ABSTRACT
A bacterium isolated from Sterkfontein dam was confirmed to produce bioflocculant with excellent flocculation activity. The 16S rDNA nucleotide sequence analyses revealed the bacteria to have 99% similarity to Streptomyces platensis strain HBUM174787 and the sequence was deposited in the Genbank as Streptomyces platensis with accession number FJ 486385.1. Culture conditions for optimal production of the bioflocculant included glucose as a sole carbon source, resulting in flocculating activity of 90%. Other optimal conditions included: peptone as nitrogen source; presence of Mg2+ as cations and inoculum size of 1.0% (v/v) at neutral pH of 7. Optimum dose of the purified bioflocculant for the clarification of 4 g/L kaolin clay suspension at neutral pH was 0.2 mg/mL. Energy Dispersive X-ray analysis confirmed elemental composition of the purified bioflocculant in mass proportion (%w/w): carbon (21.41), oxygen (35.59), sulphur (26.16), nitrogen (0.62) and potassium (7.48). Fourier Transform Infrared Spectroscopy (FTIR) indicated the presence of hydroxyl, carboxyl, methoxyl and amino group in the bioflocculant. The bioflocculant produced by S. platensis removed chemical oxygen demand (COD) in river water and meat processing wastewater at efficiencies of 63.1 and 46.6% respectively and reduced their turbidity by 84.3 and 75.6% respectively. The high flocculating rate and removal efficiencies displayed by S. platensis suggests its industrial application in wastewater treatment.
Keywords:
Culture conditions; Polysaccharide; Flocculating activity; Kaolin suspension; EDX & FTIR
Introduction
Flocculation is a process whereby suspended particles agglomerate to form a bigger flocs. Flocculating agents are widely employed in industrial processes such as potable water purification, wastewater treatment, downstream processing, food and fermentation industries.11 Guo J, Jing Y, Xin X, et al. Characterization and flocculation mechanism of a bioflocculant from hydrolyzate of rice stover. Bioresour Technol. 2015;177:393-397.,22 Salehizadeh H, Yan N. Recent advances in extracellular biopolymer flocculants. Biotechnol Adv. 2014;32:1506-1522. Generally, flocculants are categorized into (i) inorganic flocculants, such as aluminium sulfate and polyaluminium chloride, (ii) organic synthetic flocculants, such as polyacrylamide derivatives and polyethylene amine, (iii) naturally occurring flocculants, such as sodium alginate, chitosan and microbial flocculants. Chemically synthesized flocculants have been widely used in flocculation because of their cost effectiveness and flocculating efficiency. However, their usage has resulted in some environmental and health problems associated with Alzheimer's disease.33 Sam S, Kucukasik F, Yenigun O, Nicolaus B, Oner ET, Yukselen MA. Flocculating performances of exopolysaccharides produced by a halophilic bacterial strain cultivated on agro-industrial waste. Bioresour Technol. 2011;102:1788-1794.,44 Li Q, Liu HL, Qi Q, Wang F, Zhang Y. Isolation and characterization of temperature and alkaline stable bioflocculant from Agrobacterium Sp. M503. New Biotechnol. 2010;27:789-794. Therefore, the replacement of chemically synthesized flocculant with an environmentally friendly biodegradable flocculants demands urgent attention. Microbial flocculants are recognized as a biotechnological tool in remediating contaminated water through the process of bioflocculation, which is recognized to be safer and economical compared to common chemical flocculation processes.55 Liu W, Wang K, Li B, Yuan H, Yang J. Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour Technol. 2010;101:1044-1048. Bacteria, fungi, algae and actinomycetes have been reported to be responsible for the production of extracellular polymers such as polysaccharides, functional proteins and glycoproteins.66 Zhao G, Ma F, Wei L, Chua H. Using rice straw fermentation liquor to produce bioflocculants during an anaerobic dry fermentation process. Bioresour Technol. 2012;113:83-88. Bioflocculant producing microorganisms have been isolated from freshwater environment, activated sludge, soil, brewery wastewater and marine sediment.77 Ahmad HRA, Azni I, Hamid HR, Al-joubory Y, Uemura BSU, Ibn A. Flocculation behavior and mechanism of bioflocculant produced by Aspergillus flavus. J Environ Manage. 2015;150:466-471.–99 Anthony U, Sekelwa C, Leonard M, Olubukola O, Farhad A, Anthony O. Thermostable bacterial bioflocculant produced by Cobetia Spp. isolated from Algoa Bay (South Africa). Int J Environ Res Pub Health. 2012;9(6):2108-2120.
Streptomycetaceae is a family of Actinobacteria, which constitute the monotypic suborder Streptomycineae. Streptomyces are gram positive, aerobic, spore forming microorganism and they are found majorly in soil sediment and marine environment. The presence of linear chromosomes which is responsible for exchange of information is peculiar to this bacteria.1010 Kämpfer P. The family Streptomycetaceae, Part I: Taxonomy. In: Dworkin M, Falkow S, Rosenberg E, Schleifer K, Stackebrandt E, eds. The Prokaryotes. 2006:538. According to literature, Streptomyces platensis has found biotechnological application due to the production of a novel anti-migrating agent called migrastatin. In addition, the antibiotics that are produced by this organism have the ability to target specific enzymes.1111 Reymond S, Janine C. Migrastatin and analogues: new anti-metastatic agents. Compt Rend Chim. 2010;11:1447-1462. Actinomycetes producing bioflocculants have been implicated in flocculation. However, there is no report in literature confirming the industrial application of this strain in the treatment of wastewaters. Hence, in this paper we evaluated the culture conditions and establish the application of bioflocculant produced by S. platensis in river water and wastewater treatment.
Materials and methods
Source and collection of sample
The Sterkfontein Dam (28º24'31.2″ S 029º02'15.0″ E.) is located just outside the town of Harrismith, in the Eastern Free State Province of South Africa. The Dam is part of the Tugela-Vaal Water Project located on the Nuwejaarspruit, a tributary of the Wilge River in the upper catchment area of the Vaal River.
Isolation of bioflocculant-producing microorganism
The water sample was collected from the dam aseptically into a sterile air-tight bottle. The bottle was kept in a cooler box containing Ice Pack and was transported to the laboratory for processing. Serial dilutions were done and cultivation of actinomycetes from the processed water sample was done according to the method described by Jensen and co-workers1212 Jensen PR, Dwight R, Fenical W. Distribution of actinomycetes in near shoretropical marine sediments. Appl Environ Microbiol. 1991;57:1102-1108. using Yeast Malt Extract agar (YMA) supplemented with 50 mg L-1 cyclohexamide and 20 mg L-1 nalidixic acid to inhibit the growth of fungi and bacteria respectively. The composition of the media includes 0.4% yeast extract, 1% malt extract, 0.4% glucose and 1.6% of bacteriological agar in distilled water. An aliquot of 100 µL of the sample was spread over the cultivation medium and incubated at 28 ºC for one to two weeks. After the incubation period, typical colonies were purified by streaking on fresh YMA plates and purified colonies were stored on YMA slants in 20% glycerol at -80 ºC in the freezer.
