG, Bangel EV, Hess PN, Martínez-Romero E. Molecular phylogeny based on the 16S rRNA gene of elite rhizobial strains used in Brazilian commercial inoculants. Syst Appl Microbiol 2006;29:315-332.
-
43Carvalho FG, Selbach PA, Bizarro MJ. Eficiência e competitividade de variantes espontâneos isolados de estirpes de Bradyrhizobium spp. recomendadas para a cultura da soja (Glycine max). Rev Bras Ci Solo. 2005;29:883-891.
-
44Melloni R, Moreira FMS, Nobrega RSA, Siqueira JO. Eficiência e diversidade fenotípica de bactérias diazotróficas que nodulam caupi (Vigna unguiculata (L.) Walp) e feijoeiro (Phaseolus vulgaris L.) em solos de mineração de bauxita em reabilitação. Rev Bras Ci Solo. 2006;30:235-246.
Publication Dates
-
Publication in this collection
Jan-Mar 2017
History
-
Received
25 June 2015 -
Accepted
20 May 2016