Screening and culturing of bioflocculant-producing microorganisms
Total number of fifteen isolated actinomycetes strains were screened for bioflocculant production in accordance with the protocol of Xia and co-workers1313 Xia S, Zhang Z, Wang X, et al. Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresour Technol. 2008;99:6520-6527. with adaptation. The isolates were inoculated into McCartney bottles containing 5 mL of screening medium and incubated for 48 h on a rotatory shaker at 30 ºC and 160 rpm. The composition of the screening medium was 2% glucose, 0.05% urea, 0.05% yeast extract, 0.02% (NH4)2 SO4, 0.01% NaCl, 0.02% MgSO4·7H2O, 0.5% K2HPO4 and 0.2% KH2PO4 in distilled water. This pre-culture procedure was then used as the standard inocula preparation for subsequent experiment. A McCartney bottle containing 5 mL production medium was inoculated with 100 µL pre-culture of the strain and incubated at 25 ºC in a rotatory shaker (Inco shake, Labotec) at 130 rpm for 48 h. The production medium composition include 1% glucose, 0.1% peptone, 0.03% MgSO4·7H2O, 0.5% K2HPO4 and 0.2% KH2PO4 in distilled water. The initial pH of the medium was adjusted to 7.0 with NaOH (0.1 M) and HCl (0.1 M). After incubation, cell free culture supernatants were used to determine bioflocculant production against kaolin clay suspension. Finally, the strain that displayed highest flocculating efficiency was selected for further investigation.
Determination of flocculating activity
Kaolin clay suspension was used as test material in assessing the efficiency of bioflocculant production. Kaolin suspension was prepared by dissolving 4 g of kaolin clay in 1 L distilled water. A volume of 100 mL of this kaolin suspension was added to 2 mL of cell-free supernatant and 3 mL of calcium chloride (1%, w/v) in 250 mL conical flask. The mixture was thoroughly shaken for 30 s and gently poured into 100 mL measuring cylinder and allowed to stand for 5 min. The optical density (OD) of the clarifying solution was measured using a spectrophotometer UV/Visible Biowave II and Biowave II+, England at 550 nm. The control experiment was done in the same way, but cell free supernatant was replaced with 2 mL of production medium. The flocculating activity (FA) was calculated using the equation;
where A and B are the respective absorbance of the sample and control experiment at 550 nm.
Identification of organism
The genomic DNA of actinobacteria was extracted using a ZR fungal/bacteria DNA preparation kit according to the manufacturer's instructions. The 16S rRNA gene was amplified by using universal primers 27f: (5'-GAGTTTGATCCTGGCTCAG-3') 1492r: (5'-GGTTACCTTGTTACGACT-3').1414 Lane DJ. 16/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M, eds. Nucleic Acid Techniques in Bacterial Systematics. Chichester/New York/Brisbane/Toronto/Singapore: John Wiley and Sons; 1991:115–175. PCR amplification was carried out in 20 µL reaction volume containing 10 µL of Econo PCR master mix, 1 mM of each primer, 1 µL of template DNA. Sterile 8 µL distilled PCR grade water was added to a final volume of 20 µL. PCR programme used was an initial denaturation (94 ºC for 5 min), 45 cycles of denaturation (94 ºC for 30 s), annealing (55 ºC for 30 s), extension (72 ºC for 1 min, 30 s) and final extension for (72 ºC for 10 min). Thermoscientific thermal cycler PCR machine was used to run the reaction. PCR products was electrophoresed on 1% agarose gel in 1× Tris Borate EDTA buffer stained with gel red and was visualized under UV transilluminator to confirm that a fragment of the correct size had been amplified. Automated sequencing of 16S rRNA genes of the organism was done using the sequencer AB 35,100 × L genetic analyzer with 24 capillaries. Sequencing reactions was performed according to the manufacturer's protocol using Big Dye version 3.1 dye terminator cycle sequencing kit (Applied Biosystems) with 27f and 1492r primers. Sequences was aligned in the Gene bank database using the BLASTN program at the National Centre for Biotechnology information (NCBI) and percent homology score was obtained to identify the organism.
Extraction and purification of bioflocculant
The purification was done according to the method employed by Chen1515 Chen H, Zhang JF, Jiang PJ, Yang SL, Liu ZL. Composition and characterization of microbiological flocculant SC06. Environ Chem. 2002;21:360-364. and Piyo1616 Piyo N, Cosa S, Mabinya LV, Okoh AI. Assessment of bioflocculant production by Bacillus sp. Gilbert, a marine bacterium isolated from the bottom sediment of Algoa Bay. Mar Drugs. 2011;9:1232-1242. with modification. After 72 h of fermentation, the culture broth was centrifuged at 8000 × g for 30 min to remove bacteria cells. One volume of sterile distilled water was added to the supernatant phase and centrifuged at 8000 × g for 15 min to remove insoluble substances. Two volumes of ethanol were later added to the supernatant, stirred and left to stand for 12 h at 4 ºC. The precipitate was vacuum-dried to obtain crude bioflocculant. The crude product was dissolved in water to yield a solution, to which one volume of a mixed solution of chloroform and n-butyl alcohol (5:2, v/v) was added. The mixture was stirred, poured into separating funnel and allowed to stand for 12 h at room temperature. Finally, the supernatant was discarded and two volumes of ethanol were added to recover the precipitate and then lyophilized to obtain a purified bioflocculant.
Effect of cultivation parameters on bioflocculant production by S. platensis
Initial pH, Cations, carbon and nitrogen sources, inoculum size and thermal stability were the parameters investigated for optimum bioflocculant production in our test isolate.
Effect of initial pH (3–12) of the production medium on bioflocculant production were assessed at acidic, basic and neural medium. Monovalent, divalent and trivalent cations sources (Na+, K+, Mg2+, Mn2+, Al3+, Fe3+ and Ca2+) were also investigated for optimum bioflocculant production. To determine the effect of carbon and nitrogen sources on bioflocculant production, organic and inorganic sources were evaluated. Glucose as an organic carbon source was used to replace xylose, maltose, lactose, sodium acetate, sodium carbonate and phthalate which were representative of organic and inorganic carbon sources respectively. Ammonium chloride as an inorganic nitrogen source was used to replace yeast extract, peptone, tryptone and casein which serves as organic nitrogen source. The influence of inoculum size on bioflocculant production by our test organism was tested by varying the inoculum size from 0.5 to 2.0 mL (v/v). This was done by inoculating 0.5 mL, 1.0 mL, 1.5 mL and 2.0 mL of pre-cultured bacteria cultivated at 28 ºC with continuous agitation at 160 rpm for 72 h in 50 mL production medium. Heat stability was evaluated at a temperature range of 50, 60, 70, 80, 90 and 100 ºC for 25 min and the flocculating activity was determined using the protocol of Gong and co-workers.1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674.
Jar test determination of bioflocculant dosage
The optimum dose of purified bioflocculant needed for clarification of kaolin clay suspension (4 g/L) was determined using the Jar test experiment following a standard protocol. Different concentrations of powdered purified bioflocculant ranging from 0.1 to 1.0 mg/mL were prepared and their flocculating activities were measured. The standard procedure includes rapid mixing at 160 rpm for 2 min, followed by gradual flocculation period of 40 rpm for 2 min and sedimentation for 5 min. After sedimentation, 2 mL was gently withdrawn from the upper clarifying phase in order to measure the flocculating activity.1818 Lee SH, Shin WS, Shin MC, Choi SJ, Park LS. Improvement of water treatment performance by using polyamine flocculants. Environ Technol. 2001;22:653-659.,1919 Wang Y, Gao BY, Yue QY, Wei JC, Zhou WZ, Gu R. Color removal from textile industry wastewater using composite flocculants. Environ Technol. 2010;28(6):629-637.
Comparism of traditional flocculants and the purified bioflocculant
The flocculating efficiencies of polyacrylamide, polyethylenime, alum, ferric chloride and the test bioflocculant on kaolin clay suspension were investigated according to the protocol of.2020 Ugbenyen MA, Okoh AI. Characteristics of a bioflocculant produced by a consortium of Cobetia and Bacillus species and its application in the treatment of wastewaters. Water SA. 2014;40(1):. In order to compare the coagulation performance of synthesized flocculants and the purified bioflocculant, each flocculant was prepared at a different concentration with the addition of cations that stimulates flocculation processes and tested against kaolin suspension using the jar test. The control experiment was prepared in the same way but the flocculants (chemically synthesized or bioflocculant) was replaced with distilled water. The residual flocculating activity were measured as earlier discussed.
Flocculation of wastewater and river water
The meat processing wastewater was collected from an Industrial area situated on the Gauteng East Rand and the river water was collected from Wilge River in the Eastern Free State of South Africa. Meat processing wastewater and river water were collected aseptically using sterile 1 L glass bottle, labelled correctly and transported into the laboratory for processing using cooler box containing ice packs. Samples were processed immediately on arrival in the laboratory. Optimum dose of 0.2 mg/mL of the purified bioflocculant and 2 mL of 1% MgCl2 were added into 200 mL of river water and meat processing wastewater. The mixture was agitated at 160 rpm for 2 min using Jar test and the speed was later reduced to 40 rpm for 2 min to facilitate floc formation. Afterwards, it was allowed to settle for 5 min and the supernatant was gently taken for analysis. The residual COD and turbidity for each sample were determined according to the method of Gong and co-workers.1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674. To assay for chemical oxygen demand (COD), 2 mL of raw and treated samples were added into COD vials (150 mg L-1). For the blank, 2 mL of sterile distilled water was added into the COD vial. The caps were tightly closed, rinsed with water and finally wipe with a clean paper towel. The DRB 200 digester was pre heated to 150 ºC before inserting the prepared COD vials in the digester. The vials were gently inverted to mix and were inserted into the digester and were heated for 2 h. The COD vials were removed after digesting and were allowed to cool down at room temperature. After cooling, the samples were measured using a spectrophotometer (DR 3800). Turbidity of the river water and meat processing wastewater were measured using turbidimeter (HACH, USA). The removal efficiency was calculated as follows:
where A 0 is the initial value and A is the value after the flocculation treatment.
Chemical analysis and characterization of purified bioflocculant
The total protein content of purified bioflocculant of S. platensis was determined by Lowry's method using bovine serum albumin (BSA) as a standard.2121 Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin Phenol reagent. J Biol Chem. 1951;193:265-275. Lowry solution which is the composition of A [2.85 g NaOH & 14.3 g Na2CO3], B [1.42 g CuSO4·5H2O] and C [2.85 g Na2Tartarate·2 (H2O)] were prepared separately in distilled water and the solutions were mixed together in a ratio (vol:vol) of 100:1:1. BSA standard was prepared by weighing 0.05 g of BSA in 500 mL distilled water, stirred and final concentration of the stock was adjusted to 100 mg BSA L-1. The stock BSA solution and dilutions were used to prepare the standard curve. The sample was prepared in triplicates the same way the standard was done, well mixed and 0.5 mL was transferred into a glass tube. After, 0.7 mL of Lowry solution was added, vortex and incubated for 20 min at room temperature in the dark. A volume of 0.1 mL of diluted folin reagent were later added to each tube, vortex and incubated for 30 min at room temperature in the dark. The sample was later transferred into cuvette and the absorbance of the sample and the standard were measured at absorbance of 750 nm. The calibration curve from the absorbance reading of the standard was prepared and the protein content of the sample in mg BSA L-1 was calculated from the curve. Total sugar content of bioflocculant was determined by phenol sulfuric method using glucose as a standard solution.2222 Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28:350-356. Working standard solution was prepared by dissolving 100 mg of glucose in 100 mL of distilled water and 10 mL was taken from the stock and further diluted to 100 mL. The standard was prepared by taking 0, 0.2, 0.4, 0.6, 0.8 and 1 mL of the working standard into a series of test tubes and 0 serves as the blank. The volume was made up to 1 mL in all tubes including the sample tubes by adding distilled water. A volume of 1 mL of phenol solution was added to each tube and 5 mL of 96% sulphuric acid was later added and shaken thoroughly. The contents in the tubes were later shaken after 10 min and placed in water bath at 25–30 ºC for 20 min. The absorbance was measured at absorbance of 490 nm and the amount of total carbohydrate present in the sample solution was calculated from the standard graph. The functional groups of purified bioflocculant were analyzed using Fourier Transform Infrared Spectrophotometer (Perkin Elmer System 2000, England). The bioflocculant was ground with potassium bromide salt at 25 ºC and pressed into a pellet for FTIR spectroscopy at the frequency range of 4000–370 cm-1. Thermo-gravimetric analysis was done using a TGA analyzer (STA 449/C Jupiter, Netzsch, Germany Perkin Elmer TGA7 Thermo gravimetric Analyzer, USA) over a temperature range of 20–900 ºC at a heating rate of 10 ºC/min under a constant flow of nitrogen gas.
The purified bioflocculant sample placed on a carbon coated stub were gold coated in a gold coating chamber, using Eiko IB3 ION coater, afterwards, the scanning electron microscopic image of the bioflocculant was obtained using a JEOL JSM-7800F FE-SEM with an Oxford SDD X-Max EDS System which was used to analyze the elemental composition of the purified bioflocculant.
Statistical analysis
Results were expressed as means ± standard deviation of three replicates and were subjected to one way analysis of variance (ANOVA) followed by Duncan multiple range tests to determine significant differences in all the parameters using SPSS 16.0. Values were considered statistically.
Results
Bioflocculant yield and chemical analysis of the purified bioflocculant
Following 72 h of fermentation under optimal culture conditions and composition, 4.61 g of pure purified bioflocculant was obtained from 1 L of culture broth. The flocculant secreted by S. platensis had polysaccharide (83%) and (4.6%) protein as its major constituents.
Identification of bioflocculant producing actinomycetes
Fifteen strains of actinomyces isolated from water sample, collected from Sterkfontein dam producing bioflocculant were selected on the basis of flocculating activity exceeding 70%. Among the selected strains, SFD 20 exhibited the highest flocculating activity of over 90% and was used for further studies. The 16S rDNA PCR yielded a product of expected size (approximately 1.5 kb). Basic Local Alignment Search Tool (BLAST) analysis of the nucleotide sequence of the 16S rDNA revealed the bacteria to have 99% similarity to Streptomyces platensis strain HBUM174787 and the sequence was deposited in the Genbank as S. platensis with accession number FJ 486385.1.
Impact of carbon and nitrogen sources
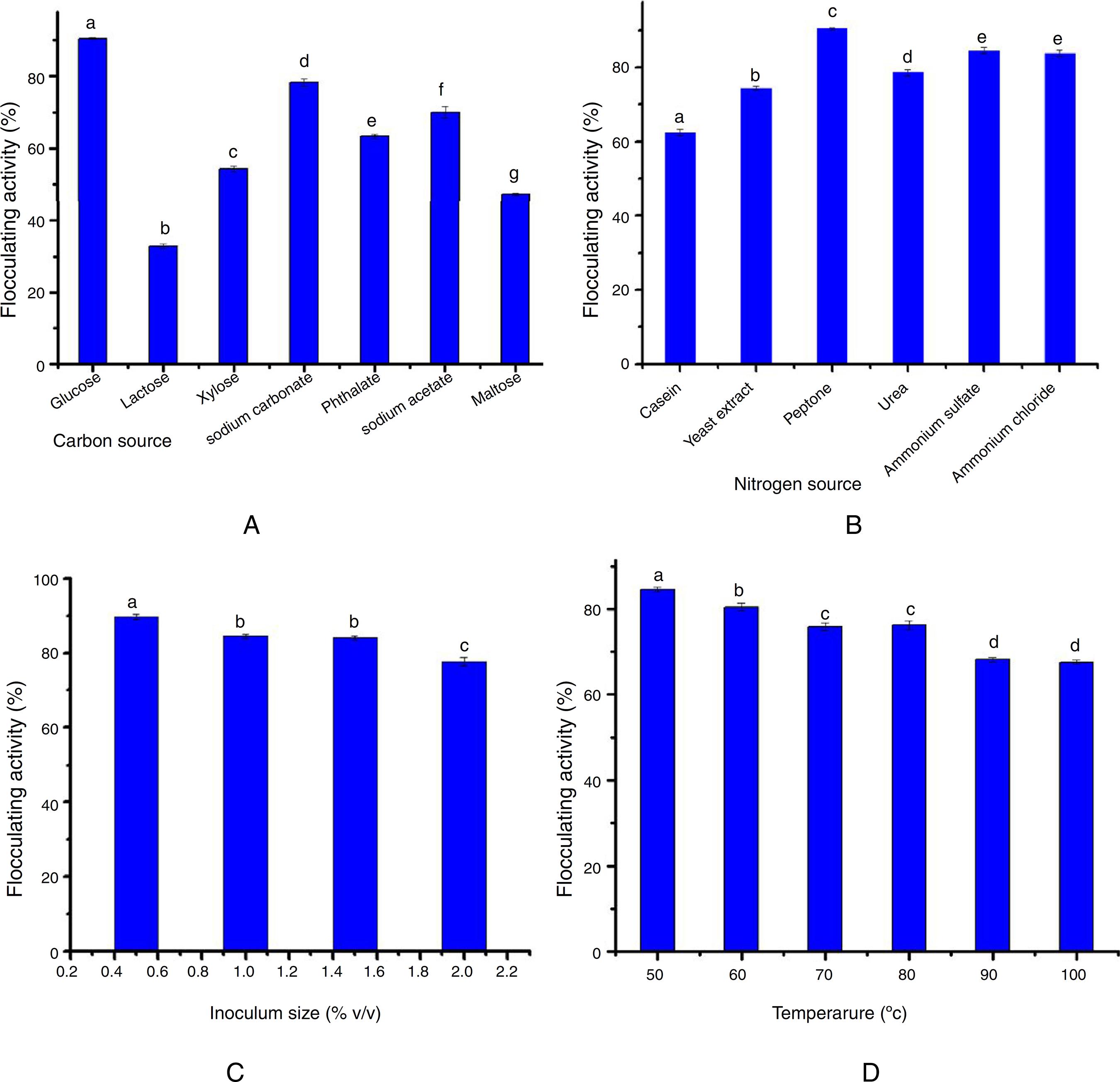
Fig. 1A reveals the bioflocculant production potential of S. platensis utilizing different carbon sources. Glucose, sodium carbonate, phthalate and sodium acetate appeared to support cell growth and bioflocculant production at a flocculating activity of over 60%, with glucose showing the maximum activity (90%) as a carbon source. On the other hand, maltose, xylose and lactose were poorly utilized by the bioflocculant production of the test organism.
Effect of various sources (A) carbon sources, (B) nitrogen sources (C) inoculum size, and (D) temperature on the flocculating activity of Streptomyces platensis. Percentage flocculating activities with different alphabetic letters are significantly different p < 0.05.
Data obtained with respect to the impact of nitrogen sources on bioflocculant production by the test organism in this study revealed that the organic (yeast extract and peptone), and the inorganic (urea, ammonium sulfate and ammonium chloride) nitrogen sources supported the growth and bioflocculant production with peptone eliciting the most prominent flocculating activity (90%) (Fig. 1B).
Impact of pH and cations on bioflocculant production by S. platensis
The effect of pH of the medium (at the range of 3–12 scale) on bioflocculant production is shown in (Fig. 2A). Our results showed that the optimum bioflocculant production was observed at the neutral pH 7. It is noteworthy that the basic medium supported the growth of the organism at an average flocculating activity of 55% compared to the weakly displayed activity at the acidic medium (Fig. 2A).
Effect of (A) pH on bioflocculant production, and (B) concentration on flocculating activity of purified bioflocculant produced by Streptomyces platensis. Percentage flocculating activities with different alphabetic letters are significantly different (p < 0.05).
The results of the effect of metal ions on the flocculating activity of S. platensis are presented in Table 1. Of the cations investigated in this study, the divalent cations appeared to support bioflocculant production with flocculating activity of over 74% (74.57–91.05%) and optimum activity of 90% was observed when MgCl2 was utilized. On the other hand, the monovalent ions partially supports flocculating activity exceeding 64% (64.74–71.15%). Also, Al3+ as a representative of trivalent cation enhanced the flocculating activity at 83% while, Fe3+ resulted in a weak flocculating activity of 58% as shown in Table 1.
Effect of cations on flocculating activity. Percentage flocculating activities with different alphabetic letters are significantly different (p < 0.05).
Inoculum size and stability
The data on the effect of inoculum size on bioflocculant production by S. platensis revealed that the optimum flocculating activity was attained at 1% (v/v) inoculum size. Further increase in the inoculum size led to a steady decrease in the flocculating activity (Fig. 2C).
Fig. 2D shows how thermally stable and the influence of heat treatment on the bioflocculant activity. Although, a temperature-dependent decrease was observed following the heat treatment, the flocculating activity of the bioflocculant culture broth was maintained at around 70% over the entire treatment conditions (50–100 ºC) for 25 min.
Jar test determination of bioflocculant dosage
Fig. 2B illustrates the coagulant dosage ranging from 0.1 to 1.0 mg/mL. Optimum flocculating activity of 94.88% was recorded when 0.2 mg/mL coagulant dosage was used to flocculate kaolin clay. In contrast, further increase in dosing with the bioflocculant from 0.3 mg/mL to 1.0 mg/mL resulted in decreased removal of turbidity and flocculating activity.
Comparison of efficiency of chemical flocculants and the purified bioflocculant
The flocculation efficiency of conventional flocculants (polyaluminium chloride, polyethylemine, ferric chloride and alum) and the purified bioflocculant against kaolin clay suspension at a varying concentrations (0.1–1.0 mg/mL) is presented in Table 2. The results revealed that the purified bioflocculant was significantly efficient at the optimum concentration (0.2 mg/mL) when compared with 0.3, 0.8 and 1.0 mg/mL for polyethylemine, alum and ferric chloride, respectively. Polyacrylamide as an inorganic flocculant was able to flocculate kaolin clay suspension at an optimum concentration of 0.1 mg/mL and there was no significant difference (p > 0.05) in its flocculating activity when compared to the test bioflocculant (Table 2).
Application of bioflocculant in the treatment of river water and wastewater
The physico-chemical properties of untreated river and meat processing waste water are shown in Tables 3 and 4 presents the flocculating efficiency of the purified bioflocculant on river and waste water. It was observed that the bioflocculant could flocculate the river water better than the waste water with efficiency, COD removal, and turbidity removal values of 91.4%, 63.1% and 84%, respectively when compared with 82%, 46.6%, and 75.6% for the waste water at 0.2 mg/mL. However, the bioflocculant could remove suspended solids in meat processing waste water better than river water at percentage removal of 72.8% and 60.2% respectively.
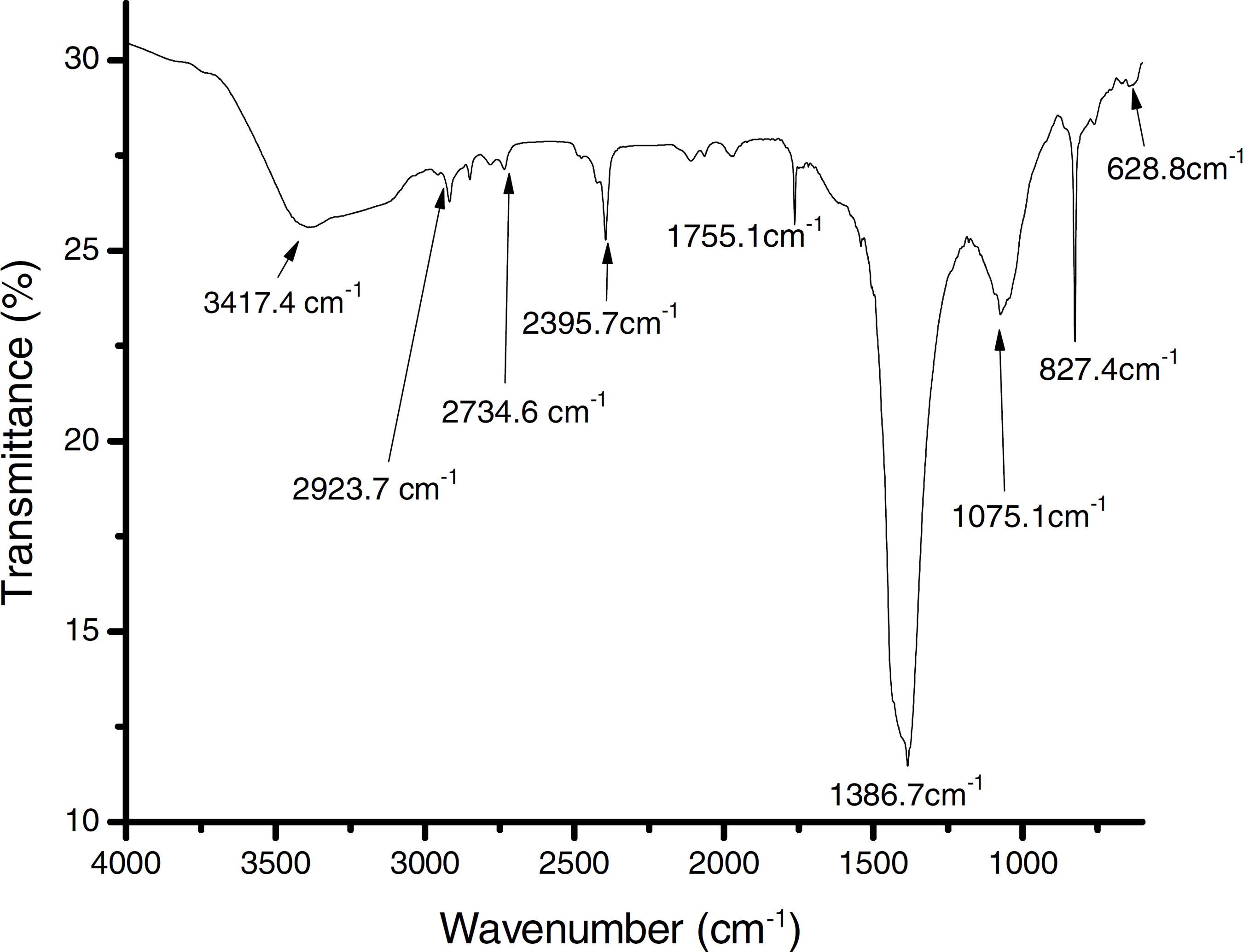
The results of the FTIR, scanning electron microscopy, and the TGA analyses of the purified bioflocculant are presented in (Figs. 3–5) respectively. Peaks corresponding to different functional groups at stretching vibration range of 628.8–3417.4 cm-1 were evident from the FTIR spectrum (Fig. 3). While the purified bioflocculant appeared whitish in nature with an agglomerate of oval shape filled with scattered stains Fig. 4A, the interaction of the bioflocculant and the kaolin clay suspension resulted in the formation of compacted flocs (Fig. 4B). From the TGA analysis, while about 20% loss in weight in the temperature range of 20–150 ºC was observed in the purified bioflocculant, there was a gradual decomposition of the material at temperature range of 150–450 ºC. Over 60% weight was retained after heating the material at 900 ºC (Fig. 5). Further elemental analysis by EDX indicated C – 21.41; O – 35.59; N – 0.62; S – 26.16; and P – 7.48%, w/w as the major constituents of the purified bioflocculant.
Fourier-transform infrared (FTIR) spectroscopy of purified bioflocculant produced by Streptomyces platensis.
Scanning electron micrograph of (A) purified bioflocculant, (B) purified bioflocculant flocculating kaolin suspension, (C) Kaolin powder suspension.
Discussion
The higher yielded purified bioflocculant suggested a higher productivity in bioflocculant produced by our test organism within a short period of time as compared with other strains that has been reported in literature. In contrast to our findings, 3.8 g L-1 of biopolymer was produced by Ochrobactrum ciceri W2,2323 Wang L, Ma F, Lee DJ, Wang AJ, Ren NQ. Bioflocculants from hydrolysates of corn stover using isolated strain Ochrobactium cicero W2. Bioresour Technol. 2013;145:259-263. while 2.43 g of purified bioflocculant was recovered from the culture broth of Virgibacillus sp.2424 Cosa S, Ugbenyen MA, Mabinya LV, Okoh IA. Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr J Microbiol Res. 2013;7(23):2925-2938. The flocculant secreted by S. platensis was mainly polysaccharide (83%) and (4.6%) protein.
The culture medium composition and conditions have been reported to have impact on bioflocculants production.1313 Xia S, Zhang Z, Wang X, et al. Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresour Technol. 2008;99:6520-6527. Optimization of carbon and nitrogen sources could result into attaining maximum flocculating activity within the shortest time of incubation.2525 Xiong Y, Wang Y, Yu Y, Li Q, Wang H, Chen R. Production and characterization of a novel bioflocculant from Bacillus licheniformis. Appl Environ Microbiol. 2010;76:2778-2782. In this study, the highest activity displayed by glucose as a carbon source was the reason for its choice for subsequent experiments. This finding is consistent with the report of1313 Xia S, Zhang Z, Wang X, et al. Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresour Technol. 2008;99:6520-6527.,2424 Cosa S, Ugbenyen MA, Mabinya LV, Okoh IA. Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr J Microbiol Res. 2013;7(23):2925-2938.,2626 Ugbenyen MA, Okoh AI. Flocculating properties of a bioflocculant produced by Bacillus sp. isolated from a marine environment in South Africa. Chem Biochem Eng. 2013;27(4):511-518. where glucose was found favourable for bioflocculant production by Proteus mirabilis TJ-1, Bacillus sp. and Virgibacillus sp. respectively. Conversely, other researchers have reported that sucrose favoured bioflocculant production by Aspergillus flavus, while fructose and glycerol inhibited the flocculating activity.2727 Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour Technol. 2013;127:489-493.
Similarly, the highest flocculating activity observed with utilization of organic nitrogen source (peptone) by our test organism could be attributed to the availability of vitamins and trace elements.2828 Sanjukta S, Neha B, Mihirjyoti P, et al. Bioflocculant production and biosorption of zinc and lead by a novel bacterial species, Achromobacter sp. TERI-IASST N, isolated from oil refinery waste. Chemosphere. 2014;113:116-124. Studies have reported that certain bioflocculant producing strains have effectively utilized peptone or yeast extract as sources of nitrogen, thereby validating peptone as one of the best and cost effective nitrogen sources.44 Li Q, Liu HL, Qi Q, Wang F, Zhang Y. Isolation and characterization of temperature and alkaline stable bioflocculant from Agrobacterium Sp. M503. New Biotechnol. 2010;27:789-794.,2727 Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour Technol. 2013;127:489-493. Carbon and nitrogen source requirement differs with different strains. For instance, bioflocculant produced by Bacillus subtilis F9 and A. flavus both utilized sucrose and peptone as their optimum carbon and nitrogen sources respectively.2727 Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour Technol. 2013;127:489-493.,2929 Giri SS, Harshiny M, Sen SS, Sukumaran V, Park SC. Production and characterization of a thermostable bioflocculant from Bacillus subtilis F9, isolated from wastewater sludge. Ecotoxicol Environ Saf. 2015;121:45-50. The presence of amino, hydroxyl, carboxyl and carbonyl groups in microbial flocculants which are complex polymers, reiterates the need for the availability of complex carbon and nitrogen sources.2525 Xiong Y, Wang Y, Yu Y, Li Q, Wang H, Chen R. Production and characterization of a novel bioflocculant from Bacillus licheniformis. Appl Environ Microbiol. 2010;76:2778-2782.,3030 Salehizadeh H, Shojaosadati SA. Removal of metal ions from aqueous solution by polysaccharide produced from Bacillus firmus. Water Res. 2003;37:4231-4235.
The effect of initial pH of the medium on bioflocculant production is shown in Fig. 2A. Xia1313 Xia S, Zhang Z, Wang X, et al. Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresour Technol. 2008;99:6520-6527. reported that initial pH of the culture medium influences the electric charge and the oxidation reduction potential of the cells, which can also affect nutrients absorption and enzymatic reactions. The effect of pH was investigated at the range of 3–12 and optimum bioflocculant production was observed at the neutral pH 7, which reduces the quantity of acids and alkali needed to adjust pH. However, basic medium also seems to support the growth at flocculating activity of over 60% compared to weak flocculating activity observed at acidic medium. The poor flocculating activity at the acidic medium could be attributed to excessive concentration of hydrogen ions that alters the electric charge. This finding corroborates the report of Giri2929 Giri SS, Harshiny M, Sen SS, Sukumaran V, Park SC. Production and characterization of a thermostable bioflocculant from Bacillus subtilis F9, isolated from wastewater sludge. Ecotoxicol Environ Saf. 2015;121:45-50. and Gong1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674. where optimum bioflocculant production by B. subtilis F9 and Serratia ficaria was attained at neutral pH. However, the effect of pH conditions on bioflocculant production varies with different strains, operational conditions and composition of the medium.3131 Shu CH, Lung MY. Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia camphorate in batch cultures. Process Biochem. 2004;39:931-937. For instance, pH range of 7–12 favoured bioflocculant production by Bacillus megaterium and maximum activity was attained at pH 9.0.3232 Zheng Y, Ye ZL, Fang XL, Li YH, Cai WM. Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour Technol. 2008;99:7686-7691. In another study, optimum bioflocculant production by Achromobacter sp. was attained at a weak acidic pH 6.2828 Sanjukta S, Neha B, Mihirjyoti P, et al. Bioflocculant production and biosorption of zinc and lead by a novel bacterial species, Achromobacter sp. TERI-IASST N, isolated from oil refinery waste. Chemosphere. 2014;113:116-124. On the other hand, the bioflocculant produced by Virgibacillus sp. with flocculating activity of 85.8% was attained at the pH 10.
Cations enhance the initial adsorption of bioflocculants on to the surface of the suspended particles, thereby minimizing the negative charge on the polymer and the particles.3333 Zhang ZQ, Lin B, Xia SQ, Wang XJ, Yang AM. Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. Environ Sci. 2007;19:667-673. Yim3434 Yim JH, Kim SJ, Ahn SH, Lee HK. Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresour Technol. 2007;98:361-367. reported that contribution of metal ions to the flocculating activity of a bioflocculant could be due to neutralizing and stabilizing the residual charge of functional groups and then forming bridges between particles. The weak flocculating activity of Fe3+ in the present study is consistent with its inhibitory role against the flocculating activity of the flocculant from the test organism. Our submission is in agreement with the report of Okaiyeto3535 Okaiyeto K, Nwodo UU, Mabinya LV, Okoli AS, Okoh AI. Evaluation of flocculating performance of a thermostable bioflocculant produced by marine Bacillus sp. Environ Technol. 2016, http://dx.doi.org/10.1080/09593330.2015.1133717.
http://dx.doi.org/10.1080/09593330.2015....
and Ugbenyen2626 Ugbenyen MA, Okoh AI. Flocculating properties of a bioflocculant produced by Bacillus sp. isolated from a marine environment in South Africa. Chem Biochem Eng. 2013;27(4):511-518. where the flocculating activity of bioflocculant produced by two Bacillus spp. were both inhibited by Fe3+. Furthermore, the significantly higher value shown by the Mg2+ is not only suggestive of its neutralizing and stabilizing effect on the charge of functional groups of the flocculant but is also consistent with previous assertion3636 Nwodo UU, Agunbiade MO, Green E, Mabinya LV, Okoh AI. A freshwater Streptomyces, isolated from Tyume river, produces a predominantly extracellular glycoprotein bioflocculant. Int J Mol Sci. 2012;13:8679-8695. where Mg2+ enhanced the flocculating activity of the bioflocculant produced by Streptomyces sp.
The inoculum size is an important parameter that enhances better flocculating activity of cell growth. Reports have demonstrated optimum and facilitated flocculating activity of the test organism when inocular size of 1% (v/v) are used and this is in agreement with the result of our finding in this study. Specifically,1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674.,3434 Yim JH, Kim SJ, Ahn SH, Lee HK. Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresour Technol. 2007;98:361-367.,3737 Li Z, Zhong S, Lei H, Chen RW, Yu Q, Li HL. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour Technol. 2009;100:3650-3656. have earlier implicated inoculum size of 1% (v/v) in facilitating maximum flocculating activity in bioflocculant produced by S. ficaria, Bacillus licheniformis and for the multi-microorganism consortia containing Staphylococcus sp. and Pseudomonas sp. Contrary to these findings, highest flocculating activity was recorded when inoculum size of 2% (v/v) was used in the bioflocculant production by A. flavus and Brachybacterium sp.2727 Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour Technol. 2013;127:489-493.,3838 Nwodo UU, Agunbiade MO, Green E, Nwamadi M, Rumbold K, Okoh AI. Characterization of an exopolymeric flocculant produced by a Brachybacterium sp. Materials. 2013;6:1237-1254. It has been established that large inoculum size results into overlapping of niches, thereby hindering bioflocculant production.22 Salehizadeh H, Yan N. Recent advances in extracellular biopolymer flocculants. Biotechnol Adv. 2014;32:1506-1522.,3939 Salehizadeh H, Shojaosadati SA. Extracellular biopolymeric flocculants-recent trends and biotechnological importance. Biotechnol Adv. 2001;19:371-385. This is well illustrated in the present study where the best and enhanced activity was observed at the lowest investigated inoculum size contrary to the relatively attenuated activity at the higher inocular sizes.
The significantly displayed heat stable tendency and flocculating activity (70%) by the produced bioflocculant from S. platensis over the different treatment conditions could be attributed to the fact that the main backbone is a polysaccharide. This finding corroborates the report of Giri2929 Giri SS, Harshiny M, Sen SS, Sukumaran V, Park SC. Production and characterization of a thermostable bioflocculant from Bacillus subtilis F9, isolated from wastewater sludge. Ecotoxicol Environ Saf. 2015;121:45-50. whereby the bioflocculant produced by B. subtilis is composed mainly of polysaccharides. Consistent with our report, the consortium of Cobetia sp. and Bacillus sp. retained its residual flocculating activity of 87% after heating at 100 ºC for 25 min.2020 Ugbenyen MA, Okoh AI. Characteristics of a bioflocculant produced by a consortium of Cobetia and Bacillus species and its application in the treatment of wastewaters. Water SA. 2014;40(1):. Likewise, bioflocculant produced by A. flavus was thermostable, retaining 90% flocculating activity within the temperature range of 10–100 ºC.
Optimization of coagulant dosage in the treatment of water is important in order to achieve maximum turbidity removal and flocculating efficiency at a cheaper cost. In this study, the enhanced flocculating activity at 0.2 mg/mL coagulant dosage suggests that the initial dose of 0.1 mg/mL used may be considered insufficient and ultimately caused incomplete and inefficient coagulation. Although, this report is contrary to the findings of Cosa2424 Cosa S, Ugbenyen MA, Mabinya LV, Okoh IA. Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr J Microbiol Res. 2013;7(23):2925-2938. and Wang4040 Yukselen MA, Gregory J. The effect of rapid mixing on the break-up and re-formation of flocs. Chem Technol Biotechnol. 2004;79(7):782-788. where optimum dosage employed to achieve maximum flocculation was 0.1 mg/mL and 12 mg/mL respectively, it has established an optimal dosage regimen (0.2 mg/mL) for the bioflocculant from the test organism. This variation in dosage could be as a result of charge reversal and destabilization of colloidal particles.4141 Wang L, Ma F, Qu Y, Sun D, Li A, Guo J. Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. W J Microbiol Biotechnol. 2011;27:2559-2565. Overdosing or insufficient dose would lead to poor performance of coagulant during the process of flocculation. Hence, validating and establishing the optimum dose needed at a particular time in treatment of portable water and wastewater in order to reduce cost of production, minimize the risk of overdosage in human beings and minimize formation of sludge in water treatment is imperative.4242 Patel H, Vashi RT. Comparison of naturally prepared coagulants for removal of COD and color from textile wastewater. Glob Nes J. 2013;15(4):522-528. This was the singular reason for the assay in this study.
The relatively good and significant flocculating efficiency of the purified bioflocculant against kaolin clay suspension and waste water treatment in this study may be ascribed to its inherent constituents and thermally stable nature. Consequent upon this, it may be inferred that the S. platensis bioflocculant may serve as a good replacement for conventional flocculants in wastewater treatment and fermentation industries.
The overall significant bioflocculating efficiencies exhibited in this study have demonstrated that the bioflocculant from S. platensis possesses high flocculating activity in kaolin suspension. However, to be applied in treatment of water, the performance of the bioflocculant should also be assessed in river water and wastewater. The river water is a representative of surface water with less COD and turbidity1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674. and the results of our evaluations on the river water in this study was quite interesting and of significant importance judging by previous reports. Gong1717 Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol. 2008;99:4668-4674. reported that the flocculant of S. ficaria flocculated river water at an efficiency of 90.4% with COD and turbidity removal efficiencies of 87.1% and 84.2%, respectively. In another study, maximum COD and turbidity removal of 61.2% and 95.6% were achieved when a bioflocculant produced by B. licheniformis X14 was used to treat low-temperature drinking water.3737 Li Z, Zhong S, Lei H, Chen RW, Yu Q, Li HL. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour Technol. 2009;100:3650-3656. In addition, the industrial application of bioflocculant produced by S. platensis was also evaluated on meat processing wastewater and its turbidity removal efficiency may be attributed to polymer-floc interaction of the bioflocculant, which leads to increase in agglomeration of particles. On the other hand, bioflocculant produced by Azotobacter indicus was able to reduce biological oxygen demand (BOD), COD and suspended solids in wastewater samples in the range of 38–80%, 37–79% and 41–68% respectively at a bioflocculant dosage of 500 mg L-1.4343 Patil SV, Patil CD, Salunke BK, Salunkhe RB, Bathe GA, Patil DM. Studies on characterization of bioflocculant exopolysaccharide of Azotobacter indicus and its potential for wastewater treatment. Appl Biochem Biotechnol. 2011;163:463-472. Our findings confirmed the possibility of the use of the bioflocculant in flocculation processes.
The FTIR spectrum of the purified bioflocculant analyzed revealed the presence of different functional groups. While the band with the peak at 3417.4 cm-1 can be assigned to the stretching vibration of hydroxyl and amino groups,4444 Desouky AM, El-haleem A, Al-thani R, Al-mokemy T, Al-marii S, Hassan F. Isolation and characterization of extracellular bioflocculants produced by bacteria isolated from Qatari ecosystems. Pol J Microbiol. 2008;3(57):231-239. those at 2923.7 cm-1, 2734.6 cm-1 and 2395.7 cm-1 indicated aliphatic C === Inserir caracter correspondente ao PDF === H bands. The peak at 1755.1 cm-1 is characteristics of C === Inserir caracter correspondente ao PDF === O stretching vibration in carboxylic group and the intense peak at 1386.7 cm-1 is a representative of CN stretching group.4545 Fujita M, Ike M, Tachibana S, Kitada G, Kim SM, Inoue Z. Characterization of bioflocculant produced by Citrobacter sp. TKF04 from acetic and propionic acids. J Biosci Bioeng. 2000;98:40-46. The peaks at 827.4 cm-1 and 628.8 cm-1 are known to be characteristics of sugar derivatives and β-glycodisic bonds between sugar monomers.2525 Xiong Y, Wang Y, Yu Y, Li Q, Wang H, Chen R. Production and characterization of a novel bioflocculant from Bacillus licheniformis. Appl Environ Microbiol. 2010;76:2778-2782. The presence of hydroxyl, amino, carboxyl group present in the IR spectrum of S. platensis might be responsible for the significant and excellently elicited flocculating activity in this study.
During the process of flocculation, kaolin clay adsorbed on to the binding sites of bioflocculant which resulted into larger flocs formation and sedimentation of the kaolin particles. This was evidently shown in this study with the interaction of the purified bioflocculant and the kaolin clay suspension forming compacted flocs (Fig. 4B) as against both the purified bioflocculant alone (Fig. 4A) and the kaolin clay suspension (Fig. 4C).
The thermogravimetric analysis was conducted to monitor the pyrolysis profile of the test bioflocculant. The over 60% weight retained following heat treatment of the purified bioflocculant at the highest treatment temperature (900 ºC) was an affirmation of the thermally stable nature of the bioflocculant. The elemental constituents revealed by the EDX analysis are a further attestation of the confirmed chemical constituents of the purified bioflocculant to be rich in polysaccharides and proteins.
Conclusion
The bioflocculant produced by S. platensis SFD 20 is composed of mostly polysaccharide with the addition of proteins. This feature contributes to its thermostability. The purified bioflocculant had a high flocculating activity at a minimal dosage requirement, against kaolin clay, river water and meat processing wastewater. The bioflocculant could also remove COD and turbidity in meat processing wastewater and river water. Based on the outstanding performance of the test bioflocculant, it could be a useful tool in environmental bioremediation and other biotechnological processes. Moreover, further studies will focus on evaluating the ability of the purified bioflocculant to flocculate other wastewaters and remove heavy metals.
Acknowledgments
Financial support from the National Research Foundation of South Africa (Grant No. 94923) is highly appreciated.
REFERENCES
-
1Guo J, Jing Y, Xin X, et al. Characterization and flocculation mechanism of a bioflocculant from hydrolyzate of rice stover. Bioresour Technol 2015;177:393-397.
-
2Salehizadeh H, Yan N. Recent advances in extracellular biopolymer flocculants. Biotechnol Adv 2014;32:1506-1522.
-
3Sam S, Kucukasik F, Yenigun O, Nicolaus B, Oner ET, Yukselen MA. Flocculating performances of exopolysaccharides produced by a halophilic bacterial strain cultivated on agro-industrial waste. Bioresour Technol 2011;102:1788-1794.
-
4Li Q, Liu HL, Qi Q, Wang F, Zhang Y. Isolation and characterization of temperature and alkaline stable bioflocculant from Agrobacterium Sp. M503. New Biotechnol 2010;27:789-794.
-
5Liu W, Wang K, Li B, Yuan H, Yang J. Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour Technol 2010;101:1044-1048.
-
6Zhao G, Ma F, Wei L, Chua H. Using rice straw fermentation liquor to produce bioflocculants during an anaerobic dry fermentation process. Bioresour Technol 2012;113:83-88.
-
7Ahmad HRA, Azni I, Hamid HR, Al-joubory Y, Uemura BSU, Ibn A. Flocculation behavior and mechanism of bioflocculant produced by Aspergillus flavus J Environ Manage 2015;150:466-471.
-
8Batta N, Subudhi S, Lal B, Devi A. Isolation of a lead tolerant novel bacterial species, Achromobacter sp. TL-3: assessment of bioflocculant activity. Ind J Exp Biol 2013;51:1004-1011.
-
9Anthony U, Sekelwa C, Leonard M, Olubukola O, Farhad A, Anthony O. Thermostable bacterial bioflocculant produced by Cobetia Spp. isolated from Algoa Bay (South Africa). Int J Environ Res Pub Health 2012;9(6):2108-2120.
-
10Kämpfer P. The family Streptomycetaceae, Part I: Taxonomy. In: Dworkin M, Falkow S, Rosenberg E, Schleifer K, Stackebrandt E, eds. The Prokaryotes 2006:538.
-
11Reymond S, Janine C. Migrastatin and analogues: new anti-metastatic agents. Compt Rend Chim. 2010;11:1447-1462.
-
12Jensen PR, Dwight R, Fenical W. Distribution of actinomycetes in near shoretropical marine sediments. Appl Environ Microbiol 1991;57:1102-1108.
-
13Xia S, Zhang Z, Wang X, et al. Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresour Technol 2008;99:6520-6527.
-
14Lane DJ. 16/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M, eds. Nucleic Acid Techniques in Bacterial Systematics Chichester/New York/Brisbane/Toronto/Singapore: John Wiley and Sons; 1991:115–175.
-
15Chen H, Zhang JF, Jiang PJ, Yang SL, Liu ZL. Composition and characterization of microbiological flocculant SC06. Environ Chem 2002;21:360-364.
-
16Piyo N, Cosa S, Mabinya LV, Okoh AI. Assessment of bioflocculant production by Bacillus sp. Gilbert, a marine bacterium isolated from the bottom sediment of Algoa Bay. Mar Drugs 2011;9:1232-1242.
-
17Gong W, Wang S, Sun F, Liu XW, Yue QY, Gao BY. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour Technol 2008;99:4668-4674.
-
18Lee SH, Shin WS, Shin MC, Choi SJ, Park LS. Improvement of water treatment performance by using polyamine flocculants. Environ Technol 2001;22:653-659.
-
19Wang Y, Gao BY, Yue QY, Wei JC, Zhou WZ, Gu R. Color removal from textile industry wastewater using composite flocculants. Environ Technol 2010;28(6):629-637.
-
20Ugbenyen MA, Okoh AI. Characteristics of a bioflocculant produced by a consortium of Cobetia and Bacillus species and its application in the treatment of wastewaters. Water SA 2014;40(1):.
-
21Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin Phenol reagent. J Biol Chem 1951;193:265-275.
-
22Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956;28:350-356.
-
23Wang L, Ma F, Lee DJ, Wang AJ, Ren NQ. Bioflocculants from hydrolysates of corn stover using isolated strain Ochrobactium cicero W2. Bioresour Technol 2013;145:259-263.
-
24Cosa S, Ugbenyen MA, Mabinya LV, Okoh IA. Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr J Microbiol Res 2013;7(23):2925-2938.
-
25Xiong Y, Wang Y, Yu Y, Li Q, Wang H, Chen R. Production and characterization of a novel bioflocculant from Bacillus licheniformis Appl Environ Microbiol 2010;76:2778-2782.
-
26Ugbenyen MA, Okoh AI. Flocculating properties of a bioflocculant produced by Bacillus sp. isolated from a marine environment in South Africa. Chem Biochem Eng 2013;27(4):511-518.
-
27Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus Bioresour Technol 2013;127:489-493.
-
28Sanjukta S, Neha B, Mihirjyoti P, et al. Bioflocculant production and biosorption of zinc and lead by a novel bacterial species, Achromobacter sp. TERI-IASST N, isolated from oil refinery waste. Chemosphere 2014;113:116-124.
-
29Giri SS, Harshiny M, Sen SS, Sukumaran V, Park SC. Production and characterization of a thermostable bioflocculant from Bacillus subtilis F9, isolated from wastewater sludge. Ecotoxicol Environ Saf 2015;121:45-50.
-
30Salehizadeh H, Shojaosadati SA. Removal of metal ions from aqueous solution by polysaccharide produced from Bacillus firmus. Water Res 2003;37:4231-4235.
-
31Shu CH, Lung MY. Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia camphorate in batch cultures. Process Biochem 2004;39:931-937.
-
32Zheng Y, Ye ZL, Fang XL, Li YH, Cai WM. Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour Technol 2008;99:7686-7691.
-
33Zhang ZQ, Lin B, Xia SQ, Wang XJ, Yang AM. Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. Environ Sci 2007;19:667-673.
-
34Yim JH, Kim SJ, Ahn SH, Lee HK. Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresour Technol 2007;98:361-367.
-
35Okaiyeto K, Nwodo UU, Mabinya LV, Okoli AS, Okoh AI. Evaluation of flocculating performance of a thermostable bioflocculant produced by marine Bacillus sp. Environ Technol 2016, http://dx.doi.org/10.1080/09593330.2015.1133717
» https://doi.org/10.1080/09593330.2015.1133717 -
36Nwodo UU, Agunbiade MO, Green E, Mabinya LV, Okoh AI. A freshwater Streptomyces, isolated from Tyume river, produces a predominantly extracellular glycoprotein bioflocculant. Int J Mol Sci 2012;13:8679-8695.
-
37Li Z, Zhong S, Lei H, Chen RW, Yu Q, Li HL. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour Technol 2009;100:3650-3656.
-
38Nwodo UU, Agunbiade MO, Green E, Nwamadi M, Rumbold K, Okoh AI. Characterization of an exopolymeric flocculant produced by a Brachybacterium sp. Materials 2013;6:1237-1254.
-
39Salehizadeh H, Shojaosadati SA. Extracellular biopolymeric flocculants-recent trends and biotechnological importance. Biotechnol Adv 2001;19:371-385.
-
40Yukselen MA, Gregory J. The effect of rapid mixing on the break-up and re-formation of flocs. Chem Technol Biotechnol 2004;79(7):782-788.
-
41Wang L, Ma F, Qu Y, Sun D, Li A, Guo J. Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. W J Microbiol Biotechnol 2011;27:2559-2565.
-
42Patel H, Vashi RT. Comparison of naturally prepared coagulants for removal of COD and color from textile wastewater. Glob Nes J 2013;15(4):522-528.
-
43Patil SV, Patil CD, Salunke BK, Salunkhe RB, Bathe GA, Patil DM. Studies on characterization of bioflocculant exopolysaccharide of Azotobacter indicus and its potential for wastewater treatment. Appl Biochem Biotechnol 2011;163:463-472.
-
44Desouky AM, El-haleem A, Al-thani R, Al-mokemy T, Al-marii S, Hassan F. Isolation and characterization of extracellular bioflocculants produced by bacteria isolated from Qatari ecosystems. Pol J Microbiol 2008;3(57):231-239.
-
45Fujita M, Ike M, Tachibana S, Kitada G, Kim SM, Inoue Z. Characterization of bioflocculant produced by Citrobacter sp. TKF04 from acetic and propionic acids. J Biosci Bioeng 2000;98:40-46.
Edited by
Publication Dates
-
Publication in this collection
Oct-Dec 2018
History
-
Received
4 Oct 2016 -
Accepted
11 Feb 2017 -
Published
5 Apr 2018